{kind=link}
{kind=link}
Seed germination and pruning effect in the posttransplant establishment of Palma Kerpis [Veitchia merrillii (Becc.), H. E. Moore, Arecaceae]
Dr. Jesús Mao Estanislao Aguilar Luna,I Francisco Domínguez Hernández,II Juan M. Barrios Díaz,III Fabián Enríquez GarcíaIII
IProfesor Investigador, Universidad de la Cañada, Oaxaca, México.
IIProfesor-Investigador del Programa de Ingeniería Agroforestal, Facultad de Ingeniería Agrohidráulica, Benemérita Universidad Autónoma de Puebla, Puebla, México.
IIIProfesores-Investigadores del Programa de Ingeniería Agrohidráulica, Facultad de Ingeniería Agrohidráulica, Benemérita Universidad Autónoma de Puebla, Puebla, México.
]]>
RESUMEN
Estudios de germinación y establecimiento postrasplante de Palma Kerpis son escasos. Por ello se realizaron dos experimentos, uno para estudiar el efecto del tamaño del fruto en la germinación y otro para evaluar el efecto de la poda radical y aérea en el establecimiento postrasplante. El primer ensayo en un diseño CA, se realizó en cámaras de germinación con tres tamaños de fruto: chico (1,34 ± 0,10 y 2,61 ± 0,21 cm), mediano (1,52 ± 0,18 y 3,02 ± 0,31 cm) y grande (2,08 ± 0,14 y 3,57 ± 0,26 cm), en diámetro ecuatorial y polar, respectivamente. El segundo ensayo en un diseño BA, con un arreglo factorial de dos factores (palmas con hojas y sin hojas), cinco niveles (0, 25, 50, 75 y 100 % de raíz) y 10 repeticiones por tratamiento. Los resultados indicaron que las semillas de frutos grandes germinaron mejor, hasta un 95 % a los 28 días; y en la medida que disminuyó el tamaño, también lo hizo el porcentaje de germinación. A los 18 meses después de la siembra, las palmas provenientes de frutos grandes alcanzaron hasta 69,60 cm en altura de planta, lo cual coincidió con el desarrollo de la primera hoja foliada; así también, a menor tamaño de fruto, menor altura. A los ocho meses después del trasplante, las palmas del tratamiento (con hojas y 75 % de raíz al momento del trasplante), se destacaron en comparación con los otros tratamientos aplicados; mostrando valores sobresalientes en altura de planta (99,81 cm), longitud radical (6894 cm) y ganancia de masa seca (57,21 g).
Palabras clave: infrutescencia, energía germinativa, raíces, área foliar, biomasa.
ABSTRACT
Studies of germination and establishment post-transplant of Palma Kerpis are few. In order to address this issue, two experiments were conducted, one for studying the fruit size effect in the germination and another one to evaluate the effect on the radical and aerial pruning in the post-transplant establishment. The first experiment in a RC design was done in germination chambers with three sizes of fruit: small (1,34 ± 0,10 and 2,61 ± 0,21 cm), medium (1,52 ± 0,18 and 3,02 ± 0,31 cm) and large (2,08 ± 0,14 and 3,57 ± 0,26 cm), in equatorial and polar diameter, respectively. The second experiment in a RCB design, using a factorial of two factors (palms with leaves and palms with leafless), five levels (0, 25, 50, 75 and 100 % of the root) and 10 repetitions per treatment. The results indicated that the large fruits seeds germinate best, up to 95 % at 28 days; and when the size is smaller, so too is the germination percentage. At 18 months after planting, the palms that come from large fruits, reached up to 69,60 cm plant height, which coincided with the development of the first foliated leaf; and also, at smaller size of the fruit, lower height. Eight months after the transplant, the treatment palms (with leaves and 75 % of root at the time of the transplant), highlighted in comparison with the other treatments; showed outstanding values in plant height (99,81 cm), root length (6894 cm) and dry mass gain (57,21 g).
Key words: infrutescence, germinative energy, roots, leaf area, biomass.
INTRODUCCIÓN
]]> La Palma Kerpis [Veitchia merrillii (Becc.), H. E. Moore, Arecaceae], debe su nombre en honor a James Veitch (1792-1893), jardinero británico y a E.D. Merrill (1876-1956), botánico americano. También es llamada palma de Navidad, por corresponder el desarrollo de su infrutescencia roja con la época invernal; o palma de Manila, dado que su origen es Filipinas. Por ser una especie con atributos morfológicos de valor ornamental y fácil reproducción, su utilización para la plantación se ha venido incrementando en años recientes, para enriquecer la estética de parques y jardines, principalmente.
MATERIALES Y MÉTODOS
El área experimental fue en la ciudad de Cancún, Quintana Roo, México (21°09´ LN, 86°49´ LO y 8 m snm). En este lugar, la temperatura media anual es de 27,1 °C y la precipitación anual es de 1337,7 mm. Se realizaron dos experimentos, donde se utilizaron frutos rojos de Palma Kerpis, los cuales fueron recolectados en los jardines del Aeropuerto Internacional de Cancún; provenientes de palmas sanas, con altura promedio de 2,5 m y con riego.
Experimento 1. Germinación de semilla de Palma Kerpis
Se eligieron cuatro infrutescencias maduras de las palmas seleccionadas, se separaron los frutos manualmente y mediante el proceso de flotación en agua, se eliminaron los inviables. El fundamento del método se basa en que los frutos vacíos, es decir, los que no han desarrollado convenientemente la semilla (embrión y endospermo) flotan en agua. Aunque hay excepciones, porque algunos frutos de palmas flotan naturalmente debido a que se dispersan por el agua; y porque si estos se llegaran a sembrar, un número regular de ellos germinaría. Los frutos se clasificaron manualmente en tres tamaños, obteniendo sus valores promedio (Tabla I).
]]>
Posteriormente, se realizó la remoción mecánica del exocarpio, mesocarpio y endocarpio de los frutos; después se lavaron en agua con hipoclorito de sodio al 5 % y se dejaron secar a la sombra durante tres días. A continuación, las semillas (compuestas en su mayor parte por endospermo) fueron sembradas (15/noviembre/2010) en charolas germinadoras a 2 cm de profundidad en sustrato peat moss, manteniendo la humedad durante todo el experimento, en un umbráculo con 40 % de sombra; a una temperatura promedio de 27,1 °C y 70 % de humedad relativa.
Al momento de la emergencia, cada plántula fue colocada en una bolsa de vivero color negro de 22 x 22 cm, y separadas en bloques de 100 plántulas por tamaño de fruto (chico, mediano y grande). Mensualmente se registró el crecimiento de las plántulas y hasta los 18 meses, que fue el momento en el cual las raíces agotaron los recursos contenidos en el suelo y se desarrollaron las primeras hojas foliadas; no se aplicó fertilización. Se utilizó como sustrato un suelo del suborden calcimórfico, que de acuerdo con la nomenclatura maya es yaaxhom (somero, pedregoso y poco fértil) y que corresponde al tipo luvisol (Tabla II).
Las variables evaluadas fueron:
Los datos obtenidos en las diferentes variables de respuesta se sometieron a análisis de varianza unidireccionales e independientes para cada experimento, el primero para un diseño completamente al azar y el segundo para un diseño en bloques completos al azar, con un arreglo factorial (2x5); las pruebas de comparación medias se realizaron por el método de Tukey (α≤0,05) usando el software MINITAB 15.
]]>RESULTADOS Y DISCUSIÓN
Germinación de semilla de Palma Kerpis
El polimorfismo o variación en tamaño y forma de la semilla en Palma Kerpis (semillas redondas o diversamente alargadas), obedece a diferencias en la adjudicación de recursos. Está relacionado con el ambiente en que la planta madre se encontró cuando los frutos se desarrollaban, incluye factores como la nutrición edáfica, la posición de la infrutescencia en la palma, la posición del fruto, la remoción de partes de la palma, la época del año, la humedad del suelo, la radiación solar, la longitud del día, la temperatura y la presencia de plagas, principalmente (10, 11).
En el área de estudio, se pueden hallar palmas con hasta ocho infrutescencias (de diferente grado de desarrollo) y frutos maduros prácticamente todo el año. Las primeras infrutescencias en desarrollarse tienen más frutos y de mayor tamaño. En el conteo de las infrutescencias utilizadas, se tuvieron en promedio 680 frutos por infrutescencia, de los cuales el 47 % fueron grandes, el 35 % fueron medianos y el 18 % fueron chicos. Lo que equivale a tener 4,8 kg de material propagativo por cada 1000 frutos. Los valores promedio por tamaño de fruto se pueden visualizar en la Tabla I, aunque debe tenerse en cuenta que se trata de los frutos y no en sí de las semillas.
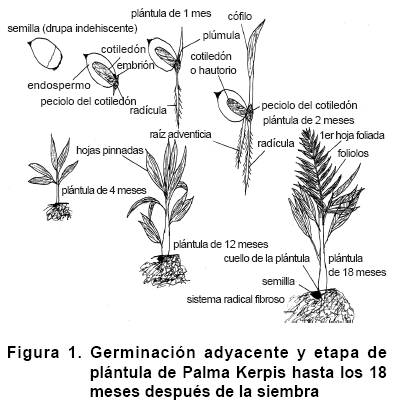
La germinación en Palma Kerpis fue adyacente, sólo el peciolo del cotiledón surge de la semilla. Aparece como un cuerpo hinchado que termina en la superficie de la semilla y se llama “botón”. La radícula y la plúmula surgen del fondo y cima del botón, respectivamente. La radícula es delgada y rápidamente fue reemplazada por raíces adventicias (formadas en la base del tallo); el cotiledón o haustorio permaneció dentro de la semilla absorbiendo los nutrimentos del endospermo (Figura 1). La persistencia de la semilla adherida al tallo de la plántula se mantuvo aproximadamente hasta los 18 MDS.
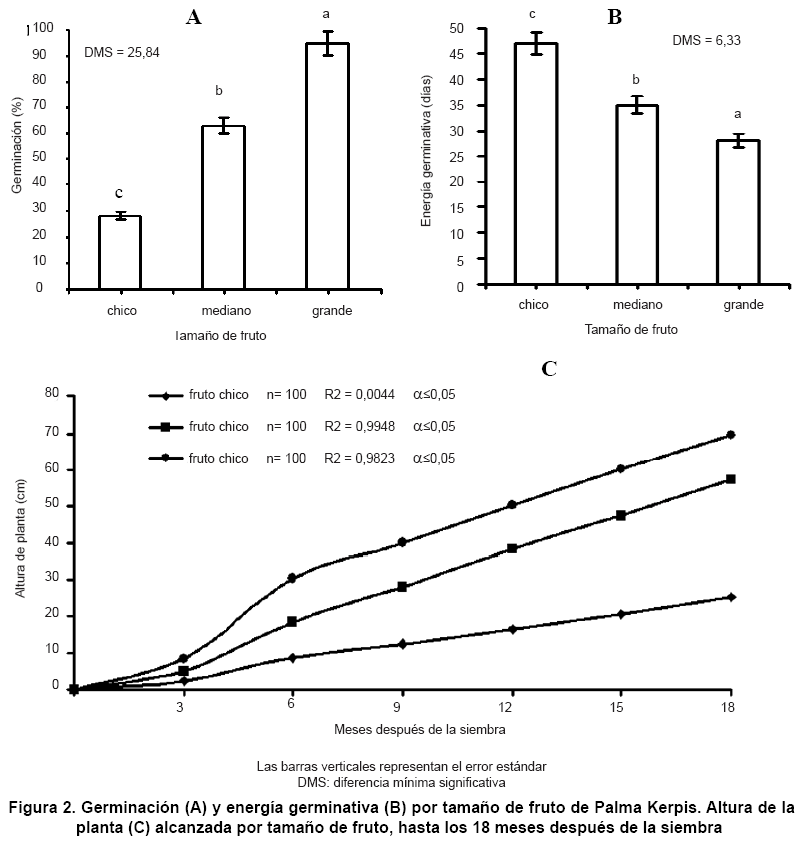
El tamaño del fruto influyó significativamente (α≤0,05) en la germinación de Palma Kerpis. Al utilizar frutos de 2,08 y 3,57 cm en su diámetro ecuatorial y polar, respectivamente (fruto de tamaño grande), se observó que la germinación de la semilla fue del 95 % (Figura 2A) y esta ocurrió a los 28 días después de la siembra (Figura 2B), alcanzando las plántulas una AP de 69,6 cm hasta los 18 MDS (Figura 2C); lo cual coincidió con el desarrollo de sus primeras hojas foliadas.
En la medida que el tamaño de la semilla ocupada para la siembra fue menor, así también lo fue la germinación y la AP, debido a una menor reserva de nutrimentos contenidos en el endospermo; así como al deficiente desarrollo de los embriones en semillas más chicas, lo cual coincide con los estudios hechos en guarumbo (Cecropia obtusifolia) (12) y en palma areca (Dypsis lutescens) (3).
El enfoque es que la Palma Kerpis, una vez superada la etapa de plántula, requiere de ambientes abiertos y es tolerante a periodos cortos de sequía, por ello produce semillas grandes que cuentan con mayor reserva de humedad. El contenido nutrimental del endospermo de la semilla es un factor que afecta la germinación, el alargamiento y el crecimiento de las plántulas. El tamaño de la semilla también está relacionado positivamente con el tamaño inicial de la planta, con el vigor y la supervivencia de plántulas (13). En esta investigación, se apreció un mayor desarrollo inicial en las plántulas provenientes de frutos (semillas) grandes. Esta ventaja de tamaño inicial de la plántula puede persistir hasta la madurez (14) o perderse con el tiempo (15).
Las semillas grandes tienen otras características que les confieren ventaja, como una mayor capacidad para emerger desde mayores profundidades del suelo, a diferencia de las pequeñas (13). Cuando se tienen deficiencias de nutrimentos en el suelo, una plántula proveniente de una semilla grande tiene más probabilidad de sobrevivir que una de semilla pequeña porque cuenta con más reservas (energía germinativa). Aunque cuando las plantas se desarrollan en ambientes limitativos tienden a mostrar mayor variación en el tamaño de las semillas que producen, cuestión observada en jara pringosa (Cistus ladanifer) (10) y en guarumbo (Cecropia obtusifolia) (12).
Los resultados de esta investigación indicaron que el tamaño de la semilla influyó en la energía germinativa (α≤0,05); dado que el polimorfismo de la semilla desempeña un papel importante en los procesos de germinación y establecimiento de plántulas, tal como fue reportado en semillas de encino blanco (Quercus rugosa) (13). En la medida que el tamaño de la semilla utilizada en la siembra fue mayor, así también se obtuvo un mayor porcentaje de germinación; lo cual coincidió con los estudios hechos en semillas de palma de aceite (Elaeis guineensis), donde las más grandes y con mayor masa, iniciaron primero su germinación y presentaron mayor energía germinativa, que en semillas más chicas (16); recomendando así la utilización de semillas grandes en la siembra. ]]>
La prueba de Tukey (α≤0,05) indicó que la germinación de todas las categorías de tamaño fue significativamente diferente (Figura 2); lo cual, en su ambiente natural puede ser una ventaja para la Palma Kerpis, dado que la variación en el tamaño de semilla provee mayores probabilidades en distintos ambientes de germinación. Resultados similares fueron obtenidos en jara pringosa (Cistus ladanifer) (10), en guarumbo (Cecropia obtusifolia) (12) y en palma de aceite (Elaeis guineensis) (16), quienes concluyeron que la germinación se incrementa en relación directa con el tamaño y masa de la semilla, presentando diferencias estadísticamente significativas entre semillas grandes, medianas y chicas.
Los tiempos que describen la germinación de la semilla y la etapa de plántula en Palma Kerpis comprende de 0 a 18 meses (tiempo en el cual agota las reservas nutrimentales del endospermo, logra su completa independencia y está lista para el trasplante al lugar definitivo), lo cual se puede apreciar en la Figura 1. Aunque hace falta más investigación acerca de qué tantos carbohidratos contiene la semilla y a qué velocidad los utiliza, lo cual es de relevancia para tener éxito en distintos ambientes de germinación (10); particularmente en aquellos en los que la plántula puede depender durante más tiempo de sus reservas en lo que logra establecerse (17).
Puede especularse que la remoción del exocarpio, mesocarpio y endocarpio favoreció la germinación, debido a que se disminuyó la concentración de inhibidores del crecimiento presentes en el fruto. Al respecto, por las pequeñas diferencias obtenidas entre frutos, con o sin pericarpio de palma corocito (Syagrus stenopetala), no se justifica esta práctica, siempre y cuando la disponibilidad del material sea suficiente (18). En Palma Kerpis esta puede ser una opción, por lo cual harían falta más estudios al respecto; sin embargo, lo que sí se recomienda, es sembrar las semillas poco después de haberlas recolectado y limpiado. Los resultados de este experimento coinciden con los obtenidos en palma real caribeña (Roystonea oleraceae), al atribuir el éxito de la emergencia, al uso de semillas maduras, lavadas y secadas adecuadamente (19).
Efecto de poda de Palma Kerpis en el establecimiento postrasplante
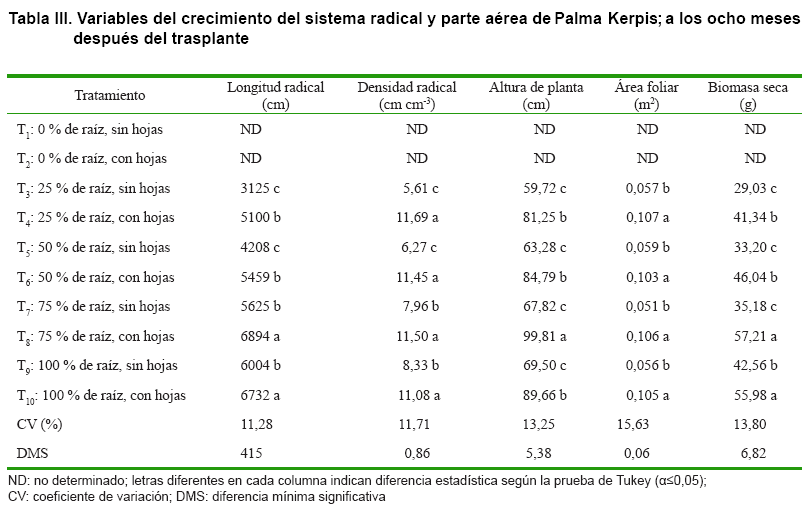
Las palmas a las cuales se les eliminó la raíz al momento del trasplante (T1 y T2) comenzaron a necrosarse y morir tempranamente; dado que esta especie no tuvo la capacidad de generar raíces nuevas, hasta los ocho meses después del trasplante (MDT), cuando previamente se le eliminaron; aun y cuando tuviera hojas en su parte aérea. Por lo cual sus valores en LR, DR, AP, ÁF y BS no se determinaron (Tabla III).
]]>
Los tratamientos sobresalientes en LR fueron T8 y T10 lo cual indica diferencia estadística para esta variable (α≤0,05). En general, los tratamientos donde se dejaron las hojas al momento del trasplante (T4, T6, T8 y T10) el crecimiento en LR fue superior, comparado con los tratamientos sin presencia de hojas al momento del trasplante (T3, T5, T7 y T9); es decir, el sistema radical creció más en LR cuando a las palmas se les dejaron hojas al momento del trasplante, en forma correspondiente a su porcentaje de raíz inicial. Mientras que la igualdad estadística en T4, T6, T7 y T9 pudo deberse a una compensación en la ritmicidad del sistema radical y su parte aérea (20).
De los sistemas radicales que crecieron más, T8 y T10 (con 75 y 100 % de raíz inicial al trasplante y ambos con hojas, respectivamente) ellos mostraron superioridad en otras variables como DR, ÁF y BS, siendo las dos últimas, las más determinantes para la productividad de una planta (5).
En DR T4, T6, T8 y T10 tuvieron valores sobresalientes para esta variable, con un incremento aproximado dos veces mayor respecto de los tratamientos a los cuales se les eliminaron las hojas al momento del trasplante (T3, T5, T7 y T9); mostrando diferencia estadística entre ellos (α≤0,05). Lo cual sugiere que la presencia de hojas (con un ÁF = 0,098 m2) al momento del trasplante, fue fundamental para llevar a cabo la producción de fotoasimilados y para estimular la DR.
En AP el tratamiento sobresaliente fue T8, lo cual indica que una poda ligera de las raíces (al 25 %) al momento del trasplante favoreció el crecimiento en AP; y que si se eliminan las hojas al momento del trasplante, la AP puede disminuir hasta un 40 %, retrasando con ello el establecimiento postrasplante. En ÁF los tratamientos sobresalientes fueron T4, T6, T8 y T10. Aquí, la respuesta de desarrollo de las hojas se vio limitada cuando estas fueron suprimidas al momento del trasplante; sin embargo, en los tratamientos en los cuales las hojas se dejaron, el ÁF no mostró diferencia estadística (T4, T6, T8 y T10).
En BS los tratamientos sobresalientes fueron T8 y T10 (con 75 y 100 % de raíz inicial al trasplante y ambos con hojas, respectivamente); en esta variable, la ganancia de masa seca (a los ocho MDT) fue casi el doble respecto del T3 (25 % de raíz inicial al trasplante y sin hojas); lo que sugiere que el efecto de poda (radical y aérea, al momento del trasplante) influye significativamente en la productividad (BS y ÁF) de la Palma Kerpis. ]]>
La finalidad de utilizar, en la propagación de Palma Kerpis, frutos grandes, es obtener plantas de mayor talla respecto del promedio en el mismo periodo de siembra. Si a las plántulas al momento del trasplante, se les realiza una poda ligera dejando un 75 % de la raíz inicial y manteniendo las hojas fotosintéticamente activas, se acelera el crecimiento de las mismas para de esta manera lograr plantas con una mayor calidad y en un menor tiempo. Al mismo tiempo, se recomienda que al momento del trasplante no se desuna la conexión de la semilla a la plántula, porque esta, aun se sigue abasteciendo de los nutrimentos contenidos en el endospermo.
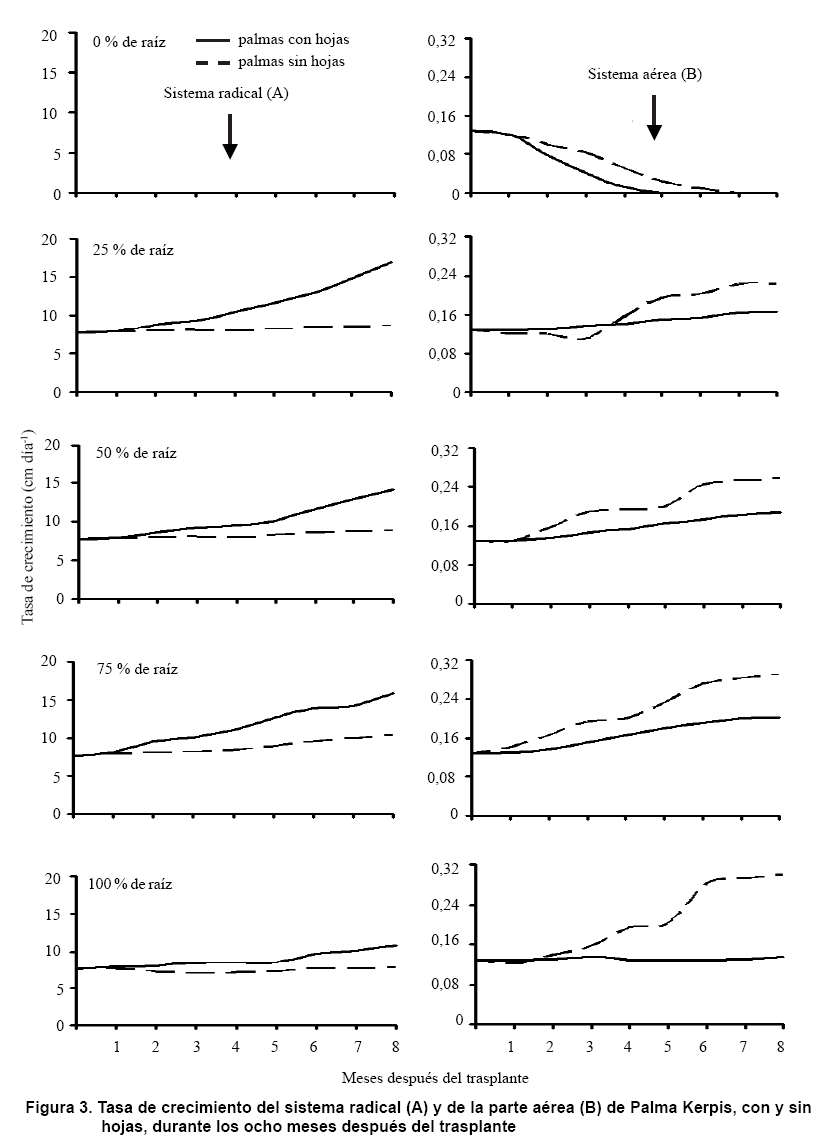
En la Figura 3, se observa que en el 1er MDT el sistema radical y la parte aérea de Palma Kerpis, no tuvieron crecimiento debido a la adaptación al nuevo medio (estrés postrasplante). Posteriormente, se inició el crecimiento de ambos y la ramificación del sistema radical. En todos los tratamientos (excepto en T1 y T2, quienes no tuvieron la capacidad de generar raíces cuando al momento del trasplante le fueron eliminadas), los sistemas radicales de Palma Kerpis mostraron el siguiente patrón de crecimiento; en palmas con hojas al momento del trasplante: de 0 al 4to MDT tuvieron un crecimiento lento (de 7,66 a 11,11 cm día-1), a partir del 4to mes el crecimiento fue acelerado, alcanzando una TC de 16,94 cm día-1 hacia los ocho MDT. Mientras que en palmas sin hojas al momento del trasplante, el crecimiento hasta el 8vo MDT siempre fue lento (de 7,77 a 10,50 cm día-1) (Figura 3A). Lo que indica que la raíz de Palma Kerpis creció 9.20 cm·día-1 más, cuando hubo hojas en su parte aérea, que cuando no las hubo; y que las podas de raíz (al 25, 50 y 75 %) al momento del trasplante, estimularon aún más el crecimiento de la misma.
El crecimiento de la parte aérea en Palma Kerpis, mostró el siguiente patrón (excepto en T1 y T2, quienes por la falta de raíces comenzaron a morir a los cinco MDT); las palmas a las cuales se les dejaron las hojas al momento del trasplante, tuvieron un crecimiento lento (0,13 a 0,20 cm día-1), hasta los ocho MDT (Figura 3B). Mientras que las palmas a las cuales se les eliminaron las hojas al momento del trasplante, de 0 al 3er MDT tuvieron un crecimiento lento, a partir del 3er y hasta el 8vo MDT su crecimiento fue acelerado con ligeras estacionalidades; alcanzando una TC de 0,29 cm día-1 (Figura 3B). Lo que indica que el crecimiento de la parte aérea en Palma Kerpis fue mayor cuando al momento del trasplante se eliminaron las hojas y se mantuvo del 75 al 100 % de la raíz inicial.
Solo en la parte aérea se pudo apreciar una TC sigmoidal, dado que el tiempo de observación (ocho meses) no fue suficiente para observar un ciclo de alternancia completo para esta especie; sin embargo, los resultados coinciden con lo obtenido en otros cultivos como: roble (Quercus pubescens) (20), chaya (Cnidoscolus chayamansa) (21) y guanábano Annona muricata (22). Esta TC sigmoidal comienza en un origen, avanza lentamente, aumenta en forma exponencial (en relación proporcional con el tamaño del organismo), se mantiene en reposo y nuevamente aumenta.
En esta etapa, se observó un crecimiento sincrónico, tanto del sistema radical como de la parte aérea; por lo tanto, los fotosintatos se estuvieron traslocando tanto al meristemo apical de la raíz donde hubo división celular y alargamiento, como al meristemo apical del tallo. Sin embargo, es muy probable que en esta especie se presente una alternancia entre la raíz y la parte aérea debido a que se trata de sistemas complementarios, en donde primero crece un sistema y después el otro; así, la estrecha relación que guardan ambos componentes estructurales parece ser una generalidad sin importar la especie o el ambiente en el que se desarrollen (23).
CONCLUSIONES
Para obtener plántulas de Palma Kerpis de calidad con buen crecimiento postrasplante se recomienda utilizar semilla proveniente de frutos grandes y maduros (color rojo). Las semillas germinan hasta un 95 % a los 28 días en promedio, aunque el porcentaje de germinación disminuye en relación directa con el tamaño del fruto. Mientras que, el trasplante al lugar definitivo se debe realizar cuando las palmas tengan 18 meses o el desarrollo de su primera hoja foliada; y al momento del trasplante se debe realizar una poda dejando el 75 % de la raíz inicial y las hojas verdes fotosintéticamente activas.
REFERENCIAS
1. Lim, T. K. Edible Medicinal and Non-Medicinal Plants: Volume 1, Fruits. Springer Netherlands. 2012, pp. 257-259.
2. Evert, R. F.; Esau, K. y Eichhorn, S. E. Anatomía vegetal: meristemas, células y tejidos de las plantas: su estructura, función y desarrollo. Ed. Omega, v.1, 3ra.ed. Barcelona, España, 2008, pp. 614.
3. Benítez, B. y Soto, F. El cultivo de la palma areca (Dypsis lutescens, H. Wendel). Cultivos Tropicales, 2010, vol. 31, no. 1, pp. 62-69. ISSN 1819-4087.
4. Hunt, R.; Causton, D. R.; Shipley, B. y Askew, A. P. A modern tool for classical plant growth analysis. Annals of Botany, 2002, vol. 90, no. 4, pp. 485-488. ISSN 1095-8290.
5. López-Ríos, G. F. Ecofisiología de árboles. 2da. Edición. Universidad Autónoma Chapingo, Texcoco, México, 2009, pp. 490.
6. NOM. Norma Oficial Mexicana NOM-021-RECNAT-2000, que establece las especificaciones de fertilidad, salinidad y clasificación de suelos, estudios, muestreo y análisis. México, 2000.
7. Kaspar, T. C. y Ewing, R. P. Rootedge: Software for measuring root length from desktop scanner images. Agronomy Journal, 1997, vol. 89, pp. 932-940. ISSN 1435-0645.
8. Casanova, F.; Escamilla, J. A. y Ramírez, L. Modelo teórico para la estimación de la densidad radicular de dos especies leñosas. Tropical and Subtropical Agroecosystems, 2009, vol. 10, pp. 263-271. ISSN 1870-0462.
9. Newman, E. I. A method to estimating the total length of root in a sample. Journal of Applied Ecology, 1966, vol. 3, no. 1, pp. 139-145. ISSN 1365-2664.
10. Delgado, J. A.; Serrano, J. M.; López, F. y Acosta, F. J. Seed size and seed germination in the Mediterranean fire-prone shrub Cistus ladanifer. Plant Ecology, 2008, vol. 197, no. 2, pp. 269-276. ISSN 1573-5052.
11. Ding, X. D.; Zhang, S. R.; Li, Y.; Tian, C. Y. y Feng, G. Seed polymorphism of an euhalophyte, Suaeda acuminata and its adaptation strategy to extremely saline soil. Acta Botanica Boreali-Occidentalia Sinica, 2010, vol. 30, pp. 2293-2299. ISSN 1000-4025.
12. Tenorio-Galindo, G.; Rodríguez-Trejo, D. A. y López-Ríos, G. Efecto del tamaño y color de la semilla en la germinación de Cecropia obtusifolia Bertol (Cecropiaceae). Agrociencia, 2008, vol. 42, pp. 585-593. ISSN 1405-3195.
13. Huerta-Paniagua, R. y Rodríguez-Trejo, D. A. Efecto del tamaño de semilla y la temperatura en la germinación de Quercus rugosa Née. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 2011, vol. 17, no. 2, pp. 179-187. ISSN 2007-4018.
14. Weis, M. I. The effects of propagule size on germination and seedling growth in Mirabilis hirsuta. Canadian Journal of Botany, 1982, vol. 60, pp. 1868-1874. ISSN 1480-3305.
15. Zimmerman, J. K. y Weis, M. I. Fruit size variation and its effects on germination and seedling growth in Xanthium strumarium. Canadian Journal of Botany, 1983, vol. 61, pp. 2309-2315. ISSN 1480-3305.
16. Myint, T.; Chanprasert, W. y Srikul, S. Effect of seed weight on germination potential of different oil palm (Elaeis guineensis Jacq.) crosses. Seed Science and Technology, 2010, vol. 38, no. 1, pp. 125-135. ISSN 1819-5717.
17. Bonfil, S. C. The effects of seed size, cotyledon reserves and herbivory on seedling survival and growth in Quercus rugosa and Q. laurina (Fagaceae). American Journal of Botany, 1998, vol. 85, pp. 79-87. ISSN 1537-2197.
18. Maciel, N. y Briceño, A. Efecto de la madurez de frutos, escarificación de la semilla y temperatura en la emergencia de Syagrus stenopetala Burret. Revista de la Facultad de Agronomía de la Universidad del Zulia, 2009, vol. 26, pp. 196-211. ISSN 1690-9763.
19. Maciel, N. Emergencia de la palma real venezolana (Roystonea oleraceae) en función de condiciones variables del fruto y la semilla. Bioagro, 2001, vol. 13, no. 3, pp. 105-110. ISSN 2074-8647.
20. Willaume, M. y Pagès, L. Correlated responses of root growth and sugar concentrations to various defoliation treatments and rhythmic shoot growth in oak tree seedlings (Quercus pubescens). Annals of Botany, 2011, vol. 107, pp. 653-662. ISSN 1095-8290.
21. Aguilar-Luna, J. M. E.; García-Villanueva, E.; Macario-Mendoza, P. A.; Huerta-Lwanga, E. y de Alba-Becerra, R. Effect of the plantation density in the growth and productivity of Cnidoscolus chayamansa McVaugh (Euphorbiaceae). Tropical and Subtropical Agroecosystems, 2012, vol. 15, pp. 77-86. ISSN 1870-0462.
22. Aguilar-Luna, J. M. E. y García-Villanueva, E. Crecimiento postgerminativo y concentración nutrimental en hojas de Annona muricata L. cultivadas en suelos calcimórficos. Revista Brasileira de Ciências Agrárias, 2013, vol. 8, no. 1, pp. 62-70. ISSN 1981-0997.
23. Fourcaud, T.; Zhang, X.; Stokes, A.; Lambers, H. y Körner, C. Plant growth modelling and applications: the increasing importance of plant architecture in growth models. Annals of Botany, 2008, vol. 101, no. 8, pp. 1053-1063. ISSN 1095-8290.
Recibido: 9 de mayo de 2013
Aceptado: 3 de septiembre de 2013
Dr. Jesús Mao Estanislao Aguilar Luna, Profesor-Investigador del Programa de Ingeniería Agroforestal, Facultad de Ingeniería Agrohidráulica, Benemérita Universidad Autónoma de Puebla, Puebla, México. Email: maoaguilar@colpos.mx
]]>{kind=link}
{kind=link}
{kind=link}