Desarrollo de las raíces como indicador de la sustentabilidad del suelo de los sistemas de cultivo en el Bioma de la Mata Atlántica
Root development as an indicator of soil sustainability of cropping systems in the Atlantic Forest Biome
Leocimara Sutil de Oliveira-Pessoa Paes, Marcelo Ricardo de Lima, Glaciela Kaschuk, Fabiane Machado-Vezzani
Departamento de Solos e Engenharia Agrícola, Universidade Federal do Paraná. Rua dos Funcionários, 1540, Curitiba, Paraná, Brazil, Postcode: 80.035-050.
]]>
RESUMEN
La sustentabilidad agrícola depende de la funcionalidad del suelo en mecanismos que suporten la productividad vegetal y animal, el ciclo del agua y de los nutrientes y el tamponamiento de contaminación. Para tanto, las muestras de raíces del suelo de dos sistemas agrícolas típicos en el bioma de la Mata Atlántica fueron colectadas para evaluar su sustentabilidad a través de la relación entre raíces y funcionalidad del suelo. Los sistemas agrícolas eran cultivo anuales, maíz (Zea mays L.) y perennes palmeras, (Bactris gasipaes K.). Las raíces fueron colectadas de monolitos (48 x 30 x 5 cm) y lavadas suavemente. Los perfiles de raíz fueron digitalizados y después analizados con los softwares IDRISI y SAFIRA. Las muestras del suelo fueron sometidas a evaluaciones de biomasa microbiana-C y atributos físicos y químicos. El cultivo de palmeras produjo más materia seca de raíces y tuvo mayor área y volumen de raíz que el campo de maíz (cf. 1,37 contra 0,14 t ha-1). Considerando la camada de 0 a 5 cm, el campo de palmeras sostuvo mayor peso medio de diámetro de los agregados del suelo (4,2 contra 3,4 mm) y mantuvo mayor saturación de bases (65 contra 47 %) que el maíz. Aunque el cultivo de palmeras no tenga una cantidad total de C orgánico total (20 contra 24 g kg-1), estimuló la biomasa microbiana (MB-C; 942 versus 428 mg MB-C kg-1) y disminuyó el cociente metabólico (0,023 contra 0,034 mg C-CO2 g-1 MB-C h-1) en relación al cultivo de maíz. Esta relación clara entre los atributos de la raíz y del suelo indica que un mejor desarrollo de raíces contribuyó para mejorar la funcionalidad del suelo y, consecuentemente, la sustentabilidad de los sistemas agrícolas. Por tanto, los sistemas de culturas y manejo del suelo que privilegian el crecimiento de las raíces son mejores opciones para alcanzar la sustentabilidad agrícola.
Palabras clave: actividad biológica en el suelo, agregados del suelo, materia orgánica del suelo.
ABSTRACT
Agricultural sustainability depends on the soil functionality in mechanisms that support plant and animal productivity, water and nutrient cycling, and contamination buffering. Therefore, roots and soil samples from two typical agricultural systems in the Atlantic Forest biome were collected, to evaluate their sustainability through the relationship between roots and soil functionality. The agricultural systems were annual fields as, maize (Zea mays L.) and perennial, palm (Bactris gasipaes K.). The roots were collected from monoliths (48 x 30 x 5 cm) and after, they were gently washed. The root profiles with the IDRISI and SAFIRA softwares were scanned and then analyzed. Soil samples were submitted to evaluations of microbial-C biomass and physical and chemical attributes. The palm field produced more dry matter roots and had greater root area and volume than the maize field (cf. 1.37 versus 0.14 t ha-1). Considering the 0-5 cm layer, the palm field sustained larger mean weight diameter of soil aggregates (4.2 versus 3.4 mm) and held higher saturation of bases (65 versus 47 %) than the maize. Although the palm field did not have larger total organic C content (20 versus 24 g kg-1), it stimulated microbial biomass (MB-C; 942 versus 428 mg MB-C kg-1 soil) and decreased metabolic quotient (0.023 versus 0.034 mg C-CO2 g-1 MB-C h-1) in relation to maize field. This clear relationship between root and soil attributes indicates that better root development contributes to improve soil functionality and for consequence the sustainability of agricultural systems. Therefore, crop systems and soil managements that privilege root growth are better choice to reach the agricultural sustainability.
Key words: biological activity in soil, soil aggregates, soil organic matter.
INTRODUCCIÓN
]]> La sostenibilidad de los sistemas agrícolas depende de la funcionalidad del suelo que respalde la productividad de las plantas y los animales, el ciclo del agua y de los nutrientes, así como el tamponamiento intermedio de la contaminación. La funcionalidad del suelo es el resultado de la interacción de minerales, organismos del suelo y plantas (1) y esta interacción es impulsada por las raíces (2). Las raíces agregan carbono al suelo al descargar exudados y desprendimiento de raíz (3). Por lo tanto, el carbono agregado estimula a los organismos que desempeñan papeles cruciales en procesos como la descomposición de la materia orgánica del suelo, el ciclo de nutrientes, la degradación de los pesticidas y controla la formación y estabilización de la agregación de origen biológico (4-7).En este estudio, probamos la hipótesis de que la sostenibilidad ecológica del suelo de los sistemas agrícolas es una función de los atributos de la raíz (abundancia y composición). Las raíces proporcionan una fuente más duradera de sustratos para el crecimiento microbiano ya que tienen más lignina y celulosa que los brotes (7). La exudación de raíces y los subproductos estimulan el crecimiento microbiano en la rizosfera, afectando la disponibilidad de nutrientes del suelo y la composición de la materia orgánica (5). Además, a través de las interacciones organominerales, la exudación de la raíz contribuye a la formación de microagregados (<0,25 mm), que se unen en macroagregados (> 0,250 mm) por micelios y segmentos de raíz (6,7). Las interacciones de la planta, los organismos del suelo, particularmente los microorganismos y las partículas organominerales del suelo cooperan para mejorar la funcionalidad del suelo (1) y en consecuencia, contribuyen a la sostenibilidad de los sistemas agrícolas.
El bioma forestal atlántico de Brasil abarca 1,110,182 km2 y se considera un punto clave para la conservación de la biodiversidad; sin embargo, cerca del 90 % de su extensión ha sido destruida por la urbanización, la agricultura y las actividades mineras (8) y se requieren esfuerzos mayores para establecer estrategias que mejoren la sostenibilidad y productividad del suelo en áreas ya antropogenizadas, permitiendo que los bosques restantes se conserven completamente. Con la confirmación de que los atributos de la raíz rigen las funciones ecológicas del suelo, los agrónomos podrían diseñar mejores estrategias para mejorar la funcionalidad del suelo y la sostenibilidad de los sistemas agrícolas, por ejemplo, eligiendo especies de plantas, densidades de siembra y arreglos.Nuestro objetivo fue evaluar la relación entre el desarrollo de raíces de los sistemas de cultivo anuales y perennes y los atributos biológicos, físicos y químicos del suelo a la luz de la mejora de la sostenibilidad para indicar las direcciones de gestión de los sistemas agrícolas de la región.
MATERIALES Y MÉTODOS
Sitio de campo y muestreo
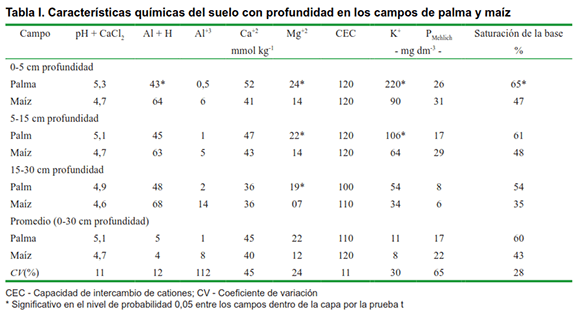
El estudio se realizó en una granja en las coordenadas 25º16’S y 48º42’W en la costa norte del estado de Paraná, en el bioma forestal del sur de la Mata Atlántica, Brasil. El clima es Cfa subtropical según la clasificación de Koppen, con una temperatura media anual de 22 0C y una variación de 10 a 18 0C en los meses más fríos (9). La precipitación promedio anual es de 2,587 mm por año, con un promedio de 208 días lluviosos por año. La granja está en un suelo Cambisol según la FAO, que consiste en un 49 % de arcilla, un 46 % de limo y un 5 % de arena (10). Las características químicas del suelo de los sitios de campo se presentan en la Tabla I.
Se tomaron cuatro muestras compuestas de dos campos experimentales, con 180 m de separación y orientadas una hacia la otra, elegidas para representar la composición de raíces agrícolas variadas y la abundancia del bioma de la Mata Atlántica. Los campos experimentales se describen de la siguiente manera: 1) cultivo perenne de palmeras (Bactris gasipaes K.) cultivado bajo labranza cero durante 16 meses, espaciado entre hileras de 1,40 m, espaciamiento entre plantas de 0,80 m, que comprende 7,142 plantas ha- 1; de ahora en adelante referido como palma; 2) cultivo anual con maíz (Zea mays L.), espaciado entre hileras de 1,40 m, espaciamiento entre plantas de 0,55 m, que comprende 17,647 plantas ha-1, bajo labranza convencional (arado y rastrillado rotativo semanal a 15 cm de profundidad) por dos temporadas de crecimiento; en adelante referido como maíz. Los fertilizantes se aplicaron siguiendo las recomendaciones oficiales para cada cultivo (11) siendo para maíz 40 kg N ha-1, 20 kg P2O5 ha-1 y 80 kg K2O ha-1; y para la palma 80 kg N ha-1, 20 kg P2O5 ha-1 y 80 kg K2O ha-1.Las malezas fueron controladas por azada rotativa (cultivo anual) y siega (cultivo perenne). La enfermedad de las plantas no se identificó durante el período de estudio y los cultivos no fueron irrigados. Se recolectaron muestras en cada campo experimental durante los meses de marzo de 2014, cuando el maíz estaba en la etapa de llenado de granos y las palmeras tenían 16 meses de edad.
Raíz de abundancia y composición
]]> Se obtuvieron cuatro muestras de raíces cavando monolitos con dimensiones de 48×30×5 cm (largo × alto × ancho) (12). Los monolitos se lavaron suavemente con una corriente de agua hasta que se eliminó la suciedad y los perfiles de raíz se fotografiaron con una cámara digital (Nikon Coolpix P90). Más tarde, las imágenes de perfil se analizaron con el software IDRISI Selva 17.0 (Clark Labs). Las raíces muestreadas también se separaron de acuerdo con sus capas de profundidad originales (0-5, 5-15 y 15-30 cm) y se escanearon con un escáner portátil (Lexmark 1200 Series) a 200 ppp. Las imágenes se analizaron con el software SAFIRA para el cálculo de la longitud, densidad, área, volumen y diámetro de las raíces. La densidad de longitud de la raíz se obtuvo dividiendo la longitud de la raíz por volumen de suelo en cada profundidad. Los contenidos de carbono y nitrógeno en la materia seca de la raíz se determinaron mediante combustión seca utilizando un analizador de CN (Vario El III - Elementar®). La lignina se determinó con la metodología descrita por Morais et al., (13). No se proporcionan estadísticas para las mediciones de lignina porque quedaba muy poco material de raíz y su determinación solo podía lograrse al componer todas las réplicas.Análisis químico y físico del suelo
Las muestras de suelo se tomaron con un cuchillo de acero inoxidable en las capas 0-5, 5-15 y 15-30 cm de profundidad, cerca de cada uno de los cuatro sitios de muestreo de raíz. Los análisis químicos y físicos del suelo siguieron los procedimientos estándar de la siguiente manera: química (pH en CaCl2, Al + H, Ca2+, Mg2+, K+, PMehlich, carbono orgánico total y nitrógeno total) con la metodología descrita por Marques y Motta (14); polisacáridos totales del suelo (15); y la distribución del tamaño del agregado húmedo en clases de diámetro y el diámetro medio del peso (16) modificado utilizando muestras de áridos no humidificadas previamente (proceso de extinción).
Indicadores microbianos del suelo
Se midió la respiración microbiana del suelo (17) y se determinó la biomasa microbiana del suelo (MB-C) (18). El cociente microbiano (qMic) se estimó como la relación entre MB-C y C orgánico total y el cociente metabólico (qCO2) como relación entre la respiración microbiana del suelo y MB-C.
Análisis estadístico
Primero se verificaron los datos de normalidad y homogeneidad con las pruebas de Shapiro-Wilk y Bartlett. Se aplicó ANOVA de una vía considerando muestras de un diseño completamente aleatorizado que compara cada parámetro dentro de su capa. En los casos en que se detectó significación estadística, los promedios se compararon con la prueba de Tukey a p <0,05.
RESULTADOS Y DISCUSIÓN
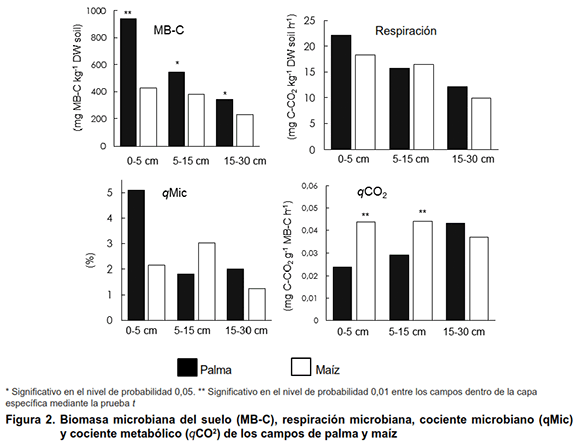
El campo de palma tenía arreglos de raíces más densos y su perfil de raíz contenía fragmentos distribuidos más uniformemente a través de direcciones laterales y verticales en comparación con el maíz (Figura 1). Junto con la Figura 1, la Tabla II muestra que el campo de palma tenía una mayor área de raíz y volumen que el campo de maíz. En ambos campos, la mayor proporción de raíces se formó por raíces muy finas, que comprenden fragmentos con diámetros menores de 0,5 mm. Al considerar la fracción de raíces finas (0,5-2,0 mm), la palma tenía más segmentos de raíz significativos que el maíz (Tabla II). Proporciones más grandes de raíces finas favorecen la funcionalidad del suelo porque las mismas permiten que las plantas exploten los nutrientes en volúmenes de suelo más grandes (6,7). Las raíces finas son importantes para el ciclo de nutrientes porque tienen una tasa de descomposición más rápida que otros desechos orgánicos (19).
]]>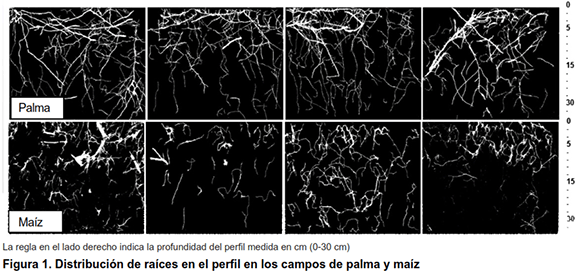
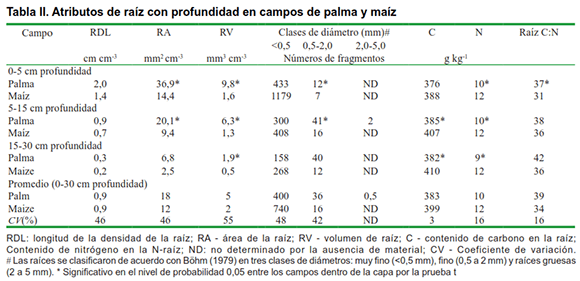
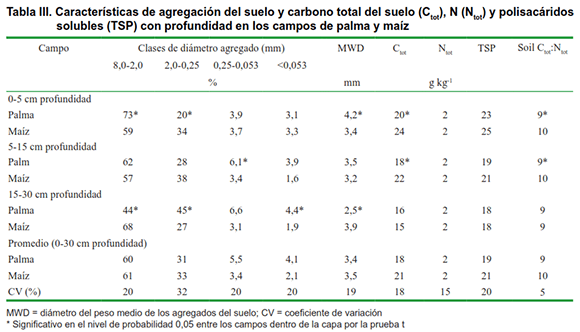
Al ensamblar todo el material (en todas las profundidades desde 0-30 cm), calculamos que el campo de palma formó 1,37 t ha-1 de raíces de materia seca mientras que el campo de maíz se formó aproximadamente 10 veces menos (0,14 t ha-1). La menor cantidad de raíces de materia seca es consecuencia de las líneas de hileras anchas y la baja densidad de plantas en el campo de maíz, que es típico del tipo de sistemas agrícolas en la región estudiada. La palma tenía la relación brote / raíz más pequeña (9,9) que el maíz (13,6), y la materia seca de las raíces superiores del campo de la palma, especialmente en las capas superiores (Tabla II), promovió el carbono de la biomasa microbiana del suelo (Figura 2). Curiosamente, el cociente metabólico en la profundidad de 0-5 y 5-15 cm fue significativamente menor en el campo de palma en comparación con el maíz. Hubo una tendencia a que la respiración microbiana del suelo disminuye con la profundidad, pero no hubo diferencias significativas en la respiración y el cociente microbiano entre los campos.
Más raíces y carbono de biomasa microbiana se asociaron con una mayor proporción de clases más grandes de diámetro agregado en la capa superior en el campo de palma, es decir, el diámetro medio del peso fue de 4,2 mm en el campo de palma y 3,4 mm en el campo de maíz en el 0-5 cm capa de profundidad (Tabla III). Estos resultados apoyan nuestra hipótesis de que las raíces juegan un papel importante en el control de los atributos microbiológicos y físicos del suelo que determinan la funcionalidad del suelo y la sostenibilidad del sistema agrícola. Tisdall & Oades (3) explicaron que los macroagregados del suelo (>0,250 mm) se forman durante el crecimiento y la exudación de raíces y microbios (particularmente hongos).
Aunque no hubo diferencias en el nitrógeno total del suelo, el C orgánico total en el campo de la palma fue significativamente más bajo que el del maíz (Tabla III). Indicó que el contenido de lignina es uno de los factores más importantes para estabilizar el carbono rhizodepositado en el suelo (20). Considerando que las raíces cosechadas en el campo de palma contenían solo 140 mg de lignina g-1 de raíces mientras que las raíces de maíz contenían 300 mg g-1, podemos suponer que la composición de raíz (contenido de lignina) contribuyó a evitar el carbono orgánico total por descomposición microbiana en el campo de maíz curiosamente, los resultados del carbono orgánico total, podrían estar contradiciendo el hecho de que un mejor desarrollo de la raíz da como resultado un aumento del carbono orgánico total del suelo (21) (Tabla III). Sin embargo, aunque el campo de palma perenne no tenía un mayor contenido de carbono orgánico total (Tabla III), admitió una mayor biomasa microbiana de carbono y un cociente metabólico más bajo que los campos anuales de maíz (Figura 2). Esa es una fuerte indicación de que el campo del maíz podría estar perdiendo Ctot con el tiempo, debido al mayor cociente metabólico, que a menudo se asocia con la labranza convencional (22), típica en los sistemas agrícolas de la región (Figura 2).
El carbono orgánico total y la proporción de carbono total de nitrógeno (Ctot: Ntot) de los residuos de raíz (sustratos microbianos potenciales) del campo de palma fue ligeramente mayor que el del campo de maíz (Tabla II), aunque solo fue significativo en la capa superior (0- 5 cm de profundidad), pero las relaciones Ctot: Ntot del suelo fueron más bajas (Tabla III). De forma similar a lo que se ha observado (23), encontramos que el cociente metabólico se relacionó positivamente con las relaciones Ctot: Ntot del suelo (Tabla III, Figura 2). Varios estudios han demostrado que aumentar la diversidad de carbono orgánico (según la rotación de cultivos en comparación con el monocultivo) y la acumulación de carbono a largo plazo estimula el desarrollo de comunidades microbianas que son mucho más eficientes en la utilización de carbono, lo que resulta en un mayor cociente microbiano y menor cociente metabólico (4, 24). En nuestro estudio, parece que el manejo perenne de la palmera produce cambios en la funcionalidad del suelo al diversificar los sistemas radiculares (raíces de palma y hierba), cambiar las relaciones Ctot: Ntot del suelo y estimular un uso más eficiente de los recursos de carbono, lo que podría conducir al sistema de sostenibilidad. De igual forma estos, concluyeron en su estudio que el cultivo perenne de cereales es una forma de lograr la sostenibilidad agrícola (25).
Hubo ahorros de nutrientes (como más Mg y K) en el suelo del campo de palma (Tabla I). La saturación de bases fue del 65 % en el campo de palma y del 47 % en el campo de maíz. Es probable que los sistemas que tienen una adición continua de carbono por materia seca de brotes y raíces conserven más nutrientes en la biomasa microbiana del suelo. La biomasa microbiana del suelo juega un papel importante como fuente y sumidero de nutrientes (24) y el aumento de la misma resulta en efectos positivos sobre la fertilidad y la funcionalidad del suelo (1,26).Como las raíces depositan exudados y tejidos muertos, estimulan la actividad biológica del suelo y la construcción de biomasa microbiana de carbono, que a su vez promueve la agregación de partículas al mismo y mejora las características químicas con el resultado final de una mejor funcionalidad de este. Entonces, la funcionalidad del suelo y el sistema de sostenibilidad pueden mejorarse estimulando el crecimiento de las raíces. Esto podría lograrse rotando o intercalando plantas con diferentes tipos de raíces o eligiendo manejo del suelo que no dañe el crecimiento de la raíz.
CONCLUSIÓN
]]> El cultivo perenne (palma) produjo más raíces de materia seca y tenía un mayor área y volumen de raíces que el cultivo anual (maíz). En la capa de la superficie del suelo, los cultivos perennes construyeron una mejor estructura física y química y promovieron los atributos biológicos del suelo más que los cultivos anuales. Esta clara relación entre los atributos de raíz y suelo indica que un mejor desarrollo de la raíz contribuye a mejorar la funcionalidad del suelo y por consiguiente, la sostenibilidad de los sistemas agrícolas.
BIBLIOGRAFÍA
1. Prado RB, Fidalgo ECC, Monteiro JMG, Schuler AE, Vezzani FM, Garcia JR, et al. Current overview and potential applications of the soil ecosystem services approach in Brazil. Pesquisa Agropecuária Brasileira. 2016;51:1021–38. doi:10.1590/s0100-204x2016000900002
2. Blankinship JC, Fonte SJ, Six J, Schimel JP. Plant versus microbial controls on soil aggregate stability in a seasonally dry ecosystem. Geoderma. 2016;272:39–50. doi:10.1016/j.geoderma.2016.03.008
3. Scott DA, Baer SG, Blair JM. Recovery and relative influence of root, microbial, and Structural properties of soil on physically sequestered carbon stocks in Restored Grassland. Soil Science Society of America Journal. 2017;81:50-. doi:10.2136/sssaj2016.05.0158
4. Duchene O, Vian J-F, Celette F. Intercropping with legume for agroecological cropping systems: Complementarity and facilitation processes and the importance of soil microorganisms. A review. Agriculture, Ecosystems & Environment. 2017;240:148–61. doi:10.1016/j.agee.2017.02.019
5. Tisdall JM, Oades J. Organic matter and water-stable aggregates in soils. European Journal of Soil Science. 1982;33(2):141–63. doi:10.1111/j.1365-2389.1982.tb01755.x
6. Six J, Bossuyt H, Degryze S, Denef K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil and Tillage Research. 2004;79(1):7–31. doi:10.1016/j.still.2004.03.008
7. Dignac M-F, Derrien D, Barré P, Barot S, Cécillon L, Chenu C, et al. Increasing soil carbon storage: mechanisms, effects of agricultural practices and proxies. A review. Agronomy for Sustainable Development. 2017;37(2):14. doi:10.1007/s13593-017-0421-2
8. Forzza RC, Baumgratz JFA, Bicudo CEM, Canhos DA, Carvalho Jr AA, Coelho MAN, et al. New brazilian floristic list highlights conservation challenges. BioScience. 2012;62(1):39–45. doi:10.1525/bio.2012.62.1.8
9. Alvares CA, Stape JL, Sentelhas PC, de Moraes Gonçalves JL, Sparovek G. Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift. 2013;22(6):711–28. doi:10.1127/0941-2948/2013/0507
10. FAO – Food and Agriculture Organization of the United Nations. International soil classification system fornaming soils and creating legends for soil maps. World Soil Resources Reports No. 106. FAO, Rome; 2014.
11. Sociedade Brasileira de Ciência do Solo. Manual de Adubação e Calagem para o Estado do Paraná [Internet]. Curitiba, Brasil: Núcleo Estadual do Paraná; 2017 [cited 2018 Apr 4]. 482 p. Available from: https://www.sbcs.org.br/loja/index.php?route=product/product&product_id=130
12. Böhm W. Methods of studying root systems [Internet]. Berlin Heidelberg: Springer-Verlag; 1979 [cited 2018 Apr 4]. 188 p. (Ecological Studies). Available from: https://www.springer.com/gp/book/9783642672842
13. Morais JPS, Rosa M de F, Marconcini JM. Procedimentos para análise lignocelulósica. Documento 236 [Internet]. Campina Grande: Embrapa Algodão; 2010 p. 54. Available from: https://www.infoteca.cnptia.embrapa.br/bitstream/doc/883400/1/DOC236.pdf
14. Marques R, Motta ACV. Análise química do solo para fins de fertilidade. In: Lima MR, editor. Manual de diagnóstico da fertilidade e manejo dos solos agrícolas. Universidade Federal do Paraná: 2.ed. Curitiba, Universidade Federal do Paraná; 2003. p. 81–102.
15. Lowe L. Total and labile acid extractable polysaccharide analysis of soils. In: Carter MR, editor [Internet]. Soil Sampling and Methods of Analysis. Lewis Publishers, Canadian Society of Soil Science; 1993. p. 373–6. Available from: https://books.google.com.cu/books?id=54lYLsV49zIC&printsec=frontcover&hl=es#v=onepage&q&f=false
16. Kemper WD, Rosenau RC. Aggregate stability and size distribution. In: 2nd Edition. American Society of Agronomy, Soil Science Society of America; [Internet]. Methods of soil analysis, Part 1. Physical and mineralogical methods-Agronomy Monograph no.9. 1986. p. 425–42. Available from: https://eprints.nwisrl.ars.usda.gov/732/3/585.pdf
17. Alef K. Soil respiration. In: Alef K, Nannipieri P. editors [Internet]. Methods in applied soil microbiology and biochemistry. London?: Academic press, 1995; 1995. p. 214–20. Available from: https://lib.ugent.be/catalog/rug01:000385148
]]> 18. Jenkinson DS, Powlson DS. The effects of biocidal treatments on metabolism in soil—V. Soil Biology and Biochemistry. 1976;8(3):209–13. doi:10.1016/0038-0717(76)90005-519. Violita V, Triadiati T, Anas I, Miftahudin M. Fine Root Production and Decomposition in lowland rainforest and oil palm plantations in Sumatra, Indonesia. HAYATI Journal of Biosciences. 2016;23(1):7–12. doi:10.1016/j.hjb.2015.10.008
20. Panettieri M, Rumpel C, Dignac M-F, Chabbi A. Does grassland introduction into cropping cycles affect carbon dynamics through changes of allocation of soil organic matter within aggregate fractions? Science of The Total Environment. 2017;576:251–63. doi:10.1016/j.scitotenv.2016.10.073
21. Pausch J, Tian J, Riederer M, Kuzyakov Y. Estimation of rhizodeposition at field scale: upscaling of a 14C labeling study. Plant and Soil. 2013;364(1–2):273–85. doi:10.1007/s11104-012-1363-8
22. Zuber SM, Villamil MB. Meta-analysis approach to assess effect of tillage on microbial biomass and enzyme activities. Soil Biology and Biochemistry. 2016;97:176–87. doi:10.1016/j.soilbio.2016.03.011
23. Spohn M, Chodak M. Microbial respiration per unit biomass increases with carbon-to-nutrient ratios in forest soils. Soil Biology and Biochemistry. 2015;81:128–33. doi:10.1016/j.soilbio.2014.11.008
24. Tiemann LK, Grandy AS, Atkinson EE, Marin-Spiotta E, McDaniel MD. Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Hooper D, editor. Ecology Letters. 2015;18(8):761–71. doi:10.1111/ele.12453
25. Marquardt K, Vico G, Glynn C, Weih M, Eksvärd K, Dalin P, et al. Farmer perspectives on introducing perennial cereal in Swedish farming systems: a sustainability analysis of plant traits, farm management, and ecological implications. Agroecology and Sustainable Food Systems. 2016;40:432–50. doi:10.1080/21683565.2016.1141146
26. Delgado-Baquerizo M, Grinyer J, Reich PB, Singh BK. Relative importance of soil properties and microbial community for soil functionality: insights from a microbial swap experiment. Allen E, editor. Functional Ecology. 2016;30(11):1862–73. doi:10.1111/1365-2435.12674
]]>
Recibido: 09/05/2017
Aceptado: 12/02/2018
Fabiane Machado-Vezzani, Departamento de Solos e Engenharia Agrícola, Universidade Federal do Paraná. Rua dos Funcionários, 1540, Curitiba, Paraná, Brazil, Postcode: 80.035-050. Email: fabianevezzani@gmail.com
]]>