Aspectos de interés sobre las acuaporinas en las plantas
Interest aspects about aquaporines in plants
Ms.C. Licet Chávez Suárez,I Alexander Álvarez Fonseca,II Dr.C. Ramiro Ramírez FernándezIII
IInvestigador Agregado de la Dirección de Investigaciones y Servicios Ambientales, Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, carretera vía Manzanillo km 17, Bayamo, Granma, Cuba.
IIReserva Científica de la Dirección de Investigaciones y Servicios Ambientales, Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, carretera vía Manzanillo km 17, Bayamo, Granma, Cuba. ]]>
IIIInvestigador Titular de la Dirección de Investigaciones y Servicios Ambientales, Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, carretera vía Manzanillo km 17, Bayamo, Granma, Cuba.
RESUMEN
Las acuaporinas son proteínas que regulan el movimiento de agua a través de las membranas celulares. Se ha comprobado la participación de estos transportadores en la respuesta de las plantas ante estrés abiótico. Es propósito de este trabajo reseñar aspectos fundamentales sobre las acuaporinas tales como: estructura y función, regulación y su participación en la respuesta de la planta ante el estrés abiótico.
Palabras clave: acuaporinas, estructura, función, regulación, estrés abiótico.
ABSTRACT
Aquaporine are proteins that regulated the water’s movement across the cellular membranes. It is comprobated the participation of this transporters participation on plant response against the abiotic stress. It is purpose of this article to resume essential aspects about the aquaporines such as: structure and function, regulation and their participation on plant response against abiotic stress.
Key words: aquaporine, structure, function, regulation, abiotic stress.
]]>
INTRODUCCIÓN
Las acuaporinas (AQPs) son proteínas formadoras de canales que regulan la entrada y salida de agua en la célula. Estas proteínas se encuentran presentes en todos los organismos y tienen pesos moleculares de entre 26 y 35 kDa (1).
Los estudios sobre las acuaporinas han conducido a la comunidad científica a descubrir otras funciones de las mismas, más allá del transporte de agua a través de las membranas. El transporte de solutos de gran significado fisiológico, tales como el CO2, H2O2, boro, o ácido silícico, permite relacionar las acuaporinas con muchas funciones, que incluyen el metabolismo del carbono, la respuesta ante el estrés oxidativo, y la nutrición mineral de la planta (2).
Cuando las plantas detectan cambios en la disponibilidad de agua en el medio que las rodea, sufren una alteración en el potencial osmótico; para balancear este cambio, las células vegetales regulan la expresión de sus AQPs. Se ha demostrado que estímulos ambientales como sequía y salinidad, así como incrementos en la síntesis del ácido abscísico, traen como consecuencia cambios en la regulación de los niveles de expresión de AQP (3).
Es objetivo del presente trabajo reseñar brevemente aspectos fundamentales sobre las acuaporinas tales como su estructura, función y regulación, así como su actividad en la planta en condiciones de estrés abiótico.
]]>
Descubrimiento de las acuaporinas
El transporte de agua a través de las membranas biológicas, proceso fundamental de los organismos vivos, fue motivo de estudio y debate por largas décadas. Inicialmente, se pensó que el agua atravesaba la membrana celular exclusivamente difundiendo entre sus moléculas lipídicas. Una vía ciertamente restringida que no concordaba con observaciones biofísicas en células con alta permeabilidad al agua, tal como el eritrocito de mamífero. Así, tempranos estudios biofísicos condujeron a predecir la existencia de canales de membrana que facilitaran el paso del agua (3).
Aunque las bases biofísicas del transporte de agua a través de las membranas biológicas habían sido anunciadas tan temprano como en la década de los 50, es el descubrimiento de las acuaporinas en 1992, el que popularizó el concepto de canales de agua de membrana (4).
En el año 2003, Peter Agre, investigador de la Universidad Johns Hopkins (Baltimore, USA), recibió el Premio Nobel de Química por haber identificado una proteína que permitía el paso selectivo de agua a través de la membrana celular, a la cual denominó “acuaporina”. Los trabajos habían comenzado hacía más de una década, cuando en los hematíes se descubrió una proteína inicialmente llamada CHIP-28 (channel-like integral protein-proteína integral semejante a canal de 28 kD ), que explicaba la elevada permeabilidad al agua en las membranas que la poseían. Posteriormente a su clonación y secuenciación, se sugirió el nombre de acuaporina-1 (AQP1) para esta proteína, el cual fue adoptado oficialmente por la Organización del Genoma Humano en 1997 (5).
Luego se describieron varias proteínas del mismo tipo, las cuales han recibido el mismo nombre seguido de un número secuencial en relación con la cronología de su descubrimiento. A partir de ahí la investigación alrededor de estas proteínas se ha incrementado notablemente y en la actualidad se han determinado gran parte de sus características moleculares y funcionales y se han descubierto varios miembros adicionales de esta familia (2).
]]>
Estructura de las acuaporinas
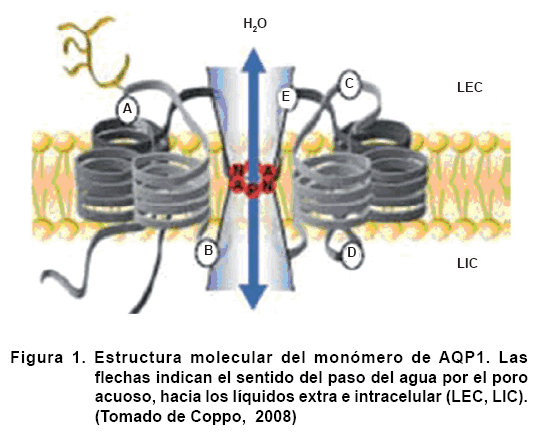
Las AQPs son proteínas transmembranosas que delimitan un canal de 2 nm de largo por 0,3 nm de ancho (Figura 1), su diámetro es de aproximadamente 0,28 nm.
Se calcula que las AQP son capaces de transportar 3 mil millones de moléculas de agua por segundo. Así, una porción de membrana de 10 cm2 filtraría un litro de agua en unos siete segundos (3).
Todas las AQPs presentan una similitud estructural bastante notable (Figura 1). Tienen sus segmentos amino y carboxilo terminal intracelulares. Están conformadas por dos dominios muy semejantes entre sí, unidas por el lazo C (6). Exhiben seis hélices transmembrana (7) y los lazos B y E son esenciales para la permeabilidad al agua del canal, es decir, son vitales en la formación del poro (8). Todas son tetraméricas (9), aunque algunas pueden formar oligómeros más pequeños, como AQP4.
El canal es hidrofóbico, por lo cual las moléculas de agua formarían una fila estableciendo puentes de hidrógeno con aminoácidos polares específicos dispuestos a lo largo del camino de permeabilidad. En el centro del canal, el diámetro es ligeramente mayor al de una molécula de agua. Aunque la longitud de esta restricción es solo la de un aminoácido, ello bastaría para bloquear el paso de iones y otros solutos más grandes, ya que no existe la estructura necesaria para despojarlos de sus capas de hidratación.
]]>
La barrera para el paso de protones está constituida por un fuerte dipolo formado por dos segmentos proteicos que contienen la secuencia asparagina-prolina-alanina (NPA), que reorienta las moléculas de agua interrumpiendo las interacciones entre una molécula y la siguiente, lo cual elimina la posibilidad del transporte simultáneo de protones (10). Las dos asparaginas del triplete NPA contienen residuos polares para la formación de puentes de hidrógeno y producen una reorientación transitoria del dipolo de la molécula de agua. Así, las moléculas de agua que penetran en el poro disponen sus oxígenos hacia abajo, pero al interactuar con ambas asparaginas giran y continúan su paso con el oxígeno mirando hacia arriba (Figura 2). En la zona más estrecha del canal existe una cisteína cuyo grupo sulfhidrilo determina la sensibilidad a los iones mercurio (3).
Las acuaporinas en las plantas
Los estudios acerca de las acuporinas proporcionan perspectivas únicas, en múltiples aspectos de la biología vegetal. Primeramente las acuaporinas despertaron un interés considerable debido a la actividad como canal de agua. Este descubrimiento era inesperado en plantas y cambió el conocimiento sobre el transporte de agua a través de la membrana (11, 12), lo que condujo a los investigadores a revisar muchos aspectos de las relaciones hídricas de las plantas y a relacionar esos aspectos con el nuevo contexto fisiológico. Actualmente se ha comprobado que estas proteínas son más que canales de agua (13), pues transportan, además, sustratos de gran significado fisiológico. Estudios de expresión en ovocitos de Xenopus laevis también muestran que, similar a como ocurre en acuagliceroporinas en bacterias y animales, algunas isoformas de acuaporinas vegetales pueden transportar pequeños solutos neutros tales como glicerol (14), urea (15), formamida, acetamida (16), metilamonio (17), ácido bórico (18), ácido silícico (19), o ácido láctico (20). El transporte de amoníaco (NH3), amonio (NH4+) y CO2, también se ha detectado (17, 21).
Durante los últimos años se han caracterizado AQP en más de 30 especies vegetales, incluyendo mono y dicotiledóneas. Como en los animales, presentan una estructura tetramérica y cada monómero posee un poro funcional individual; algunas se inhiben por mercuriales. Su gran abundancia en plantas, comparada con las de los animales, sugiere varias posibilidades. Una de ellas es que las AQP en plantas se puedan encontrar distribuidas en membranas de los diferentes compartimentos celulares (mitocondria, cloroplasto, peroxisoma, retículo endoplásmico, complejo de Golgi y diferentes tipos de vacuolas y vesículas). La presencia de AQP en los diferentes compartimentos pudiera ser el mecanismo que las células utilizan para regular los cambios en el potencial osmótico que sufren durante los diferentes tipos de estrés a los que las plantas se ven continuamente sometidas. Una segunda posibilidad sería que la expresión de algunas AQP puede depender, ya sea del estado de desarrollo de la planta, o de las condiciones ambientales a las que está expuesta (22).
Se ha encontrado que las acuaporinas en las plantas exhiben una marcada multiplicidad de isoformas, lo que confirma lo planteado anteriormente. La secuenciación del genoma estableció que el número de genes de acuaporinas en Arabidopsis es 35 (23, 24) y 33 en arroz (Oryza sativa) (25). Además se han identificado 36 isoformas en maíz (Zea mays L.) (26) y 37 en tomate (Solanum lycopersicum L.) (27).
]]>
Teniendo en cuenta la homología en la secuencia, las acuaporinas de las plantas se pueden subdividir en cuatro subgrupos que se corresponden con las diferentes localizaciones celulares (28). Las proteínas intrínsecas del tonoplasto (PIT) y las proteínas intrínsecas de la membrana plasmática (PIP) corresponden a acuaporinas que se expresan de forma abundante en la membrana vacuolar y plasmática, respectivamente. Las PIP se subdividen en dos subgrupos filogenéticos PIP1 y PIP2. Debido a su abundancia, las PIPs y las PITs representan vías centrales para el transporte de agua transcelular e intracelular (29). Un tercer grupo comprende las proteínas intrínsecas de membrana semejantes a Nodulina26 (PIN) por ejemplo, que presentan homología con GmNod26, una acuaporina abundante en la membrana peribacteroide de los nódulos simbióticos fijadores de nitrógeno de raíces de soya (Glycine max Merr.) (30). Las PINs están presentes en plantas no leguminosas, donde se localizan en la membranas plasmática e intracelulares (31). El cuarto grupo de acuaporinas vegetales está integrado por las proteínas pequeñas intrínsecas básicas (SIP: small basic intrinsic proteins), (32) y se localizan en su mayoría en el retículo endoplasmático (33).
Aunque las cuatro clases de acuaporinas se conservan entre las especies de plantas, esta familia genética muestra signos de reciente y rápida evolución (25). Además algunas especies de plantas han adquirido adicionales y nuevos tipos de acuaporinas. Por ejemplo, un homólogo del GipF (bacterial glicerol facilitator) ha sido adquirido, mediante transferencia horizontal de genes (34), por el musgo Physcomitrella patens, y el genoma de este organismo y el de algunas plantas superiores codifican una quinta clase de acuaporinas, que están estrechamente relacionadas, pero claramente diferente a las PIPs.
Los estudios sobre la función integral de las acuaporinas en plantas se han visto limitados de alguna manera, por la elevada diversidad genética en estos organismos, la carencia de inhibidores específicos y dificultades metodológicas para medir el transporte de agua. Sin embargo existen evidencias sustanciales, basadas en la alteración de las funciones de las acuaporinas por el mercurio o la genética reversa, que muestran que las acuaporinas contribuyen significativamente al transporte de agua en las raíces (35) y en tejidos internos de hojas que transpiran (36).
El transporte de agua mediado por las acuaporinas es también crucial durante el movimiento de la hoja y de los pétalos (37), la reproducción (38), la elongación celular (39) y la germinación de la semillas (40). También se ha propuesto el papel de las acuaporinas intracelulares en la osmorregulación de las células vegetales (41). Se ha informado que las acuaporinas participan en el balance y la eficiencia en el uso del agua en las plantas (42, 28).
Regulación de la función de las acuaporinas
]]>
Las modificaciones post-traduccionales son fundamentales en la regulación de la estructura y función de las proteínas, y por tanto para modular y controlar la actividad de las mismas, su localización subcelular, su estabilidad y su interacción con otras proteínas. Se ha identificado residuos de serina en los extremos N terminal y C terminal, de varias acuaporinas de las plantas (43). En particular, se identificaron dos sitios de fosforilación en el extremo C terminal de las acuaporinas de Arabidopsis AtPIP2;1 (44) y At-PIP2;6 (45) y de espinaca Spinacea oleracea SoPIP2;1 (46). Además, todas las AtPIPs muestran un sitio conservado de fosforilación en el lazo B (47). Basado en el análisis funcional en Xenopus laevis se propuso que la fosforilación de SoPIP2;1 en este sitio y en la Ser262 (en el extremo C terminal) fue capaz de regular su actividad de transporte de agua (47).
La espectrometría de masa y otros adelantos recientes en la proteómica de membrana se han utilizado en la descripción de los patrones de modificación co- y post-traduccional en acuaporinas (28). Dichos estudios confirman que las acuaporinas de membrana plasmática en Arabidopsis pueden ser fosforiladas en múltiples sitios del extremo C-terminal. También se reveló que el extremo N-terminal de PIP2s pueden ser metilados en sitios adyacentes. En particular, la Lys3 y el Glu6 en AtPIP2 llevan una dimetilación y una monometilación, respectivamente (48). Este es el primer informe de proteínas de membrana metiladas en plantas.
Estudios de marcaje in vivo e in vitro, experimentos con anticuerpos antifosfopéptido y análisis de espectrometría de masa han proporcionado evidencias directas de la fosforilación también de residuos de Ser en los extremos N y C terminal de las acuaporinas de Phaseolus vulgaris PvTIP3;1 y GmNOD26 (43, 49). Se han purificado proteínas quinasas dependientes de calcio que actúan sobre las acuaporinas (50, 51). Aunque la mayoría de las acuaporinas de las plantas no muestran glicosilación, este tipo de modificación se observó en GmNOD26 (52).
La actividad de las acuaporinas puede ser afectada por el estado oligomérico de la proteína. Estas se encuentran generalmente de forma tetramérica, tanto in vitro como in vivo. La tetramerización involucra interacciones entre los monómeros vecinos, a través de las a-helices y los lazos extramembrana, los cuales contribuyen a la estabilidad del tetrámero (53). Algunos experimentos muestran que varias acuaporinas forman homotetrameros estables (54, 55, 56). Sin embargo, varias isoformas de las plantas forman heterotetrámeros (53).
También se ha informado la regulación de la permeabilidad de las acuaporinas PIPs por el calcio y el pH (57, 58, 59). La permeabilidad de la membrana plasmática al agua, en suspensiones celulares de Arabidopsis, o en células de la raíz, se reduce en presencia de Ca2+ y bajo pH (57). La acidosis citosólica se observa durante la falta de oxígeno o anoxia resultante del anegamiento del suelo y se asocia a la reducción de la permeabilidad al agua de las células de la raíz (58). Resulta interesante que en Xenopus laevis las acuaporinas de Arabidopsis AtPIP2;1, At-PIP2;2, AtPIP2;3 y AtPIP1;2, se cierran después de un cambio del pH citosólico de siete a seis. Se plantea que el residuo responsable de este tipo de regulación en el cierre de AtPIP2;2 es la His 197, que se localiza en el lazo D intracelular. La sustitución de la His 197 por un residuo de Ala, reduce el efecto de la acidificación citosólica (58).
]]>
Por otro lado, las acuaporinas en las plantas son destinadas específicamente a ciertas membranas. Estudios de inmunocitoquímica e inmunodetección han permitido determinar que la mayoría de las acuaporinas están localizadas tanto en la membrana plasmática, la membrana vacuolar o la membrana peribacteroide de los nódulos fijadores de nitrógeno de la raíz.
Para arribar a este destino, las acuaporinas deben ser transportadas a través de vías secretoras que incluyen el retículo endoplasmático, el complejo de Golgi y dentro de diferentes tipos de vesículas, de acuerdo a la membrana blanco. La regulación del tráfico de las acuaporinas entre membranas, por tanto pudiera representar un mecanismo básico para modular la permeabilidad de las membranas al agua en respuesta a diferentes condiciones ambientales y de disponibilidad de agua (53).
Otro mecanismo de regulación de la actividad de las acuaporinas, pudiera ser las fuerzas de cohesión / tensión en presencia de altas concentraciones de solutos osmóticos (60). Acorde a este modelo, los osmolitos excluidos del canal pudieran causar tensión (presiones negativas) dentro del poro e inducir una deformación de la proteína y eventualmente su cierre.
En conjunto, estos estudios muestran que en cada acuaporina de las plantas pueden ocurrir diversas modificaciones, lo que refleja posiblemente los complejos mecanismos que participan en la regulación de su actividad. Aunque se han hecho significativos progresos, se requiere un mejor conocimiento de la expresión de las mismas en compartimentos subcelulares específicos y en los diferentes tipos celulares (28).
Papel de las acuaporinas en la respuesta de la planta al estrés abiótico
]]>
Las plantas necesitan continuamente ajustar su estatus acuoso en respuesta a las condiciones cambiantes del medio y las acuaporinas juegan un papel importante en estos procesos (61, 62). En particular, estudios genéticos y fisiológicos, han proporcionado evidencias del papel de las acuaporinas en la regulación, en respuesta al estrés abiótico, del transporte de agua de la raíz y de la conductividad hidráulica de la misma (Lpr) (62, 63). Por ejemplo la exposición de plantas de Arabidopsis a la salinidad (100 mM NaCl) indujo una rápida (tiempo promedio de 45 minutos) y significativa disminución (-70 %) en la Lpr, que fue mantenida al menos 24 horas (64). Mientras que el efecto a largo plazo del estrés por NaCl se explica por una completa inhibición transcripcional de las acuaporinas, el mecanismo molecular involucrado en la temprana disminución de la Lpr por el NaCl no se ha explicado todavía completamente. Estos mecanismos involucran un ligero decrecimiento en la cantidad total de las proteínas AtPIP1, a los 30 minutos después de la exposición al NaCl, y un transporte de isoformas AtPIP1 y AtPIP2 entre la membrana plasmática y los compartimentos celulares, que pudiera contribuir a reducir la abundancia de AtPIPs en la membrana plasmática y por tanto de la conductividad hidráulica de las células de la raíz estresada por salinidad (65).
La congelación es otro tipo de estrés abiótico que inhibe la Lpr, y en este contexto se estableció la relación entre la regulación de las acuaporinas y las especies reactivas del oxígeno (66). En pepino (Cucumis sativus L.), por ejemplo, el peróxido de hidrogeno (H2O2) acumulado en respuesta a la congelación y el tratamiento de las raíces con H2O2, inhibe la Lpr en igual magnitud que la congelación. En Arabidopsis también se observó una disminución rápida en la Lpr en respuesta a 2 mM de H2O2 (28). Debido a su extensión (> 70 %) y rapidez (tiempo medio alrededor de ocho minutos), esta disminución se debe indudablemente a la inhibición de las acauporinas de las raíces.
La halófita Mesembryanthemum crystallinum se ha tomado como modelo de estudio, para conocer el papel de las AQP’s durante la exposición de las plantas a la salinidad y el estrés osmótico. De las cuatro AQPs que se han estudiado hasta el momento (McPIP1;4, McPIP1;2, McTIP1;2 y McPIP2;1), se observó que una de ellas, McPIP2;1, sólo se expresa en la raíz, específicamente, en la epidermis y en la endodermis. Además, se constató un aumento en los niveles de proteína de esta acuaporina en respuesta al estrés salino. Estos resultados sugieren que McPIP2;1 tiene un papel importante en el transporte de agua en las raíces y por consiguiente en la adaptación de la planta a la salinidad. Para comprender mejor la participación de la acuaporina McPIP2;1 en la adaptación de la planta a la salinidad esta AQP se ha estudiado con mayor detalle desde el punto de vista funcional, de distribución y de regulaciónA.
Numerosos estudios se han dirigido a comprender los factores que participan en la regulación de la actividad y expresión de las TIP. La actividad de las TIP se regula por señales del desarrollo así como ambientales, tanto a nivel transcripcional como post-transcripcional. Otros estudios han mostrado que la expresión de las TIP se regula por la salinidad (67). Análisis sistemáticos de la expresión de las TIP, en respuesta al estrés abiótico se han llevado a cabo en Arabidopsis y maíz (Z. mays) (68, 69). Los resultados indican que la mayoría de las TIP fueron reprimidas por la salinidad y la sequía en ambos cultivos. En arroz (Oryza sativa L.) la expresión de OsTIP1;1 se incrementó bajo estrés salino (70). La correlación entre los niveles de ARNm de las TIP y el volumen de savia circulante sugiere una relación entre la toma de agua por las raíces y la expresión de las TIP (25).
Se ha informado sobre la caracterización de la acuaporina TaNIP (Triticum asetivum L. nodulin 26-like intrinsic protein) (71). Este gen se identificó en el mutante de trigo tolerante a la salinidad RH8706-49 sometido a estrés salino. La sobreexpresión de este gen en Arabidopsis permitió una mayor tolerancia a la salinidad que las plantas silvestres. Las plantas transgénicas acumularon mayor cantidad de K+, Ca++ y prolina y menor contenido de Na+ en comparación con el fenotipo silvestre. También en las plantas transformadas se observó la activación de la expresión de otros genes asociados al estrés. Se demostró que la acuaporina TaNIP se expresa en la membrana plasmática.
]]>
La mayoría del transporte simplástico de agua en las plantas ocurre vía acuaporinas pero no se conoce hasta qué punto contribuyen al estatus acuoso de la planta bajo condiciones favorables y bajo estrés abiótico. Debido a ello se ha sobreexpresado la acuaporina de membrana plasmática de Arabidopsis PIP1b, en plantas de tabaco transgénicas (72). En condiciones favorables de crecimiento, la sobreexpresión de PIP1b incrementó de forma significativa la tasa de crecimiento de las plantas, la tasa de transpiración, la densidad estomática y la eficiencia fotosintética. Por el contrario, la sobreexpresión de PIP1b no tuvo un efecto beneficioso bajo estrés salino. Estos autores sugieren que el transporte de agua a través de la vía simplástica de las acuaporinas representa un factor limitante para el crecimiento y vigor de las plantas en condiciones favorables, mientras que el incremento de este tipo de transporte, no tiene un efecto beneficioso bajo estrés salino.
Por otro lado la expresión de algunos genes que codifican acuaporinas de la membrana plasmática, tales como el gen RD28 de Arabidopsis y NeMip2 y NeMip3 de Nicotiana excelsior, se estimulan bajo estrés por sequía (73, 74). En contraste la expresión del gen MIPA, que codifica para una acuaporina de la membrana plasmática en Mesembryanthemum crystallinum es reprimida bajo estrés por salinidad (74), mientras que la expresión del gen PIP1b de Arabidopsis no se altera de forma significativa en condiciones estresantes (75). La reducción de los niveles de expresión del gen PIP1a en Arabidopsis por medio de una construcción antisentido bajo un promotor 35S, resultó en un cambio significativo en la morfología de la raíz y un incremento en la masa de la misma (76); sin embargo, no se afectó el potencial hídrico ni el peso de la planta. En otras investigaciones se redujeron los niveles de expresión de un homólogo de PIP1b en tabaco (NtAQP1) (35), por un método similar a los autores anteriores. Esto resultó en una mayor sensibilidad al polietilénglicol, menor conductividad hidráulica de la raíz y disminución en la tasa de transpiración, que en aquellas plantas que no fueron transformadas, bajo estrés por sequía.
Durante el estrés osmótico se han informado aumentos de la expresión de AQP en diferentes tejidos y especies vegetales, como plántulas y partes aéreas de A. thaliana, tallos de tomate, anteras e inflorescencia del coliflor, raíces y partes aéreas del arroz y hojas y raíces de girasol. En contraste, se ha observado que el estrés osmótico causa una disminución de la expresión de AQP en hojas, raíces y tallos de Nicotiana glauca y hojas de Mesembryanthemum crystallinum. Trabajos recientes apoyan el papel de algunas AQP en la tolerancia al estrés hídrico. Se ha observado que la represión de la expresión de AQP resulta en cambios en la conductividad hidráulica de las raíces, velocidad de transpiración, permeabilidad osmótica y capacidad de las plantas para recuperarse en suelos deficientes en agua (22).
En la halófita M. crystallinum se ha observado que existe una regulación diferencial en los niveles de expresión de las diferentes AQP por estrés salino y por estrés osmótico. Durante el estrés salino se reduce la expresión de AQP en hojas, mientras que se incrementa en la raíz. El estrés osmótico causa un aumento de AQP en hojas y raíces, que contrasta con la disminución causada por el estrés salino. Estos hallazgos sugieren que la planta es capaz de discriminar exactamente entre el estrés iónico y el estrés osmótico, posiblemente a través de diferentes mecanismos de transducción de señales involucrados en la adaptación a estos tipos de estrés (3, 22).
Otros autores evaluaron los niveles de ARNm de la acuaporina HtPIP2;4, además de algunos indicadores fisiológicos (77), en condiciones de salinidad, en dos líneas de cebada (Hordeum vulgare L.), una tolerante y otra resistente. Obtuvieron que el tratamiento salino disminuyó más la transpiración y la conductividad hidráulica, en la línea sensible que en la tolerante. Este efecto fue paralelo a la más rápida y persistente inhibición de la acuaporina, en el cultivar sensible que en el tolerante, por lo que concluyeron que los cambios en los niveles de expresión de las acuaporinas están implicados en el control de la conductividad hidráulica de la planta completa y en la regulación de las relaciones hídricas en el tallo.
]]>
A nivel de la planta completa el principal efecto de la sequía es la reducción de la transpiración a través del cierre estomático. No obstante, en condiciones extremas, la elevada tensión en el xilema puede conducir a la oclusión por embolización. Algunos autores propusieron que las acuaporinas tienen una función específica en el relleno del embolismo y la recuperación de la conductancia axial del tallo después de la sequía, en experimentos de inhibición con mercurio, en Vitis vinifera L. (78) Resultados similares se obtuvieron en la especie Populus trichocarpa (79).
Por otro lado, en Juglans nigra L., una especie maderable, los transcriptos y las proteínas correspondientes a dos homólogos de las PIP2, mostraron variaciones estacionales y se acumularon en el parénquima xilemático durante las bajas temperaturas del invierno, lo que sugiere también una función específica de las acuaporinas en la reparación del embolismo; teniendo en cuenta que esta se lleva a cabo por la hidrólisis del almidón en las células parenquimáticas adyacentes y la exudación de los azúcares resultantes dentro de la vesícula. El influjo de agua concomitante trae consigo que se rompa la burbuja de aire y se repare el embolismo (80).
El anegamiento de los suelos conlleva a una aguda falta de oxígeno de las raíces de las plantas, lo que impone un fuerte estrés a las mismas. Otros autores delinearon las bases celulares de este proceso en raíces de Arabidopsis como se mostró en el capítulo sobre la regulación de la función de las acuaporinas (58). Se ha planteado además, que este tipo de regulación puede evitar la excesiva dilución de la savia xilemática después de la inundación o en períodos posteriores favorecer la acumulación del etileno, lo que en cambio induce la diferenciación del aerénquima (81). En Arabidopsis, ocurre una represión general de los genes de las PIP y las TIP en respuesta a la hipoxia. Por el contrario, la expresión de AtNIP2;1 se induce marcadamente después de la inundación y la hipoxia (82). La acuaporina AtNIP2;1, transporta ácido láctico y puede, por tanto, ser una vía para la liberación de los productos de la fermentación de las células radicales para contribuir a la regulación del pH y la adaptación metabólica a la hipoxia por un período prolongado (20).
Es ampliamente conocida la función del ácido abscísico (ABA) en la respuesta de la planta frente a la sequía en la inducción del cierre estomático y otros mecanismos de defensa de la planta (83). Se ha sugerido que esta hormona influye en la conductancia hidráulica de la planta, regulando la actividad y la expresión de las acuaporinas (isoformas PIPs) en líneas transgénicas de maíz, por lo que contribuye a mantener el estatus hídrico de la planta en condiciones de sequía (84).
Por otro lado se informó que la expresión de AtPIP2;2, AtPIP2;4, AtPIP2;5, y AtPIP2;7, que son permeables al H2O2 en células de levaduras, fueron moduladas diferencialmente por los diferentes tipos de estrés abiótico (85). En particular, la expresión de AtPIP2;2, AtPIP2;4 y AtPIP2;7, fue inhibida por la aplicación de H2O2 y por la sequía. Los autores concluyeron que la regulación concomitante en la expresión de ciertos miembros de la familia de acuaporinas puede jugar un papel importante en la respuesta de la planta mediada por el H2O2, frente a condiciones ambientales cambiantes. Mientras que otros autores también relacionaron el efecto del H2O2, con la regulación de las acuaporinas y el control de las propiedades hidráulicas de la raíz en Phaseolus vulgaris (86).
]]>
Consideraciones finales
Las acuaporinas son los únicos transportadores moleculares de agua en las plantas y su estudio ha mejorado significativamente el conocimiento sobre los mecanismos integrados de transporte de agua, en las raíces, en particular. Sin embargo, en vistas de los complejos mecanismos de expresión y regulación de las acuaporinas, su papel en la regulación del transporte de agua y en muchos otros procesos fisiológicos, que incluyen la germinación de las semillas, regulación de los estomas y el transporte de agua en las hojas, merecen investigaciones futuras.
Otro desafío futuro es comprender cómo las acuaporinas pueden ser específicamente situadas en subdominios específicos de las membranas en las células vegetales y cómo esto contribuirá a su especialización funcional. El mecanismo celular que determina el transporte de las acuaporinas y su relocalización celular en respuesta a los diferentes estímulos será, sin lugar a dudas, objeto de intensas investigaciones en los próximos años.
Por último, las funciones de las acuaporinas deben ser integradas a la fisiología de la planta como un todo. Esto requiere una mejor comprensión de cómo las varias actividades de transporte de las acuaporinas se acoplan con aquellas de otras proteínas transportadoras. También debe ser esclarecida la cadena de eventos que conllevan al control de las funciones de las acuaporinas mediante señales locales o a larga distancia, durante el desarrollo o en respuesta a señales de estrés biótico y abiótico.
REFERENCIAS
1. Du, S.; Dai, Q.; Chen, C.; Liu, T.; Tian, X.; Gong, Y.; Sun, Y. y Wang, J. Molecular cloning and characterization of the plasma membrane aquaporin gene (OvPIP) from Orychophragmus violaceus. African Journal of Biotechnology, 2011, vol. 10, no. 19, pp. 3709-3714.
]]>
2. Maurel, C.; Verdoucq, L.; Luu, D. y Santoni, V. Plant Aquaporins: membrane channels with multiple integrated functions. Annu. Rev. Plant Biol., 2008, vol. 59, pp. 595-624.
3. Coppo, J. A. Acuaporinas. Rev. Vet., 2008, vol. 19, no. 2, pp. 167-178.
4. Maurel, C.; Kado, R. T.; Guern, J. y Chrispeels, M. J. Phosphorylation regulates the waterchannel activity of the seed-speci?c aquaporin a-TIP. EMBO J., 1995, vol. 14 pp. 3028-3035.
5. Agre, P. Molecular physiology of water transport: aquaporin nomenclature workshop. Mammalian aquaporins. Biol. Cell., 1997, vol. 89, pp. 255-257.
6. Jung, J. S.; Preston, G. M.; Smith, B. L.; Guggino, W. B. y Agre, P. Molecular structure of the water channel through aquaporin CHIP: the hourglass model. J. Biol. Chem., 1994, vol. 269, pp. 14648-14654.
]]>
7. Vanos, C. H.; Deen, P. M. y Dempster, J. A. Aquaporins: water selective channels in biological membranes. Biochim Biophys Acta, 1994, vol. 1197, pp. 291-309.
8. De Groot, B. L.; Engel, A. y Gubmuller, H. A refined structure of human aquaporin-1. FEBS Letters, 2001, vol. 504, pp. 206-211.
9. Waltz, T.; Smith B. L.; Zeidel, M. L.; Engel, A. y Agre, P. Biologically active two-dimensional crystals of aquaporin CHIP. J .Biol. Chem., 1994, vol. 269, pp. 1583-1586.
10. Tamargo, J. Los poros y los canales iónicos regulan la actividad celular. An. R. Acad. Nac. Farm., 2004, vol. 70, p. 9-31.
11. Schaffner, A. R. Aquaporin function, structure, and expression: are there more surprises to surface in water relations? Planta, 1998, vol. 204, pp. 131-139.
]]>
12. Tyerman, S. D.; Bohnert, H. J.; Maurel, C.; Steudle, E. y Smith, J. A. Plant aquaporins: their molecular biology, biophysics and significance for plant water relations. J. Exp. Bot., 1999, vol. 50, pp. 1055-1071.
13. Tyerman, S. D.; Niemietz, C. M. Plant aquaporins: multifunctional water and solute channels with expanding roles. Plant Cell Environ., 2002, vol. 25, pp. 173-194.
14. Biela, A.; Grote, K.; Otto, B.; Hoth, S.; Hedrich, R. y Kaldenhoff, R. The Nicotiana tabacum plasma membrane aquaporin NtAQP1 is mercury-insensitive and permeable for glycerol. Plant J., 1999, vol. 18, pp. 565-570.
15. Gerbeau, P.; Guclu, J.; Ripoche, P. y Maurel, C. Aquaporin Nt-TIPa can account for the high permeability of tobacco cell vacuolar membrane to small neutral solutes. Plant J., 1999, vol. 18, p. 577-587.
16. Rivers, R. L.; Dean, R. M.; Chandy, G.; Hall, J. E.; Roberts, D. M. y Zeidel, M. L. Functional analysis of Nodulin 26, an aquaporin in soybean root symbiosomes. J. Biol. Chem., 1997, vol. 272, pp. 16256-16261.
]]>
17. Holm, L. M.; Jahn, T. P.; Moller, A. L.; Schjoerring, J. K. y Ferri, D. NH3 and NH4+ permeability in aquaporin-expressing Xenopus oocytes. Pflugers Arch., 2005, vol. 450, pp. 415-428.
18. Takano, J.; Wada, M.; Ludewig, U.; Schaaf, G.; von Wiren, N. y Fujiwara, T. The Arabidopsis major intrinsic protein NIP5;1 is essential for efficient boron uptake and plant development under boron limitation. Plant Cell., 2006, vol. 18, pp. 1498-1509.
19. Ma, J. F.; Tamai, K.; Yamaji, N.; Mitani, N. y Konishi, S. A silicon transporter in rice. Nature, 2006, vol. 440, pp. 688-691.
20. Choi, W. G. y Roberts, D. M. Arabidopsis NIP2;1, a major intrinsic protein transporter of lactic acid induced by anoxic stress. J. Biol. Chem., 2007, vol. 282, pp. 24209-24218.
21. Uehlein, N.; Lovisolo, C.; Siefritz, F. y Kaldenhoff, R. The tobacco aquaporin NtAQP1 is a membrane CO2 pore with physiological functions. Nature, 2003, vol. 425, pp. 734-737.
]]>
22. Vera-Estrella, R.; Barkla, B. J. y Pantoja, O. La biología y la fisiología de las acuaporinas en plantas. Biojournal.net, 2005, vol. 1, pp. 1-15. On line: http://www.biojournal.net/pdf/05.pdf.
23. Johanson, U.; Karlsson, M.; Gustavsson, S.; Sjovall, S.; Fraysse, L.; Weig, A. R. y Kjellbom, P. The complete set of genes encoding Major Intrinsic Proteins in Arabidopsis provides a framework for a new nomenclature for major intrinsic proteins in plants. Plant Physiol., 2001, vol. 126, pp. 1358-1369.
24. Quigley, F.; Rosenberg, J. M.; Shachar-Hill, Y. y Bohnert, H. J. From genome to function: the Arabidopsis aquaporins. Genome Biol., 2001, vol. 3, pp. 1-17.
25. Sakurai, J.; Ishikawa, F.; Yamaguchi, T.; Uemura, M. y Maeshima, M. Identification of 33 rice aquaporin genes and analysis of their expression and function. Plant Cell Physiol., 2005, vol. 46, pp. 1568-1577.
26. Chaumont, F.; Barrieu, F.; Wojcik, E.; Chrispeels, M. J. y Jung, R. Aquaporins constitute a large and highly divergent protein family in maize. Plant Physiol., 2001, vol. 125, pp. 1206-1215.
]]>
27. Sade, N.; Vinocur, B. J.; Diber, A.; Shatil, A.; Ronen, G.; Nissan, H.; Wallach, R.; Karchi, H. y Moshelion, M. Improving plant stress tolerance and yield production: is the tonoplast aquaporin SlTIP2;2 a key to isohydric to anisohydric conversion? New Phytol., 2009, vol. 181, pp. 651-661.
28. Maurel, C. Plant aquaporins: novel functions and regulation properties. FEBS Lett., 2007, vol. 581, pp. 2227-2236.
29. Cohen, D.; Bogeat, M. B; Vialet, S.; Merret, R.; Courty, P. E. /et al./. Developmental and environmental regulation of aquaporine gene expression across Populus species: divergency or redundancy? PloS ONE, vol. 8, no. 2, p. E55506. doi:10.1371/journal.pone.0055506.
30. Wallace, I. S.; Choi, W. G. y Roberts, D. M. The structure, function and regulation of the nodulin 26-like intrinsic protein family of plant aquaglyceroporins. Biochim. Biophys. Acta., 2006, vol. 1758, pp. 1165-1175.
31. Mizutani, M.; Watanabe, S.; Nakagawa, T. y Maeshima, M. Aquaporin NIP2;1 is mainly localized to the ER membrane and shows root-specific accumulation in Arabidopsis thaliana. Plant Cell Physiol., 2006, vol. 47, pp. 1420-1426.
]]>
32. Johanson, U. y Gustavsson, S. A new subfamily of major intrinsic proteins in plants. Mol. Biol. Evol., 2002, vol. 19, pp. 456-461.
33. Ishikawa, F.; Suga, S.; Uemura, T.; Sato, M. H. y Maeshima, M. Novel type aquaporin SIPs are mainly localized to the ER membrane and show cell-specific expression in Arabidopsis thaliana. FEBS Lett., 2005, vol. 579, pp. 5814-5820.
34. Gustavsson, S.; Lebrun, A.; Norden, K.; Chaumont, F. y Johanson, U. A novel plant major intrinsic protein in Physcomitrella patens most similar to bacterial glycerol channels. Plant Physiol., 2005, vol. 139, pp. 287-295.
35. Siefritz, F.; Tyree, M. T.; Lovisolo, C.; Schubert, A. y Kaldenhoff, R. PIP1 plasma membrane aquaporins in tobacco: from cellular effects to function in plants. Plant Cell., 2002, vol. 14, pp. 869-876.
36. Cochard, H.; Venisse, J. S.; Barigah, T. S.; Brunel, N.; Herbette, S.; Guilliot, A.; Tyree, M. T. y Sakr, S. New insights into the understanding of variable hydraulic conductances in leaves. Evidence for a possible implication of plasma membrane aquaporins. Plant Physiol., 2007, vol. 143, pp. 122-133.
]]>
37. Azad, A. K.; Sawa, Y.; Ishikawa, T. y Shibata, H. Phosphorylation of plasma membrane aquaporin regulates temperature-dependent opening of tulip petals. Plant Cell Physiol., 2004, vol. 45, pp. 608-617.
38. Bots, M.; Vergeldt, F.; Wolters-Arts, M.; Weterings, K.; van As, H. y Mariani, C. Aquaporins of the PIP2 class are required for efficient anther dehiscence in tobacco. Plant Physiol., 2005, vol. 137, pp. 1049-1056.
39. Hukin, D.; Doering-Saad, C.; Thomas, C. R. y Pritchard, J. Sensitivity of cell hydraulic conductivity to mercury is coincident with symplasmic isolation and expression of plasmalemma aquaporin genes in growing maize roots. Planta, 2002, vol. 215, pp. 1047-1056.
40. Vander, C.; Postaire, O.; Tournaire-Roux, C.; Boursiac, Y. y Maurel, C. Expression and inhibition of aquaporins in germinating Arabidopsis seeds. Plant Cell Physiol., 2006, vol. 47, pp. 1241-1250.
41. Khaled, K.; Karim, K.; Mourad, O.; Abdekarim, K. y Nouredine, K. PIP Aquaporin protein of in durum wheat, homology modelling and 3D structure analysis. Biomirror, 2011, vol. 2, no. 6, pp. 1-7.
]]>
42. Johansson, I.; Karlsson, M.; Johanson, U.; Larsson, C. y Kjellbom, P. The role of aquaporins in cellular and whole plant water balance. Biochimica et Biophysica Acta, 2002, vol. 1465, pp. 324-342.
43. Daniel, J. A. y Yeager, M. Phosphorylation of aquaporin PvTIP3;1 defined by mass spectrometry and molecular modeling. Biochemistry, 2005, vol. 44, pp. 14443-14454.
44. Nuhse, T. S.; Stensballe, A.; Jensen, O. N. y Peck, S. C. Phosphoproteomics of the Arabidopsis plasma membrane and a new phosphorylation site database. Plant Cell., 2004, vol. 16, pp. 2394-2405.
45. Niittyla, T.; Fuglsang, A. T.; Palmgren, M. G.; Frommer, W. B. y Schulze, W. X. Temporal analysis of sucrose-induced phosphorylation changes in plasma membrane proteins of Arabidopsis. Mol. Cell. Proteomics, 2007, vol. 6, pp. 1711-1726.
46. Johansson, I.; Larsson, C.; Ek, B. y Kjellbom, P. The major integral proteins of spinach leaf plasma membranes are putative aquaporins and are phosphorylated in response to Ca and apoplastic water potential. Plant Cell., 1996, vol. 8, p. 1181-1191.
]]>
47. Johansson, I.; Karlsson, M.; Shukla, V. K.; Chrispeels, M. J.; Larsson, C. y Kjellbom, P. Water transport activity of the plasma membrane aquaporin PM28A is regulated by phosphorylation. Plant Cell., 1998, vol. 10, pp. 451-459.
48. Santoni, V.; Verdoucq, L.; Sommerer, N.; Vinh, J.; Pflieger, D. y Maurel, C. Methylation of aquaporins in plant plasma membrane. Biochem. J., 2006, vol. 400, pp. 189-197.
49. Guenther, J. F.; Chanmanivone, N.; Galetovic, M. P.; Wallace, I. S.; Cobb, J. A. y Roberts, D. M. Phosphorylation of soybean nodulin 26 on serine 262 enhances water permeability and is regulated developmentally and by osmotic signals. Plant Cell., 2003, vol. 15, pp. 981-991.
50. Harveng, P.; Vlerick, A.; Fuks, B.; Wattiez, R.; Ruysschaert, J. M. y Homble, F. Lentil seed aquaporins form a hetero-oligomer which is phosphorylated by a Mg2+-dependent and Ca2+-regulated kinase. Biochem. J., 2000, vol. 352, pp. 183-190.
51. Sjovall-Larsen, S.; Alexandersson, E.; Johansson, I.; Karlsson, M.; Johanson, U. y Kjellbom, P. Purification and characterization of two protein kinases acting on the aquaporin SoPIP2;1. Biochim. Biophys. Acta., 2006, vol. 1758, pp. 1157-1164.
]]>
52. Vera-Estrella, R.; Barkla, B. J.; Bohnert, H. J. y Pantoja, O. Novel regulation of aquaporins during osmotic stress. Plant Physiol., 2004, vol. 135, pp. 2318-2329.
53. Chaumont, F.; Moshelion, M. y Daniels, M. Regulation of plant aquaporin activity. Biol. Cell., 2005, vol. 97, pp. 749-764.
54. Fu, D.; Libson, A.; Miercke, L. J.; Weitzman, C.; Nollert, P.; Krucinski, J. y Stroud, R. M. Structure of a glycerol-conducting channel and the basis for its selectivity. Science, 2000, vol. 290, pp. 481-486.
55. Murata, K.; Mitsuoka, K.; Hirai, T.; Walz, T.; Agre, P.; Heymann, J. B.; Engel, A. y Fujiyoshi, Y. Structural determinants of water permeation through aquaporin-1. Nature, 2000, vol. 407, pp. 599-605.
56. Fotiadis, D.; Jeno, P.; Mini, T.; Wirtz, S.; Mulle, S. A.; Fraysse, L.; Kjellbom, P. y Engel, A. Structural characterization of twoaquaporins isolated from native spinach leaf plasma membranes. J. Biol. Chem., 2001, vol. 276, pp. 1707-1714.
]]>
57. Gerbeau, P.; Amodeo, G.; Henzler, T.; Santoni, V.; Ripoche, P. y Maurel, C. The water permeability of Arabidopsis plasma membrane is regulated by divalent cations and pH. Plant J., 2002, vol. 30, pp. 71-81.
58. Tournaire-Roux, C.; Sutka, M.; Javot, H.; Gout, E.; Gerbeau, P.; Luu, D. T.; Bligny, R. y Maurel, C. Cytosolic pH regulates root water transport during anoxic stress through gating ofaquaporins. Nature, 2003, vol. 425, pp. 393-397.
59. Nemeth-Cahalan, K. L. y Hall, J. E. pH and calcium regulate the water permeability of aquaporin. J. Biol. Chem., 2000, vol. 275, pp. 6777-6782.
60. Ye, Q.; Wiera, B. y Steudle, E. A cohesion/tension mechanism explains the gating of water channels (aquaporins) in Chara internodes by high concentration. J. Exp. Bot., 2004, vol. 55, pp. 449-461.
61. Vandeleur, R.; Niemietz, C. M.; Tilbrook, J. y Tyerman, S. D. Role of aquaporins in root responses to irrigation. Plant Soil., 2005, vol. 274, pp. 141-161.
]]>
62. Luu, D. T. y Maurel, C. Aquaporins in a challenging environment: molecular gears for adjusting plant water status. Plant Cell Environ., 2005, vol. 28, pp. 85-96.
63. Javot, H. y Maurel, C. The role of aquaporins in root water uptake. Ann. Bot., 2002, vol. 90, pp. 301-313.
64. Boursiac, Y.; Chen, S.; Luu, D. T.; Sorieul, M.; van den Dries, N. y Maurel, C. Early effects of salinity on water transport in Arabidopsis roots. Molecular and cellular features of aquaporin expression. Plant Physiol., 2005, vol. 139, pp. 790-805.
65. Prak, S.; Hem, S.; Boudet, J.; Viennois, G.; Sommerer, N.; Rossignol, M.; Maurel, C.; Santoni, V. Multiple phosphorylations in the C-terminal tail of plant plasma membrane aquaporins role in subcellular trafficking of AtPIP2;1 in response to salt stress. Molecular & Cellular Proteomics, 2008, vol. 7, no. 6, pp. 1019- 1026.
66. Lee, S. H.; Singh, A. P. y Chung, G. C. Rapid accumulation of hydrogen peroxide in cucumber roots due to exposure to low temperature appears to mediate decreases in water transport. J. Exp. Bot., 2004, vol. 55, pp. 1733-1741.
]]>
67. Kaldenhoff, R. y Fischer, M. Functional aquaporin diversity in plants. Biochim. Biophys. Acta, 2006, vol. 1758, pp. 1134-1141.
68. Alexandersson, E.; Fraysse, L.; Sjovall-Larsen, S.; Gustavsson, S.; Fellert, M. y Karlsson, M. Whole gene family expression and drought stress regulation of aquaporins. Plant Mol.Biol., 2005, vol. 59, pp. 469-484.
69. Zhu, C.; Schraut, D.; Hartung, W. y Schaffner, A. R. Differential responses of maize MIP genes to salt stress and ABA. J. Exp.Bot., 2005, vol. 56, pp. 2971-2981.
70. Liu, Q.; Umeda, M. y Uchimiya, H. Isolation and expression analysis of two rice genes encoding the major intrinsic protein. Plant Mol. Biol., 1994, vol. 26, pp. 2003-2007.
71. Gao, Z.; He, X.; Zhao, B.; Zhou, C.; Liang, Y.; Ge, R.; Shen, Y. y Huang, Z. Overexpressing a putative aquaporin gene from wheat, TaNIP, enhances salt tolerance in transgenic Arabidopsis. Plant Cell Physiol., 2010, vol. 51, no. 5, pp. 767-775.
]]>
72. Aharon, R.; Shahak, Y.; Wininger, S.; Bendov, R.; Kapulnik, Y. y Galili, G. Overexpression of a plasma membrane Aquaporin in transgenic tobacco improves plant vigor under favorable growth conditions but not under drought or salt stress. The Plant Cell., 2003, vol. 15, pp. 439-447.
73. Yamaguchi, K.; Koizumi, M.; Urao, S. y Shinozaki, K. Molecular cloning and characterization of 9 cDNAs for genes that are responsive to desiccation in Arabidopsis thaliana: Sequenceanalysis of one cDNA clone encodes a putative transmembrane channel protein. Plant Cell Physiol., 1992, vol. 33, pp. 217-224.
74. Yamada, S.; Komory, T.; Myers, P. M.; Kuwata, S. y Imaseki, H. Expression of plasma membrane water channel genesunder water stress in Nicotiana excelsior. Plant Cell Physiol., 1997, vol. 38, pp. 1226-1231.
75. Grote, K.; Trzebiatovski, P. y Kaldenhoff, R. RNA levels of plasma membrane aquaporins in Arabidopsis thaliana. Protoplasma, 1998, vol. 204, pp. 139-144.
76. Kaldenhoff, R.; Grote, K.; Zhu, J. J. y Zimmermann, U. Significance of plasmalemma aquaporins for water transport in Arabidopsis thaliana. Plant J., 1998, vol. 14, pp. 121-128.
]]>
77. Vysotskaya, V.; Hedley, P.; Sharipova, G.; Veselov, D.; Kudoyarova, G.; Morris, J. y Jones, H. Effect of salinity on water relations of wild barley plants differing in salt tolerance. AoB PLANTS, vol. 2010, p. 006, doi:10.1093/aobpla/plq006.
78. Lovisolo, C. y Schubert A. Mercury hinders recovery of shoot hydraulic conductivity during grapevine rehydration: evidence from a whole-plant approach. New Phytol., 2006, vol. 172, pp. 469-478.
79. Secchi F. y Zwieniecki, M. A. Patterns of PIP gene expression in Populus trichocarpa during recovery from xylem embolism suggest a major role for the PIP1 aquaporin subfamily as moderators of refilling process. Plant Cell and Environment, 2010, vol. 33, p. 1285-1297.
80. Sakr, S.; Alves G.; Morillon, R.; Maurel, K. y Decourteix, M. Plasma membrane aquaporins are involved in winter embolism recovery in walnut tree. Plant Physiol., 2003, vol. 133, pp. 630-641.
81. Holbrook, N. M. y Zwieniecki, M. A. Plant biology: water gate. Nature, 2003, vol. 425, p. 361.
]]>
82. Liu, F.; Vantoai, T.; Moy, L. P.; Bock, G.; Linford, L. D. y Quackenbush, J. Global transcription pro?ling reveals comprehensive insights into hypoxic response in Arabidopsis. Plant Physiol., 2005, vol. 137, pp. 1115-1129.
83. Chávez, L.; Fonseca, A. y Ramírez, R. Apuntes sobre algunos reguladores del crecimiento vegetal que participan en la respuesta de las plantas frente al estrés abiótico. Cultivos Tropicales, 2012, vol. 33, no. 3, pp. 47-56.
84. Parent, B.; Hachez, C.; Redondo, E.; Simonneau, P.; Chaumont, S. y Tardieu, F. Drought and Abscisic Acid effects on aquaporin content translate into changes in hydraulic conductivity and leaf growth rate: a trans-scale approach. Plant Physiology, 2009, vol. 149, pp. 2000-2012.
85. Young, J.; Ye, J.; Chae, G. y Hunseung, G. Aquaporin as a membrane transporter of hydrogen peroxide in plant response to stresses. Plant Signaling & Behavior, 2012, vol. 7, no. 9, pp. 1180-1181.
86. Benabdellah, K.; Ruiz-Lozano J. M. y Aroca, R. Hydrogen peroxide effects on root hydraulic proper-ties and plasma membrane aquaporin regulation in Phaseolus vulgaris. Plant. Mol. Biol., 2009, vol. 70, pp. 647-651.
Recibido: 15 de febrero de 2013
Aceptado: 17 de mayo de 2013
Ms.C. Licet Chávez Suárez, Investigador Agregado de la Dirección de Investigaciones y Servicios Ambientales, Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, carretera vía Manzanillo km 17, Bayamo, Granma, Cuba. Email: licet@dimitrov.cu
]]>{kind=link}
{kind=link}