MOLECULAR CLONING AND EXPRESSION OF A FRAGMENT OF THE GENE CODIFYING FOR THE PROTEIN ERNS OF CLASSICAL SWINE FEVER VIRUS
CLONAJE MOLECULAR Y EXPRESIÓN DE UN FRAGMENTO DEL GEN QUE CODIFICA PARA LA PROTEÍNA DEL VIRUS DE LA PESTE PORCINA CLÁSICA, ERNS
J.A. Agüero*†, O. Sánchez**, Maritza Barrera* y J.R. Toledo**
*National Centre for Animal and Planth Health (CENSA). Carretera de Jamaica y Autopista Nacional. Apto.10. San José de las Lajas. La Habana. CP. 32700. Cuba.Corresponding author: jaaguero@censa.edu.cu **Centre for Genetic Engineering and Biotechnology (CIGB). Ave 31 e/ 158 y 190 Playa. P.O. Box 6162. Habana 10600. ]]>
ABSTRACT
Classical swine fever virus (CSFV), belonging to the genus Pestivirus of the Flaviviridae family, is an enveloped positive stranded RNA virus highly contagious that can cause a fatal disease, characterized by fever, leukopenia and hemorrhage, with substantial economic losses. There is a great demand for a marker vaccine against CSFV. C, Erns, E1, and E2 are the structural proteins of the virus. E2 is the best candidate to be incorporated in vaccine, while Erns becomes an ideal candidate as an antigen in a differential diagnostic test. A synthetic fragment of the Erns gene (codifying for aa 109-160) was subcloned into pET28a vector. The cloning was screened by restriction analysis. The gene was expressed as a his-tag fusion protein in BL21 (DE3) E. coli strain. The recombinant polypeptide formed aggregates of about 7.9; 15.8; 23.7; 31.6; 39.5; 47.5 and 55.3kDa. The protein was not recognized by Western blot using an antibody against the virus. Using an ion metal affinity chromatography procedure, a 90% pure recombinant product was obtained. The potential use of this antigen for detection of CSFV antibodies should be further evaluated.
Key words: Erns; CSFV; recombinant protein; IMAC
RESUMEN
El virus de la fiebre porcina clásica (CSFV), perteneciente al género Pestivirus de la familia Flaviviridae, es un virus ARN positivo, altamente contagioso capaz de causar una enfermedad fatal caracterizada por fiebre, leucopenia y hemorragia, con considerables pérdidas económicas. Existe una gran demanda de una vacuna marcadora contra CSFV. C, Erns, E1 and E2 son las proteínas estructurales del virus. E2 es el mejor candidato para ser incorporado a una vacuna, mientras Erns resulta entonces un candidato ideal como antígeno en un ensayo de diagnóstico diferencial. En el vector pET28a se subclonó un fragmento sintético del gene que codifica para Erns (codificante para los aa 109-160). El clonaje se monitoreó por análisis de restricción. El gen se expresó como una proteína de fusión a histidina en la cepa BL21(DE3) de E. coli. El polipéptido recombinante formó agregados de aproximadamente 7.9; 15.8; 23.7; 31.6; 39.5; 47.5 y 55.3kDa. La proteína no fue reconocida por anticuerpos contra el virus por Western blot. Con el empleo de la cromatografía de afinidad a iones metálicos se obtuvo un producto recombinante con un 90% de pureza. Más adelante debe ser evaluado el empleo potencial de este antígeno para la detección de anticuerpos contra CSFV.
Palabras clave: Erns; CSFV; proteína recombinante; IMAC
]]>
INTRODUCTIONClassical swine fever virus (CSFV), Bovine viral diarrhoea virus (BVDV) and Border disease virus (BDV) belong to the genus Pestivirus of the Flaviviridae family (4). CSFV is highly contagious and can cause a fatal disease in pigs. Although pigs can be infected by all these pestiviruses (16; 22), only CSFV induces severe disease and it is often fatal. The disease is characterized by fever, leukopenia and hemorrhage, and can run an acute, chronic, or subclinical course substantial economic losses (18). In several countries, classical swine fever (CSF) outbreaks are controlled by the eradication of all pigs from infected and neighbouring farms. Although effective live-attenuated vaccines are available, sometimes they are not used due to the fact that vaccinated and infected pigs are serologically indistinguishable. That is why there is a great demand for a marker vaccine against CSFV.
Like other members of the family, CSFV is an enveloped positive stranded RNA virus (13, 3). Similar to other members of the genus, the 12.5-kb CSFV genome contains a single large open reading frame encoding a polyprotein precursor of approximately 4,000 amino acids (aa). The precursor is cleaved co- and post-translationally by cellular and viral proteases into structural proteins C, Erns, E1, and E2 and nonstructural proteins NS2, NS3, NS4A, NS4B, NS5A, and NS5B (19).
Antibodies against Erns, E2, and NS3 have been demonstrated in infected animals (7; 10; 14; 15; 16), with Erns and E2 capable of inducing neutralizing antibodies (5; 6; 24). Glycoprotein E2 is the best candidate to be incorporated in vaccine because it is the most immunogenic protein of pestiviruses and elicits high titers of neutralizing antibodies after infection (20; 25). Additionally, vaccination of pigs with E2 has been shown to give protection against a lethal homologous challenge (1; 5). In this scenario, Erns becomes an ideal candidate as an antigen in a differential diagnostic test. It has been shown that antibodies to full-length Erns or even a 37-mer peptide derived from its C-terminal end (aa 191 to 227) could be used as an indicator of CSFV infection in pigs (7; 14). Recent studies have indicated that an Erns-based enzymelinked immunosorbent assay (ELISA) can be used as a companion diagnostic test to identify CSFV-infected pigs in herds vaccinated with the E2 subunit marker vaccine (14). Recently, an immunodominant region encompassing three overlapping antigenic regions that induce antibody responses during CSFV infection: aa 65 to 145, aa 84 to 160 , and aa 109 to 220 was mapped (11) and antibody responses of pigs to defined Erns fragments (selected from the above mentioned regions), after classical swine fever virus (CSFV) infection, were studied by using an enzyme-linked immunosorbent assay (ELISA). In this study, Erns aa 109-145 and Erns aa 109-160 offered the best specificity and sensitivity (Lin et al., 2005).
In this work, the molecular cloning of the DNA segment encoding for the Erns aa 109-160 fragment is described, as well as its expression and the purification of the product obtained.
MATERIALS AND METHODS
Vectors, bacterial strains and growth conditions. The XL1 Blue competent cells, as well as BL21(DE3) competent were obtained from the strain bank of the Centre for Genetic Engineering and Biotechnology (CIGB) (La Habana, Cuba). The plasmid 050778pPCR-Script containing the synthetic fragment from the Erns gene fragment, codifying for the Erns aa 109-160 polypeptide (Brescia strain), was provided by GeneArt. The fragment was designed in a way that its sequence was optimized for expression in E.coli, additionally Nde I and BamH I restriction sites were introduced in its extremes 5´and 3´, for its subsequent, in frame, cloning in the expression vector pET28a. Competent XL1-Blue cells were separately transformed with both plasmids and growth O/N at 37°C in Luria-Bertani (LB) medium supplemented with 100µg of ampicillin/mL (050778pPCR-Script) and 50µg of kanamycin/mL (pET28a).
Vector Preparation. The pET28a plasmid (Novagen) was purified using the Wizard Plus SV Minipreps DNA Purification System (Promega). 5µg of the vector were digested with 50u each of Nde I and BamH I (Promega) in 100µL of total volume and in presence of 0.1mg/mL BSA and 10µL of MULTI-CORETM buffer, the digestion was carried out for 3 hours at 37°C. The product digested was applied in a low melting point agarose gel at 0.8% from where, after electrophoresis (constant voltage 100V; in TB buffer), the 5329bp band corresponding to digested pET28 plasmid, was isolated. This band was purified using the QIAquick Gel Extraction Kit (QIAGEN).
When kits where used, all the procedures were carried out according to the manufacturer's instructions. ]]>
Recombinant plasmid generation. The 050778pPCR-Script plasmid was digested with Nde I and BamH I restriction enzymes to obtain the Erns gene fragment. For this, 5µg of the plasmid were digested with 50u each of Nde I and BamH I (Promega) in 100µL of total volumen and in presence of 0.1mg/mL BSA and 10µL of MULTI-CORETM buffer; the digestion was carried out for 3 hours at 37°C. The product digested was applied in a low melting point agarose gel at 1.5% from where, after electrophoresis, the 173bp band corresponding to the Erns gene fragment, was isolated. This digestion product was purified using the QIAquick Gel Extraction Kit (QIAGEN).To obtain the construction pET28a-Erns, the previously digested Erns gene fragment and pET28a vector were ligated. In the ligation mix the vector and insert concentrations were 31ng/mL and 5ng/mL, respectively. 4u of T4 ligase (Promega), 2mL of T4 ligasa 10x buffer (Promega) were employed in a 20mL of total volumen and the reaction was carried out at 22°C, for 5h.
For transformation, Ca2+ competent cells from XL1-Blue of E. coli were employed, and transformed as follow: 10µL of the ligation product were added to 150µL of competent cells. The mix was incubated in ice for 20 min, then 2 min at 42°C and to finish 5 min again in ice. 1mL of LB medium was added to the mix and this was incubated 1h at 37°C at 50rpm in an orbital shaker. This culture was inoculated to LB plates supplemented with 50µg of kanamycin/mL and the plates were incubated O/N at 37°C.
The screening of recombinant clones was done by restriction analysis with Nde I and BamH I restriction enzymes.
Expression of recombinant Erns fragment. For the expression experiments, the method of autoinduction developed by Studier (2005) (21) was employed with some modifications. E. coli BL21(DE3) cells harboring the expression construct pET28a-Erns-1 were cultured O/N at 37°C in a Luria-Bertani broth supplemented with 50µg of kanamycin/mL. Next day, 100mL (in a 1L erlenmeyer) of auto-induction media (1% triptone; 0.5% yeast extract; 25mM Na2HPO4; 25mM KH2PO4; 50mM NH4Cl; 5mM Na2SO4; 0.5% glicerol; 0.05% glucose; 0.2% alfa-lactose monohydrate; 2mM MgSO4 heptahydrate; 0.05mM FeCl3) supplemented with 200µg of kanamycin/mL were inoculated with 10µL of the O/N inoculum and grown for 16 hours at 37°C with shaking at 310rpm,. The cells were harvested by centrifugation at 5,000 x g for 10 min and conserved at -20°C.
Purification of recombinant proteins by metal ion affinity chromatography (IMAC). Cell pellets from cultures were resuspended in 10mL of protein extraction buffer (6M guanidine hydrochloride, 0.1M Na-Phosfate buffer, [pH 8.0]) and lysed with a French press at 1,500 kgf/cm2. The homogenates were spun at 12,000 x g for 30 min at 4°C, and the supernatant was filtered by 0.45µm and then loaded onto a Protein Purification Resin (Promega), previously equilibrated with the extraction buffer column (1 by 1 cm) of HisLink™. The column was washed with buffer A (8M urea, 0.1M Na-Phosfate buffer [pH 8.0]), followed by buffer B (buffer A plus 0.5 M NaCl [pH 6.3]) and buffer C (buffer B plus 5 mM imidazole [pH 5.9]). The denatured protein was then refolded on the column by washes with Tris-buffered saline (Na-Phosfate buffer) (pH 7.4) containing 1 M urea, followed by Na-Phosfate buffer (pH 7.4). The recombinant protein was eluted and collected with fractions of 10 ml from the column using Na-Phosfate buffer (pH 7.4) containing 500 mM imidazole. A final washing step was done using Na-Phosfate buffer (pH 7.4) containing 500 mM imidazole and 8 M urea. The percent of Erns fragment in each fraction was determined by densitometry of the Coomassie Blue stained gels with the Molecular Analyst software. Proteins were quantified by using the Bradford method (2), with bovine serum albumin as a standard.
Sera. The sera from pigs experimentally infected with CSFV were provided by the Virology group of the National Centre for Animal and Plant Health (CENSA) (Cuba).
SDS-PAGE and Western blot. Briefly, SDS-PAGE was performed at constant current of 30mA in a 15% acrylamide gels. The proteins were analyzed either through staining with Coomassie Blue or by Western blotting. From the 10mL fractions, 10µL aliquots were taken and analized by SDS-PAGE and Western Blotting. For Western blots, the proteins were transferred to nitrocellulose membranes using semi-dry transfer equipment. The membranes were blocked for 1 hour with PBS-BSA 1%. The blots were probed with swine CSFV antisera from experimentally infected pigs for 2 hours at 37°C. Bound antibodies were detected by using horseradish peroxidase (HRP)_conjugated anti_swine IgG antibodies (Sigma) and a 4-chloro-1- naphthol-H2O2 substrate kit (Bio-Rad), according to the manufacturer's instructions.
RESULTS AND DISCUSSION ]]>
The focus of this study is the obtainment of a recombinant fragment of the CSFV Erns structural protein. This polypeptide (Erns aa 109-160) had been previously described as good antigen candidate for detection of antibodies against the virus, offering, together with Erns aa 109-145 fragment, the best specificity and sensitivitiy measured by ELISA, among other fragments from an immunodominant Erns region (12). In particular, antibody responses to Erns aa 109-160 can be detected as early as 7 dpi, then this small Erns fragment would offer a diagnostic antigen superior to the full-length protein, which can only be used to detect CSFV-specific antibodies in vaccinated or unvaccinated pigs as early as 14 dpi (14).The 173bp band corresponding to the Erns aa 109-160 gene fragment was sussesfully isolated from plasmid 050778pPCR-Script (results are not shown) and subcloned in the pET28a vector using the Nde I and BamH I restriction sites introduced in the synthetic gene. Four positive recombinant clones were detected by restriction analysis.
The initial transformation was done in XL1-Blue cells of E. coli because this strain facilitates the propagation and subsequent purification of plasmids, but for protein production, in order to quarantee greater control and a higher level of the gene expression it was decided to use BL21(DE3) cells of E. coli, which were transformed with the four recombinant plasmids. In pET plasmids, the gene expression is under the control of the T7 RNA polymerase. This enzyme is so specific, active, and processive that the amount of target RNA produced can be comparable to the amount of ribosomal RNA in a cell. A problem in using inducible T7 expression systems is that T7 RNA polymerase is so active that a small basal level can lead to substantial expression of target protein even in the absence of added inducer. If the target protein is sufficiently toxic to the host cell, establishment of the target plasmid in the expression host may be difficult or impossible, or the expression strain may be unstable or accumulate mutations. In BL21(DE3)-pET system, this problem is overcome, the lac operator sequence (the binding site for lac repressor) is placed in both, the start site of a T7 promoter (of the pET plasmid) and in the lac promoter, under the control of which is the T7 RNA polimerase in cell genomes. In this way, the basal level of target protein in uninduced cells is substantially reduced, but induction leads to the typical high levels of expression (21). The BL21(DE3) clones, obtained from the four original recombinant plasmids, were thus designated pET28a-Erns1, pET28a-Erns2, pET28a-Erns3 and pET28a-Erns4. This clones were then tested for protein expression, using the classical IPTG induction method. In all four cases, a band of approximate 8kDa, corresponding to the expected size, was detected in the induced cell lysates by SDS-PAGE (Fig. 1). Taking into consideration that the level of expression was similar in all clones, the work with pET28a-Erns1 clone was continued.
]]>
For the rest of the experiments, it was decided to test the auto-induction method described by Studier (2005) (21). Auto-induction allows efficient screening of many clones in parallel for expression and solubility, as cultures have only to be inoculated and grown to saturation. Yields of target protein are typically several-fold higher than those obtained by conventional IPTG induction. In addition, IPTG, a very expensive reagent, is substituted by the most shipper ?-lactose. In this system, lactose is used as inductor, and the unintended level of expression of the target protein in the initial moments of the culture, is abolished by the addition of glucose to the media. Glucose prevents induction by lactose by well-studied mechanisms. In the original protocol, expression strains grow O/N to saturation in non-inducing medium, which is then inoculated into the autoinduction medium (21). This non-inducing medium was substituted by the normal LB, because the yields were similar and the LB medium was easier to prepare.
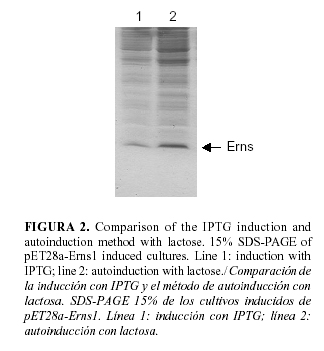
The obtained results were categorical. Although the synthesis level of the recombinant protein was similar when both methods of induction were used (about 10%). When auto-induction was employed, the culture growth was always higher in at least 3-4 times in comparison with the growth of the IPTG culture, given yields of target protein several-fold higher than the obtained by conventional IPTG induction (Fig. 2).
]]>
When it was tried to antigenically confirm the expression of our target polypeptide, Western blot assayed with anti-CSFV antibodies failed to detect the expression of the protein obtained, or the reaction was very weak (results are not shown), although the polypeptide was expressed in abundance, as evidenced by SDS-PAGE. These results are in agreement with the previously obtained. First, some data showed that Erns aa 109_145, the consensus region defined by AR1, AR2, and AR3 (three overlapping antigenic region), and Erns aa 109_160, a fragment 15 amino acids larger than the consensus region, were not recognized by CSFV antisera on Western blot (11). Other results showed that both Erns aa 109_145 and Erns aa 109_160, after refolding and eluting from a Ni-agarose column, were recognized by antibodies in sera from CSFV-infected animals in ELISA (12). These results provide evidence that the epitopes located within the consensus region are conformational and, as they are destroyed under the denaturing conditions of the SDS-PAGE, they cannot be detected by Western blot. This is consistent with the proposed three-dimensional model of Erns (8), which predicts the cysteine-rich consensus region to be on a large surface-exposed loop between helix 6 and helix 7 and likely to form two disulfide bonds in this region.
When the recombinant proteins are expressed at high levels, they usually form insoluble inclusion bodies (9; 17). For any further use, the proteins should be refolded in vitro. Surprisingly, when the solubility of our product was checked, it was observed that our protein was located in the supernatant after the centrifugation of the lysis suspension, without the use of any chaotropic agent, what meant, that it was soluble (Fig. 3). Maybe this was influenced by the small size of our polypeptide.
]]>
However, when the purification experiments started, it was impossible to attach the protein to the column, and in all cases it was eluted in the first wash buffer. Supposing that it was due to an effect of the protein conformation, that made the protein hide its Histidine tale in the inner part of its three dimensional structure, it was decided to add urea to the lysis buffer. After the addition of at least 1M urea our suspicion was confirmed, the protein could be attached to the quelating matrix. After this result, it was decided to use, for lysis and protein purification, a very similar protocol to the one developed by Lin et al. (2005) (12) that included 6M guanidine hydrochloride in the lysis buffer.
]]>
The supernantant thus obtained, was then applied into a His-Link matrix, charged with Ni ions. The Erns aa 109-160 polypeptide was obtained with more than 90% of purity after elution with 500mM of imidazole without urea, because the protein was refolded in the column (Fig. 4). The purity was so high, that it was not necessary to use a second anion exchange purification step, as it was employed in the original work (12).
]]>
Purification fractions were first analyzed in reducing conditions by SDS PAGE. In these conditions, in the Erns aa 109-160 elution fraction, an additional protein of 15.8kDa was observed. As this protein was not observed in the same fraction of the negative control (the same purification protocol for an induced culture of the pET28a plasmid without the insert, was runned in parallel), then, and taking into account that Erns protein forms a disulfide-bridged homodimer in the virion (8; 23), it was possible that the 15.8kDa protein was an aggregation product of the Erns aa 109-160 polypeptide. When samples were analyzed under non-reducing conditions by SDS-PAGE, a very complex aggregation pattern was observed, seven proteins with sizes coincided with the possible polymers that could form our truncated Erns (Fig. 5). This multimerization is not reported by Lin et al. (12). One possible explanation is the presence of four cysteines in the sequence of the Erns aa 109-160 polypeptide. Although in the Erns predicted structure, these cysteins do not play any role in the formation of disulfide bridges between two Erns homodimers, and only in the formation of intra disulfide bridges (8). The overexpression of this truncated Erns version in E. coli could induce this rare behaviour.
]]>
Whether this phenomenon affects or not the antigenicity of the recombinant Erns aa 109-160 fragment in an ELISA based detection system is an open question. Further experiments have to be done to elucidate the impact of this aggregation in the antigenicity of this polypeptide.
CONCLUSIONS
As conclusions, it can be said that the construct obtained, as well as the described methodology, including the use of the auto induction protein expression system, will permit the obtainment, in a very simple way, of large quantity of Erns antigen that could facilitate the development and validation of a test to be used for serological survey.
Of course, the potential use of this polypeptide, as an antigen for detection of CSFV antibodies will depend on the antigenicity studies that still have to be carried out.
]]>
ACKNOWLEDGEMENTSThe authors are grateful to Alejandro Martin and the members of the lab of Dengue Vaccine of the CIGB, for their technical and material assistance.
REFERENCES
1. Bouma A, de Smit AJ, de Kluijver EP, Terpstra C, Moormann RJ. Efficacy and stability of a subunit vaccine based on glycoprotein E2 of classical swine fever virus. Vet Microbiol. 1999;66:101-114.
2. Bradford M. A rapid and sensitive method for the quantitation of microgram quantities of proteins utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248-154.
3. Colett MS, Larson R, Gold C, Strick D, Anderson DK, Purchio AF. Molecular cloning and nucleotide sequence of the pestivirus bovine viral diarrhea virus. Virology. 1988;165:191-199.
4. Francki RIB, Faquet DL, Knudson DL, Brown F. Fifth report of the International Committee on the Taxonomy of Viruses. Arch Virol Suppl. 1991;2:223-233.
5. Hulst MM, Westra DF, Wensvoort G, Moormann RJ. Glycoprotein E1 of hog cholera virus expressed in insect cells protects swine from hog cholera. J Virol. 1993;67:5435-5442.
6. Konig M, Lengsfeld T, Pauly T, Stark R, Thiel HJ. Classical swine fever virus: independent induction of protective immunity by two structural glycoproteins. J Virol. 1995;69:6479-6486.
7.Langedijk JP, Middel WG, Meloen RH, Kramps JA, de Smit JA. Enzyme-linked immunosorbent assay using a virus type-specific peptide based on a subdomain of envelope protein Erns for serologic diagnosis of pestivirus infections in swine. J Clin. Microbiol. 2001;39:906-912.
8. Langedijk JP, van Veelen PA, Schaaper WM, de Ru AH, Meloen RH, Hulst MM. A structural model of pestivirus Erns based on disulfide bond connectivity and homology modeling reveals an extremely rare vicinal disulfide. J Virol. 2002;76:10383-10392.
9. Li M, Su ZG, Janson JC. In vitro protein refolding by chromatographic procedures. Protein Expr Purif. 2004;33(1):1-10.
10.Lin M, Lin F, Mallory M, Clavijo A. Deletions of structural glycoprotein E2 of classical swine fever virus strain alfort/187 resolve a linear epitope of monoclonal antibody WH303 and the minimal N-terminal domain essential for binding immunoglobulin G antibodies of a pig hyperimmune serum. J Virol. 2000;74(24):11619-11625.
11.Lin M, Trottier E, Pasick J, Sabara Marta. Identification of Antigenic Regions of the Erns Protein for Pig Antibodies Elicited during Classical Swine Fever Virus Infection. J Biochem. 2004;136(6):795-804.
12.Lin M, Trottier E, Pasick J. Antibody Responses of Pigs to Defined Erns Fragments after Infection with Classical Swine Fever Virus. Clinical Diagnostic Lab Immunol. 2005;12(1):180-186.
13.Moennig V. Characteristics of the virus, p. 55-58. In B. Liess (ed.). Classical swine fever and related viral infections. Martinus Nijhoff Publishing, Boston, Mass. 1988.
14.Moormann RJ, Bouma A, Kramps JA, Terpstra C, de Smit HJ. Development of a classical swine fever subunit marker vaccine and companion diagnostic test. Vet Microbiol. 2000;73(2-3):209-219.
15.Moser C, Ruggli N, Tratschin JD, Hofmann MA. Detection of antibodies against classical swine fever virus in swine sera by indirect ELISA using recombinant envelope glycoprotein E2. Vet Microbiol. 1996;51(1-2):41-53.
16.Paton DJ, Ibata G, Edwards S, Wensvoort G. An ELISA detecting antibody to conserved pestivirus epitopes. J Virol Methods. 1991; 31:315-324.
17.Platis D, Foster GR. High yield expression, refolding, and characterization of recombinant interferon alpha2/alpha8 hybrids in Escherichia coli. Protein Expr Purif. 2003;31(2):222-30.
18.Pluimers FH, de Leeuw PW, Smak JA, Elbers AR, Stegeman JA. Classical swine fever in The Netherlands 1997-1998: a description of organisation and measures to eradicate the disease. Prev Vet Med. 1999;42:139-155.
19.Rice MC, Lindenbach BD. Flaviviridae: the viruses and their replication, 2001: p. 991-1041. In D. M. Knipe and P. M. Howley (ed.), Fields virology. Lippincott Williams & Wilkins, Philadelphia, Pa.
20.Rumenapf T, Stark R, Meyers G, Thiel HJ. Structural proteins of hog cholera virus expressed by vaccinia virus: further characterization and induction of protective immunity. J Virol. 1991;65:589-597.
21.Studier FW. Protein production by auto-induction in high-density shaking cultures. Protein Expr and Purif. 2005;41:207-234.
22.Terpstra C, Wensvoort G. Natural infections of pigs with bovine viral diarrhoea virus associated with signs resembling swine fever. Res Vet Sci. 1988;45:137-142.
23.Thiel HJ, Stark R, Weiland E, Rumenapf T, Meyers G. Hog cholera virus: molecular composition of virions from a pestivirus. J Virol. 1991;65:4705-4712.
24.van Rijn PA, Bossers A, Wensvoort G, Moormann RJ. Classical swine fever virus (CSFV) envelope glycoprotein E2 containing one structural antigenic unit protects pigs from lethal CSFV challenge. J Gen Virol. 1996;77:2737-2745.
25.van Zijl M, Wensvoort G, de Kluyver E, Hulst M, van der Gulden H, Gielkens A, et al. Live attenuated pseudorabies virus expressing envelope glycoprotein E1 of hog cholera virus protects swine against both pseudorabies and hog cholera. J Virol Methods. 1991;60:103-108.
]]>
(Recibido 7-11-2007; Aceptado 15-2-2008)
]]>
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}