
Patrón de la actividad de las β -1,3-glucanasasy quitinasas en la interacción hma-sistemina en tomate. II fase temprana de la simbiosis
activitys pattern of β-1,3-glucanases and quitinases in the AMF - systemin interaction in tomato. II Early symbiotic phase
Dra.C. Blanca M. de la Noval-Pons,I Dra.C. Ondina León-Díaz,II M.Cs. Norma A. Martínez-Gallardo,III Dr.C. Eduardo Pérez-Ortega,I Dr.C. John P. Délano-FrierIII
IInstituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
IICentro Nacional de Sanidad Agropecuaria (CENSA). Apartado 10. San José de las Lajas, Mayabeque, Cuba, CP 32700.
IIIUnidad de Biotecnología e Ingeniería Genética de Plantas del Cinvestav-Campus Irapuato. Km 9,6. Libramiento Norte, Carr. Irapuato-León. A.P. 629, C.P. 36821, Irapuato, Gto. México.
]]>
RESUMEN
La sistemina (s) y el acido jasmónico (AJ) juegan un papel central en la respuesta al daño en tomate (Solanum lycopersicum). El AJ es también conocido como un regulador de la simbiosis micorrízica arbuscular (MA) en esta y en otras especies de plantas. Se realizó un experimento en el cual se evaluó la posible participación de la sistemina en etapas tempranas del establecimiento de la micorrización. La aplicación exógena de la sistemina produjo un rápido y transciente incremento de los patrones de actividad de las glucanasas y quitinasas a los 3, 6 y 12 días post emergencia (dpe). Las especies de los hongos micorrízicos arbusculares (HMA) produjeron un incremento gradual con picos transcientes de actividad, a los 3 y 6 dpe (para quitinasa) y 9 dpe (para β-1,3-glucanasa). El patrón sugiere el establecimiento de un diálogo pre-simbiótico rápido planta-HMA que parece ser modulado en parte por la sistemina, avalado por el efecto sinérgico sobre la actividad de β-1,3-glucanasa en el tratamiento de la sistemina y HMA. Se observó un efecto positivo similar sobre la actividad de glucanasa después de la aplicación de la sistemina en las etapas más tempranas de la colonización. Esto sugiere que la sistemina podría tener un impacto sobre el proceso de micorrización del tomate con la modulación de las respuestas de defensa.
Palabras clave: enzimas pectinolíticas, mycorrhizae, respuesta de defensa, sistemina.
ABSTRACT
Systemin (s) and jasmonic acid (JA) play a central role in the damage response in tomato (Solanum lycopersicum). JA is also known to regulate the arbuscular mycorrhizal (AM) symbiosis in this and other plant species. An experiment was made in which the possible participation of the systemin in early stages of the establishment of the mycorrhiza was evaluated. Exogenous systemin appliction to a very rapid and transient increase of root glucanase and quitinase activity patterns at 3, 6 and 12 days after emergence (dae). Arbuscular mycorrhizal fungi (AMF) species produced a gradual increment leading to transient peaks of activity, at 3 and 6 dae (for chitinase) and 9 dae (for β-1,3-glucanase). The pattern suggests the establishment of a pre-symbiotic dialogue plant - AMF which appeared to be partly modulated by systemin, judging by the synergic effect on β-1,3-glucanase activity observed in the systemin and AMF treatment. A similarly positive effect on β-1,3-glucanase activity was observed after systemin application at the earliest colonization stages. The above suggests that systemin could impact the tomato mycorrhizal process with the modulation of defense responses.
Key words: pectinolytic enzymes, mycorrhizae, defense mechanisms, sistemine.
INTRODUCCIÓN
Bajo condiciones naturales, las raíces de más del 80 % de las plantas están asociadas con hongos simbióticos del suelo, donde las Micorrizas Arbusculares (MA) constituyen uno de los grupos principales (1). La colonización radical por HMA induce importantes cambios en la expresión de diferentes genes relacionados con las respuestas de defensa en la planta hospedera, que le permiten superar los estreses bióticos y abióticos (2).
En este proceso se produce la inducción de respuesta de resistencia sistémica, similar a la producida ante ciertos hongos y bacterias, con la inducción de proteínas diversas relacionadas con la patogenicidad (PRs), entre las que se reportan las β-1,3-glucanasas, quitinasa, y enzimas relacionadas con los procesos oxidativos (3,4).
La inducción de estos mecanismos en etapas tempranas del establecimiento de la asociación micorrízica ha recibido especial atención debido a su posible implicación en la regulación de la simbiosis, así como en la protección de las plantas contra el ataque de patógenos. Estas respuestas de defensa ocurren de forma transciente, similar a como se observa en las interacciones planta-patógeno (5).
Otros autores estudiando la cascada de señales inducida durante el ataque por insectos herbívoros o por daño mecánico en hojas de tomate (6), encontraron la inducción de una hormona polipeptídica de 18 amino ácidos, la sistemina, involucrada en la expresión de genes de defensa en plantas de tomate y otras Solanáceas. Una vez reconocida por las células diana, la sistemina produce la activación de una cascada de señales que involucran a la vía de octadecanoicos produciendo la acumulación transciente de ácido jasmónico (AJ) en hojas de tomate y la expresión de genes relacionados con la respuesta sistémica inducida (7).
Se ha informado que esta hormona es requerida para la inducción de la respuesta sistémica de la llamadas proteínas de respuesta sistémicas al daño (8), la cual incluye varias proteínas asociadas a las vías de señalización y otras similares a las inducidas por insectos y herbívoros (9).
Estudios recientes dirigidos a analizar el posible papel de la sistemina en la modulación local o sistémica de la asociación micorrízica indican que la aplicación en plantas de tomate micorrizadas induce la acumulación de β-1,3-glucanasas y quitinasas (10). En este trabajo se muestran cambios en los patrones de inducción de la respuesta sistemina inducida (ISR, por su término en inglés) en plantas micorrizadas por mecanismos no identificados, en estadio tardío.
Con el objetivo de poder dilucidar el posible papel de la sistemina en el dialogo entre el HMA y la planta, con la participación de mecanismos de señalización comunes, se evaluó el patrón de la actividad de proteínas PRs inducidas por esta interacción.
MATERIALES Y MÉTODOS
Obtención del material vegetal y aplicación de los productos
Se emplearon plantas de tomate (Solanum lycopersicum L.) del cultivar “Amalia” (11). Las semillas fueron desinfectadas superficialmente con hipoclorito de sodio comercial al 10 % durante 10 min, seguido de tres lavados con agua destilada (H2O d) estéril, posteriores a los cuales se procedió a su siembra. El sustrato estuvo conformado por una mezcla de suelo Ferralítico Rojo Lixiviado típico y humus de lombriz en relación 1:1 (v:v), estéril (Tabla I). Las plantas se desarrollaron en cámara de crecimiento, bajo condiciones controladas, con fotoperíodo de 8 h de oscuridad a 16 °C y 16 h de luz a 28 °C.
Se seleccionó la especie de HMA Funneliformis mosseae (T.H. Nicolson & Gerd.) (12) (Fm), procedente del cepario del laboratorio de micorrizas del Instituto Nacional de Ciencias Agrícolas (INCA) (Registro de patente No.2264), la cual se conservó en un sustrato desarrollado para estos fines a 4 ºC. El inóculo empleado poseía un título promedio de 50 esporas g-1 de sustrato, certificado por dicho laboratorio.
Se emplearon 100 semillas del cultivar Amalia, fueron sembradas directamente sobre macetas de 1 kg. La inoculación con HMA se realizó mediante el recubrimiento de las semillas (14). Posteriormente, en el momento de la emergencia se aplicó la sistemina de tomate, para lo cual se empleó un producto obtenido de forma sintético (Laboratorios BQ SOS, de México), fue adicionado al sustrato en solución (1 mL por planta a 44,06 nmol). Se mantuvo un tratamiento control que no poseía HMA ni sistemina, en el cual se adicionó agua destilada a razón de 2 mL por planta.
Para los muestreos se siguió una dinámica desde los tres hasta los 24 días posteriores a la emergencia (dpe), correspondientes a los 3, 6, 9, 15, 18, 21 y 24 dpe.
En los momentos de muestreos las plantas fueron lavadas y separadas por órganos, dividiéndose las raíces en dos grupos. Una porción se empleó para la determinación de las actividades enzimáticas, la cual fue congelada en nitrógeno líquido y conservada a -70 ˚C, mientras que la otra se secó en estufa a 70 ˚C para la determinación del establecimiento de la simbiosis micorrízica (colonización micorrízica).
Colonización micorrízica
Para cuantificar la colonización micorrízica se tomaron las raíces secas a 70 ˚C, se digirieron con KOH, para luego teñirse con azul de tripano. Se evaluó el porcentaje de colonización micorrízica (% C) según el método de los interceptos y la intensidad de la colonización (% I) (10).
Ensayo enzimático
La extracción de proteínas totales y las determinaciones enzimáticas se realizaron según la metodología descrita por Noval y colaboradores (10). A los extractos se les determinó la concentración de proteínas totales mediante el método de Bradford (15), empleando el kit comercial (Bio-Rad Laboratories, USA). Se realizó la determinación de las actividades enzimáticas in vitro en las raíces y en las hojas de las plantas de tomate. Para la cuantificación de la actividad β-1,3-glucanasa (GLN; EC- 3.2.1.39) se usó laminarina como sustrato y se leyó la absorbancia a 450 nm en un lector de microplacas SUMA PR 521. Para la actividad quitinasa (QUI; EC- 3.2.1.14) se utilizó como sustrato la quitina coloidal preparada a partir de quitina grado reactivo (Fluka) y la absorbancia se leyó a 585 nm en el espectrofotómetro (10). Todas las actividades enzimáticas fueron transformadas a μKat (microkatales) y pKat (picokatales) por miligramos de proteínas, según fue necesario (16). Las lecturas de la absorbancia en los diferentes métodos empleados se realizaron en un espectrofotómetro, Ultrospec Plus Spectrophotometer, Pharmacia LKB. Todos los sustratos fueron adquiridos de Sigma-Aldrich Chemical (St. Louis, MO, USA).
Diseño experimental y análisis estadístico
El experimento se desarrolló siguiendo un diseño completamente aleatorizado con tres repeticiones, conformadas cada una por diez plantas, en cada momento de muestreo. Los datos fueron procesados estadísticamente según el análisis de varianza de clasificación simple, comparándose las medias mediante los intervalos de confianza con un grado de significación de 95 %. El experimento se repitió tres veces en el tiempo, de los cuales se seleccionó uno representativo. Todas las comparaciones se realizaron según el paquete estadístico STATGRAPHIC Plus Versión 5.1 (17). Los porcentajes de incrementos se calcularon comparando los tratamientos con el control correspondiente.
RESULTADOS
Dinámica del establecimiento de la simbiosis
Desde los primeros momentos de evaluación (3 dpe) se detectó la presencia de estructuras fúngicas en las raíces, evaluadas como porcentaje de colonización (% C) y de intensidad (% I) (Tabla II), con niveles bajos en los primeros nueve días (6,1 %C y 0,0824 % I). A partir de los 12 y 15 dpe se produjo un incremento, con valores que se mantuvieron hasta el final del experimento (46,56 % C y 1,8 % I), con similitud estadística entre ellos (15 a 24 dpe). Durante esta fase se alcanzaron niveles elevados de ambos indicadores para este cultivo, si se tienen en cuenta los resultados informados en trabajos previos, en los cuales se encontraron valores de 13 porcentaje de colonización y 0,61 porcentaje de intensidad evaluada como densidad visual en plantas de tomate micorrizadas con Funneliformis mosseae (Glomus mosseae sensu lato) a los 32 días posteriores a la germinación (18).
]]>
Se observó que la sistemina actuó como potenciador de la micorrización, a partir de los 12 dpe, al evaluar tanto el porcentaje de colonización, como de intensidad, con porcentajes de 6,98–19,44 y 26,96–228,97, respectivamente, al compararlos con los niveles alcanzados por las plantas donde se inoculó F. mosseae sin la aplicación del elicitor (Fm) durante los 12-24 dpe.
Este periodo coincide con el momento de evaluación. En las plantas micorrizadas (Fm) se produce un incremento importante de ambos indicadores del establecimiento y funcionamiento de la simbiosis, con el consecuente incremento de los procesos metabólicos relacionados con la misma. Posterior al reconocimiento de la sistemina se activa una serie de procesos de señalización, los cuales conllevan a la acumulación de JA (7) y pueden jugar un papel crucial en los procesos de establecimiento y mantenimiento de la simbiosis MA de forma funcional (19-21).
Las raíces micorrizadas exhiben un incremento de los niveles de esta hormona acompañada por la expresión de genes que son inducidos por este ácido y de genes que codifican para su propia biosíntesis; sin embargo, se han informado resultados contrastantes con su aplicación, donde se ha observado, tanto disminución de la tasa de micorrización, como mejora de la interacción HMA-planta (22).
Los jasmonatos pertenecen a una clase diversa del metabolismo lipídico, entre los que se encuentran las oxylipinas. Estudios recientes han revelado la regulación de estos compuestos durante la simbiosis micorrízica arbuscular, donde se produce la modificación del patrón de expresión de genes relacionados con las vías de las oxilipinas (23). Estos informes podrían explicar los resultados observados con la combinación del HMA y el elicitor, si se tiene en cuenta, que la sistemina es un inductor de respuestas de defensa vinculadas con la acumulación de ácido jasmónico y de señales secundarias como las oxilipinas (6).
]]>
Patrón de actividad de enzimas relacionadas con defensa: glucanasa y quitinasa
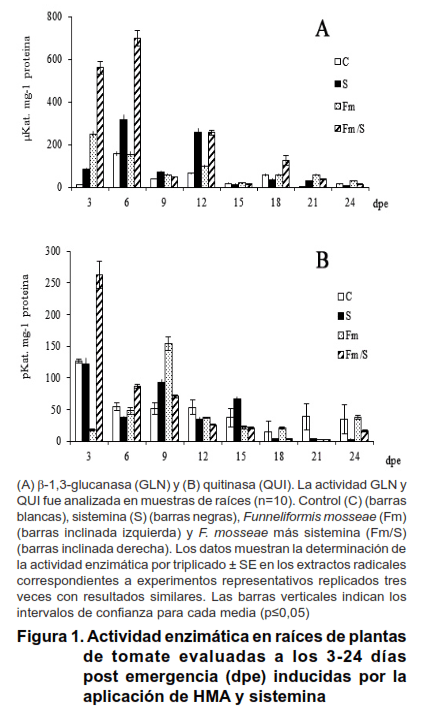
Los patrones de actividad de las enzimas GLN en las raíces de las plantas de tomate solamente micorrizadas (Fm), mostraron una tendencia a alcanzar los niveles más elevados en los primeros estadios del establecimiento de la colonización micorrízica, a los 3 y 6 dpe (Figura 1A), con valores de 248,64
y 154,52 μKat mg-1 proteína, respectivamente. Con relación a la actividad QUI, en las plantas micorrizadas se produjo un incremento de forma más tardía, alcanzando un máximo de actividad a los nueve dpe con 154,05 pKat mg-1 proteína (Figura 1B), respuesta que coincidió con el desarrollo de las primeras fases del proceso de establecimiento de la simbiosis, dada por la presencia del hongo en el interior de la raíz en niveles bajos (6,1-11,06 %C y 0,08-0,30 % I). Posteriormente, la actividad fue decreciendo gradualmente a medida que el proceso de micorrización fue avanzando, hasta alcanzar valores similares o inferiores al control.
Se encontró que la aplicación exógena de la sistemina, de forma independiente, produjo incrementos de la actividad de las GLN en los estadios tempranos, alcanzando los mayores niveles a los 3 y 6 dpe (83,47 y 317,64 µKat mg-1 proteína, respectivamente), con un segundo momento de incremento a los 12 dpe (256,67 µKat mg-1 proteína), los que mostraron diferencias significativas entre sí, con relación a los intervalos de confianza observados. El efecto sobre la actividad de las QUI fue más tardío, observándose niveles que superaron al control a los 9 y 15 dpe (92,13 y 67,05 pKat mg-1 proteína, respectivamente).
Al analizar los resultados en los tratamientos de las combinaciones se encontró un efecto sinérgico entre F. mosseae y la sistemina (Fm/S). Este fue marcado sobre la actividad de las GLU con una inducción rápida y transciente a los 3 y 6 dpe (560,49 y 699,07 µKat mg-1 proteína), los que mostraron diferencias estadísticamente significativas con los tratamientos donde se aplicaron ambos de forma independiente e incrementos de 125,42 % y 352,42 %, respectivamente.
]]>
Con relación a la actividad QUI en raíz (Figura 1B), también se observó una inducción rápida y transciente a los 3 dpe con un efecto sinérgico del HMA y la sistemina (Fm/S) (263,1 pKat mg-1 proteína), que correspondió a un 87,07 % de incremento. Solo a los 9 dpe se produjo una inducción de la actividad de esta enzima en todos los tratamientos evaluados, con relación al control (36,75-199,06 % de incremento); tiempo en el cual las plantas inoculadas con Fm mostraron los valores más elevados (154,05 pKat mg-1 proteína). En estadios tardíos tanto Fm como la sistemina produjeron la represión de la actividad.
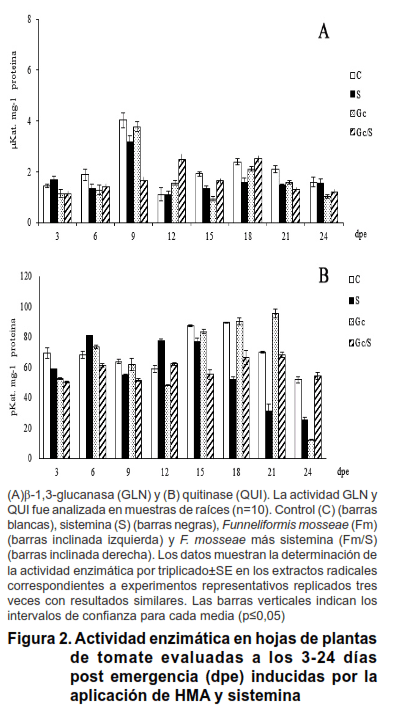
Al analizar la cuantificación de la actividad de las GLN en hojas se observó que los tratamientos mostraron diferencias estadísticamente significativas entre ellos; aunque de forma general, en todos se alcanzaron niveles muy bajos, con relación a los obtenidos en raíz, los que oscilaron entre 1,64 y 2,06 µKat mg-1 proteína (Figura 2A). Solo a los 9 dpe se observó un incremento de la actividad en las plantas control, el que fue seguido en orden decreciente por F. mosseae y por la sistemina (4,02; 3,77 y 3,16 µKat mg-1 proteína, respectivamente).
A los 12 dpe se encontró un efecto sinérgico (2,47 µKat mg-1 proteína) con 90 % de incremento con relación a los bioproductos aplicados, de forma independiente, superó a todos los tratamientos en estudio. Este efecto se mantuvo a los 15 y 18 dpe, los cuales a pesar de mostrar diferencias estadísticas con relación a los tratamientos donde se aplicó F. mosseae y el inductor por separado, no superaron a los niveles basales (control).
Los patrones de actividad de las enzimas QUI en las hojas de las plantas de tomate micorrizadas de forma independiente (Fm) (Figura 2B), mostraron niveles similares con valores cercanos o inferiores al control, con una media de 71,49 pkat mg-1 proteína, a excepción de los 18 dpe, momento donde alcanzó un máximo de 103,08 pKat mg-1 proteína. En las plantas que fueron micorrizadas con F. mosseae, sin la aplicación del elicitor (Fm), se produjo el incremento de la actividad de esta enzima a los 21 dpe
(95,27pKat mg-1 proteína). De forma general, en las hojas se observaron niveles de actividad inferiores a los obtenidos en las raíces, para ambas enzimas.
]]>
Las GLN son abundantes en las plantas y tienen una participación crucial en diferentes procesos fisiológicos. Desempeñan un papel dominante en la división celular, en el intercambio de compuestos en el plasmodesmo y en la reducción de los estreses abióticos (4). Con relación a la función de enzimas en la simbiosis MA se ha encontrado en la pared de los miembros de la familia de las Glomaceae y Acaulosporaceae, la presencia del sustrato de estas hidrolasas, los β-1,3-glucanos, lo que sugiere la posibilidad de que puedan estar involucradas en la degradación parcial de la pared celular de los HMA y de las plantas, facilitando el proceso de penetración en la célula radical (18).
Se observó que la aplicación de la sistemina produjo un incremento transitorio de la actividad de esta enzima en raíz a los 6 dpe. Se ha identificado que esta hormona polipeptídica produce la acumulación de inhibidores de proteinasas, y por lo menos de 19 proteínas más; las que abarcan componentes de las vías de transducción de señales y otras proteínas cuya función en la defensa de las plantas no han sido completamente identificadas. Sin embargo, recientemente estudios peptidómicos revelaron su posible vinculación con la expresión de proteínas PRs. En un análisis cuantitativo de 14 péptidos relacionados con el daño en tomate se obtuvo un nuevo péptido, (CAPE1). El mismo mostró una respuesta similar a la observada por la sistemina, con la inducción de la expresión de genes relacionados con la actividad contra patógenos y herbívoros como los inhibidores de proteasa (PI-1 y PI-2) y de los genes de proteínas relacionadas con la patogénesis PR-1b, β-1,3-glucanasa (PR-2) y las quitinasas, entre otras. De forma similar a la sistemina el péptido CAPE1 activa la biosíntesis de AJ (24).
Como se conoce las QUI participan en diferentes procesos fisiológicos, con funciones diversas, que pudieron haber producido el incremento puntual de isoformas de la enzima no relacionadas con la micorrización, como se observó en el tratamiento control. Estas enzimas son abundantes en las raíces donde pueden actuar liberando señales moleculares que actúan como elicitores endógenos de la morfogénesis. Se ha observado que podrían participar en el proceso de desarrollo de las raíces, al inducir la diferenciación de callos de tomate en cultivo in vitro y en la embriogénesis en zanahoria (25).
Con relación a la sistemina se desconoce cuál pueda ser la causa de la represión de la actividad de las QUI en raíz a los 12 dpe, aplicada de forma independiente, al ser comparada con las plantas control. Por otra parte, tampoco se ha encontrado una influencia negativa de los componentes de la vía de señalización de este elicitor con la inducción de QUI.
Durante los estadios iniciales del establecimiento de las MA, predominan las QUI no relacionadas con la simbiosis, las que producen respuestas de defensa atenuadas; mientras que en las fases más avanzadas ocurre la acumulación de quitinasas específicas de la micorrización (26). En un estudio de la micorrización de plantas de tomate de la variedad “Amalia” inoculadas con R. clarum se obtuvo un incremento de la actividad de esta enzima vinculadas con la micorrización a los 32 días. Al parecer en el presente experimento este momento de inducción se produjo a los 9 dpe en raíz y a los 21 dpe en hojas, lo cual confirma la ocurrencia de un proceso de señalización rápida que condujo a la inducción de respuestas de defensa sistémicas (18).
]]>
Se ha sugerido la posible participación del AJ en la señalización requerida para el establecimiento de la respuesta sistémicas inducida (ISR) en las plantas micorrizadas, como parte de la resistencia inducida por micorrizas, reconocida por MIR (por su término en inglés) (27). Este produce un efecto priming, dependiente del AJ (28), esta hormona es crucial en la amplificación de la respuesta ante el daño mecánico en tomate (8,7,29). Diversos autores refieren la inducción de la ISR en las plantas micorrizadas (30); sin embargo, la contribución del AJ al MIR no está totalmente clara, así como el control de esta respuesta a larga distancia (27).
El uso de los biofertilizantes y de los elicitores en la práctica agrícola ha tomado un gran auge en la última década, los cuales han sido empleados con objetivos diversos. Se observó un efecto sinérgico del HMA y de la sistemina, sobre la inducción de proteínas relacionadas con defensa en estadios tempranos, de forma local, lo que sugiere la posibilidad de la convergencia de los mecanismos de acción por los cuales ambos inducen la ISR.
El hecho de que la sistemina estimule los procesos de establecimiento de la simbiosis micorrízica, evaluados como porcentaje de colonización e intensidad de la misma, avala el papel relevante del AJ en los procesos de establecimiento y mantenimiento de la simbiosis MA, lo cual ha sido sugerido por diferentes autores. Los resultados evidencian la existencia de un receptor para este elicitor en las raíces, de forma similar ha sido identificado en hojas.
CONCLUSIONES
La sistemina potenció el establecimiento de la simbiosis micorrícica con F. mosseae, así como su acción sobre la inducción de GLN y QUI, al producir un incremento rápido y transciente en raíz, en los primeros estadios del establecimiento de la micorrización. Se conoce que este elicitor produce la activación rápida de respuestas de defensa en las cuales participan diferentes elementos que conllevan a la acumulación de AJ y etileno, que son inductores de la expresión de proteínas PRs. Sin embargo, estas hormonas no han sido vinculadas con procesos de represión de la actividad de estas proteínas, observado en los tiempos posteriores.
Los resultados sugieren un importante papel de la sistemina en su interacción con laμ simbiosis micorrízica arbuscular, al modular las respuestas de defensa durante el proceso de establecimiento de la micorrización, con un efecto a largo plazo, de forma que se logra potenciar la simbiosis por un mecanismo desconocido que podría implicar la regulación de las proteínas PRs.
BIBLIOGRAFÍA
1. Berruti A, Borriello R, Lumini E, Scariot V, Bianciotto V, Balestrini R. Application of laser microdissection to identify the mycorrhizal fungi that establish arbuscules inside root cells. Frontiers in Plant Science. 2013;4:10. doi: 10.3389/fpls.2013.00135.
2. Campos-Soriano L, García-Martínez J, San-Segundo B. The arbuscular mycorrhizal symbiosis promotes the sisteminatemic induction of regulatory defence-related genes in rice leaves and confers resistance to pathogen infection. Mol Plant Pathol. 2012;13(6):579–92. doi: 10.1111/J.1364-3703.2011.00773.X.
3. Evelin H, Kapoor R. Arbuscular mycorrhizal symbiosis modulates antioxidant response in salt-stressed Trigonella foenum-graecum plants. Mycorrhiza. 2014;24:197–208. doi: 10.1007/s00572-013-0529-4.
4. Balasubramanian V, Vashisht D, Cletus J, Sakthivel N. Plant b-13-glucanases: their biological functions and transgenic expression against phytopathogenic fungi. Biotechnol Lett. 2012;34:1983-90. doi: 10.1007/s10529-012-1012-6.
5. Jung SC, Martinez-Medina A, López-Ráez JA, Pozo MJ. Mycorrhiza-induced resistance and priming of plant defenses. J Chem Ecol. 2012;38:651–64. doi: 10.1007/s10886-012-0134-6.
6. Huffaker A, Pearce G, Veyrat N, Erb M, Turlings TCJ, Sartor R, Shen Z, Briggs SP, Vaughan MM, Alborn HT, Teal PEA, Schmelz E. Plant elicitor peptides are conserved signals regulating direct and indirect antiherbivore defense. PNAS. 2013;110(14):5707–12. doi: 10.1073/pnas.1214668110.
7. Coppola M, Corrado G, Coppola V, Cascone P, Martinelli R, Digilio MC, Pennacchio F, Rao R. Prosystemin overexpression in tomato enhances resistance to different biotic stresses by activating genes of multiple. Signaling Pathways. Plant Molecular Biology Report. 2015;3:1270–85. doi: 10.1007/s11105-014-0834-x.
8. Savatin DV, Gramegna G, Modesti V, Cervone F. Wounding in the plant tissue: the defense of a dangerous passage. Front Plant Sci. 2014;16:470-81. doi: 10.3389/fpls.2014.00470.
9. Rehrig EM, Appel H, Jones AD, Schultz CJ. Roles for jasmonate- and ethylene-induced transcription factors in the ability of Arabidopsis to respond differentially to damage caused by two insect herbivores. Front Plant Sci. 2014;5:407-21. doi: 10.3389/fpls.2014.00407.
10. de la Noval B, Pérez E, Martínez B, León O, Martínez N, Délano J. Exogenous sisteminatemin has a contrasting effect on disease resistance in mycorrhizal tomato (Solanum lycopersicum) plants infected with necrotrophic or hemibiotrophic pathogens. Mycorrrhiza. 2007;17:449-60. doi: 10.1007/s00572-007-0122-9.
11. Álvarez M, de Armas G, Martínez B. Informe de nuevas variedades. Amalia y Mariela dos nuevas variedades de tomate de consumo fresco. Cultivos Tropicales. 1997;18(1):83.
12. Schüßler A, Walker C. The Glomeromycota: A species list with new families and new genera [Internet]. CreateSpace Independent Publishing Platform; 2011 [cited 2017 Mar 14]. 58 p. Available from: https://www.amazon.com/Glomeromycota-species-list-families-genera/dp/1466388048
13. Paneque PVM, Calaña NJM, Calderón VM, Borges BY, Hernández GTC, Caruncho CM. Manual de técnicas analíticas para análisis de suelo, foliar, abonos orgánicos y fertilizantes químicos [Internet]. 1st ed. La Habana, Cuba: Ediciones INCA; 2010 [cited 2016 Jan 27]. 157 p. Available from: http://mst.ama.cu/578/
14. Fernández F, Gómez R, Vanegas LF, de la Noval BM, Martínez MA. Producto inoculante micorrizógeno. La Habana, Cuba; 22641, 2000.
15. Bradford M. A rapid and sensitive method for the determination of microgram quantities of protein utilizing the principle of protein dye-binding. Anal Biochem. 1976;72(1-2):248-54. doi: 10.1016/0003-2697(76)90527-3.
16. Tipton K. Principles of enzyme assays and kinetic studies. In: Eisenthal R, Danson MJ, editors. Enzyme Assays: a practical approach. UK: Oxford University Press; 1993. p. 1–58.
17. Statistical Graphics Crop. STATGRAPHICS® Plus [Internet]. 2000. (Profesional). Available from: http://www.statgraphics.com/statgraphics/statgraphics.nsf/pd/pdpricing
18. Rodríguez-Yon Y, de la Noval-Pons B, Fernández-Martín F, Rodríguez-Hernández P. Estudio comparativo del comportamiento de seis cepas de hongos micorrízicos arbusculares en su interacción con el tomate (Lycopersicon esculentum M. var “Amalia”). Ecol. Apl. 2004;3(1-2):162-71.
19. Bitterlich M, Krügel U, Boldt-Burisch K, Franken P, Kühn C. The sucrose transporter SlSUT2 from tomato interacts with brassinosteroid functioning and affects arbuscular mycorrhiza formation. Plant Journal. 2014;78:877–889. doi: 10.4161/15592316.2014.970426.
20. Pozo MJ, López-Ráez JA, Azcón-Aguilar C, García-Garrido JM. Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytol. 2015. doi: 10.1111/nph.13252.
21. Fernández I, Merlos M, López-Ráez JA, Martínez-Medina A, Ferrol N, Azcón C, Bonfante P, Flors V, Pozo MJ. Defense related phytohormones regulation in arbuscular mycorrhizal symbioses depends on the partner genotypes. J. Chemical Ecol. 2014;40:791–803. doi: 10.1007/s10886-014-0473-6.
22. Wasternack C, Hause B. Jasmonates: biosynthesis perception signal transduction and action in plant stress response growth and development. An update to the 2007 review in Annals of Botany. Annals of Botany. 2013;1:1-38. doi: 10.1093/aob/mct067.
23. Leon-Morcillo RJ, Martin-Rodríguez JA, Vierheilig H, Ocampo JA, García-Garrido JM. Late activation of the 9-oxylipin pathway during arbuscular mycorrhiza formation in tomato and its regulation by jasmonate signaling. J Exp Bot. 2012;63(10):3545-58. doi: 10.1093/jxb/ers010.
24. Chen YL, Lee CY, Cheng KT, Chang WH, Huang RN, Gil NH, Chen YR. Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell. 2014;26:4135–48. doi: 10.1105/tpc.114.131185.
25. Agrios GN. Plant Pathology. 5th ed. San Diego: Academic Press; 2005. 922 p.
26. Salzer P, Boller T. Elicitor induced reactions in mycorrhizae and their suppression. In: Current Advances in Mycorrhizae Research. Secction I: Signaling mechanisms in mycorrhizal symbiosis. USA: APS Press; 2000. p. 1-10.
27. Cameron DD, Neal AL, van Wees SCM, Ton J. Mycorrhiza-induced resistance: more than the sum of its parts? Trends Plant Sci. 2013;18(10):539–45. doi: 10.1016/j.tplants.2013.06.004.
28. Pozo MJ, Azcón-Aguilar C. Unraveling mycorrhiza-induced resistance. Curr Opin Plant Biol. 2007;10:393-8. doi: 10.1016/j.pbi.2007.05.004.
29. Yan L, Zhai Q, Wei J, Li S, Wang B, Huang T, Du M, Sun J, Kang L, Li CB, Li C. Role of tomato lipoxygenase D in wound-induced jasmonate biosynthesis and plant immunity to insect herbivores. PLoS Genetic. 2013;9(12):16 doi: 10.1371/journal.pgen.1003964.
30. Ballhorn DJ, Younginger BS, Kautz S. An aboveground pathogen inhibits belowground rhizobia and arbuscular mycorrhizal fungi in phaseolus vulgaris. BMC Plant Biology. 2014;14:321-34. doi: 10.1186/s12870-014-0321-4.
Recibido: 12/01/2016
Aceptado: 06/02/2017
Dra.C. Blanca M. de la Noval-Pons, Instituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700. Email: bdelanov@inca.edu.cu
]]>{kind=link}
{kind=link}
{kind=link}