Lic. Juan A. Bisset,1 Lic. María M. Rodríguez,2 Lic. Darjaniva Molina,3 Lic. Cristina Díaz4 y Téc. Lázaro A. Soca5
Fueron evaluadas 5 cepas de Aedes aegypti L., 1 de Cuba y 4 de Venezuela, para determinar sus niveles de resistencia a insecticidas organofosforados (temefos, clorpirifos y pirimifos metil). En las cepas venezolanas se encontró resistencia a temefos, solo en la cepa APURE. En las cepas de TACHIRA y MIRANDA se observaron moderados valores de resistencia (FR50 entre 5 y 10 x) para clorpirifos, y altos niveles de resistencia (FR 10x) fueron obtenidos para este insecticida en la cepa de ARAGUA. Todas las cepas venezolanas mostraron elevados niveles de resistencia a pirimifos metil. La cepa cubana, colectada en SANTIAGO de CUBA, mostró moderados niveles de resistencia a temefos y pirimifos metil, y altos niveles para clorpirifos. Los resultados de las pruebas bioquímicas demostraron la presencia de los mecanismos de esterasas y glutation-s-transferasa, a elevada frecuencia en casi todas las cepas. Mediante electroforesis en gel de poliacrilamida se observó una banda fuertemente teñida en todas las cepas, con un valor de Rf de 0,779, que se nombró esterasa A4, y no se observó en la cepa de referencia susceptible a insecticidas. El significado de esta esterasa en la resistencia a insecticidas organofosforados está aún por demostrar. La resistencia a estos insecticidas en Ae. aegypti es un problema serio para las operaciones de control de esta especie, por lo que se sugirieron estrategias de control integradas que permitan evitar o retardar la aparición de esta en Cuba y Venezuela.
DeCs: AEDES/enzimología; ESTERASAS/análisis; INSECTICIDAS ORGANOFOSFORADOS.
Dentro del gran número de especies de mosquitos resistentes a la acción de los insecticidas el género Aedes desempeña un papel importante en la transmisión de enfermedades virales. Comúnmente la fiebre amarilla, el dengue y la fiebre del dengue hemorrágico (FDH) son enfermedades víricas, transmitidas por Aedes aegypti L., que causan grandes impactos en la salud pública.
En 1960 se reportaron los primeros casos de resistencia a insecticidas organofosforados y carbamatos en Ae. aegypti, Fox1 reportó una cepa en Puerto Rico resistente 10x a malation y diazinon. La capacidad de resistir al malation se asoció con la destoxificación mediada por enzimas de actividad específica: carboxilesterasas. El incremento de los niveles de resistencia a temefos está, presuntamente, asociado con este mecanismo; solo que las enzimas destoxificadoras son fosfatasas. El posible papel de las enzimas específicas en el proceso de destoxificación de insecticidas, en Ae. aegypti, fue observado en larvas de la especie.2 Estos estudios sugieren la presencia de enzimas destoxificadoras específicas, que cumplen un importante papel en el mecanismo de resistencia a organofosforados.
En Cuba ocurrió un brote de dengue en el municipio de Santiago de Cuba en el año 1997, y en Venezuela, en los estados de Aragua, Miranda y Apure se reportó alta incidencia de dengue y prevalencia del vector entre los años 1997 y 1998.3 En el presente estudio se colectó Aedes aegypti de estas localidades, y se evaluó el estado de la resistencia a insecticidas organofosforados que están en uso o se piensan utilizar en el futuro, así como sus posibles mecanismos de resistencia.
Para el trabajo se utilizaron 5 cepas de Aedes aegypti: ROCKEFELLER, cepa de referencia susceptible que fue suministrada por el laboratorio del Centro de Control de Enfermedades (CDC) de San Juan de Puerto Rico; las cepas APURE, ARAGUA, MIRANDA Y TACHIRA, colectadas en Venezuela en los estados del mismo nombre y SANTIAGO DE CUBA, cepa de campo colectada en el municipio Santiago de Cuba, durante el último brote de dengue en Cuba en el año 1997. Las colonias fueron establecidas y mantenidas en el insectario del Instituto de Medicina Tropical "Pedro Kourí" (IPK) con temperatura de 25 °C y 65 % de humedad relativa.
Se emplearon los insecticidas siguientes:
]]> (A) Organofosforados:(C) Sinergistas:
Los bioensayos se realizaron siguiendo la metodología de la Organización Mundial de la Salud (OMS).4 Se emplearon 5 réplicas de cada concentración del insecticida (20 larvas por réplica) y se registró entre 2 y 98 % de mortalidad. Todas las soluciones se ajustaron a un volumen final de 1 mL con acetona. Esta concentración de acetona no causó mortalidad en los controles.
]]> La acción de los sinergistas se determinó con la exposición de las larvas a concentraciones subletales de DEF (0,4 mL de 0,01 %) y del PB (0,8 mL de 0,001 %) durante 4 h previas a la adición de los insecticidas. La lectura de las mortalidades se realizó a las 24 h. Se analizaron los resultados mediante el programa probit log de Raymond.5Las pruebas bioquímicas se realizaron en larvas de cuarto estadio temprano. Las actividades de esterasas y glutation-s-transferasa se determinaron individualmente en larvas según el método de Peiris and Hemingway.6 Se determinó la frecuencia génica según la fórmula de equilibrio de Hardy-Weinberg.
Se realizó electroforesis en gel de poliacrilamida (10 %). Se determinó la actividad enzimática de las esterasas y se seleccionaron las muestras con mayor actividad. Se aplicaron 20 mL de la mezcla (10 mL de muestra + 10 mL del indicador xilene cianol) en el gel y se realizó la corrida a 150 V, durante 45 min. Para la tinción de las bandas de esterasas, se sumergieron los geles en 50 mL de buffer fosfato (0,1 mol), que contenía 4 mL de cada uno de los sustratos inespecíficos de las esterasas (a y ß-naftilacetato). Después se añadieron 0,5 g del colorante fast blue RR, disuelto antes en agua destilada y SDS (sodio duodecil sulfato) 5 %.
En la tabla 1 se muestran los valores de las concentraciones letales 50 (CL50) y el factor de resistencia correspondiente (FR50), se muestra además la pendiente de la línea de regresión probit-log para 3 insecticidas organofosforados (temefos, clorpirifos y pirimifos metil), en las 4 cepas venezolanas (APURE, ARAGUA, MIRANDA Y TACHIRA), la cepa cubana (SANTIAGO DE CUBA) y la cepa susceptible de referencia de Aedes aegypti (ROCKEFELLER). Como se muestra en esta tabla, las cepas de Venezuela resultaron susceptibles al insecticida organo-fosforado temefos (FR <5), excepto la cepa APURE, que resultó 11,1 x resistente a este insecticida. Sin embargo resultaron resistentes (FR >10) a pirimifos metil la cepa ARAGUA, que mostró el valor más elevado de resistencia (27,6 x) y resultó ser la cepa con mayor FR a clorpirifos (21,83 x), y la cepa de MIRANDA y TACHIRA mostraron niveles moderados para este insecticida, 5,67 y 8,29 x, respectivamente. Sin embargo para la cepa de SANTIAGO DE CUBA se obtuvieron moderados niveles de resistencia a temefos (5,59 x) y pirimifos metil (8,03 x), y altos valores de resistencia se evidenciaron para clorpirifos (16,011 x).
Tabla 1. Niveles de resistencia a los insecticidas organofosforados: clorpirifos, temefos y pirimifos metil en 5 cepas de Aedes aegypti
|
| ]]> Pirimifos | ||
|
|
Temefos |
metil | Clorpirifos |
| SANTIAGO DE CUBA CL50a | 0,0713 | 0,0641 | ]]>
0,114 |
|
|
(0,0601-0,0763) |
(0,060-0,0683) |
(0,103-0,140) |
| bb | ]]> 7,2 (± 0,71) |
6,5 (± 0,59) |
6,7 (± 1,42) |
| FR50c | 5,9 | 8,1 | 16,2 |
| APURE CL50a | ]]> 0,149 | 0,140 | 0,0212 |
| bb | (0,0116-0,0180) | (0,127-0,157) | (0,0184-0,0239) |
|
|
2,2 (± 0,22) | ]]>
5,1 (± 0,51) | 2,9 (± 0,23) |
| FR50c | 11,1 |
17,8 |
3,1 |
| ARAGUA CL50a | ]]> 0,0468 |
0,224 | 0,150 |
|
|
(0,0424-0,0514) | (0,193-0,256) |
(0,121-0,201) |
| ]]> bb | 5,2 (± 0,69) |
3,9 (± 0,40) | 1,8 (± 0,25) |
| FR50c | 3,9 |
27,6 | ]]> 22,1 |
| MIRANDA CL50a | 0,0559 | 0,170 |
0,0387 |
|
|
(0,0510-0,0619) | (0,0716-0,229) | ]]> (0,0321-0,0468) |
| bb |
4,5 (± 0,56) | 3,6 (± 0,91) | 2,3 (± 0,29) |
| FR50c | 4,7 | 21,5 | ]]> 5,7 |
| TACHIRA CL50a | 0,0310 | 0,097 | 0,057 |
|
|
(0,028-0,033) |
(0,071-011) | ]]> (0,051-0,062) |
| bb | 2,55 (± 0,63) |
5,086 (± 0,71) |
2,43 (± 0,53) |
| FR50c | 2,58 | 12,28 | ]]> 8,38 |
| ROCKEFELLER CL50a | 0,0127 | 0,00701 | 0,00687 |
|
|
(0,0116-0,0144) |
(0,00698-0,00912) | ]]>
(0,00619-0,0075) |
| bb | 6,3 (± 0,73) |
3,6 (± 0,52) | 5,1 (± 0,82) |
Tabla 2. Valores de las CL50 y factor de sinergismo (FS) para insecticidas organofosforados, con la utilización del sinergista DEF, en 5 cepas de Aedes aegypti
|
|
Pirimifos | ||
|
| ]]> Temefos | metil | Clorpirifos |
| SANTIAGO DE CUBA LC50a | 0,0112 | 0,0981 |
0,00678 |
|
| ]]> (0,0103-0,0130) | (0,0914-0,109) | (0,00638-0,00718) |
| bb | 7,7 (± 1,40) | 6,6 (± 1,68) | 7,4 (± 0,74) |
| FSc | 6,45 | ]]> 0,65 | 16,2 |
| APURE LC50a | 0,00764 | 0,052 | 0,00538 |
|
|
(0,00708-0,00824) | (0,0490-0,0560) | ]]>
(0,00496-0,00571) |
| bb | 5,43 (± 0,70) | 9,9 (± 1,53) | 7,5 (± 0,92) |
| FSc | 19,7 | 2,7 | 3,9 |
| ]]> ARAGUA LC50a | 0,00918 | 0,0522 | 0,00049 |
|
|
(0,00808-0,0108) |
(0,0454-0,0591) | (0,00038-0,00064) |
| ]]> bb | 4,6 (± 0,68) | 3,2 (± 0,45) |
1,6 (± 0,25) |
| FSc | 5,1 |
4,3 | 306,1 |
| ]]> MIRANDA LC50a | 0,00524 | 0,0268 | 0,00378 |
|
|
(0,00476-0,00563) |
(0,0227-0,0310) | ]]> (0,00311-0,00490) |
| bb | 7,1 (± 0,91) |
3,8 (± 0,32) | 2,02 (± 0,26) |
| FSc | 10,7 | 6,3 | 10,2 |
| ]]> TACHIRA LC50a | 0,0064 |
0,049 |
0,0039 |
|
|
(0,0059-0,0068) | (0,041-0,052) | ]]>
(0,0032-0,0041) |
| bb | 8,2 (± 0,93) | 5,38 (± 0,81) | 5,085 |
| FSc | 4,84 | 1,98 | ]]> 14,6 |
| ROCK LC50a | 0,0121 |
0,0121 |
0,00401 |
|
|
(0,0102-0,0161) | ]]>
(0,011-0,014) | (0,004-32-00,565) |
| bb | 4,6 (± 0,98) | 6,4 (± 0,64) |
4,4 (± 0,44) |
| FSc | 1,0 | ]]> 0,64 | 1,5 |
a CL50 en mg/L: 95 % IC están entre paréntesis.
b Pendiente de la recta probit-log. La desviación estándar (±DE) está entre paréntesis.
c Factor de sinergismo (FS): CL50 del insecticida solo/CL50 del insecticida + sinergista. Número de larvas evaluadas: 500 por insecticidas.
Los resultados del FS obtenidos con el sinergista PB, indicaron que las enzimas multifunción oxidasas (MFO) no desempeñaron un papel importante en la resistencia detectada a clorpirifos, temefos y pirimifos metil (tabla 3).
Tabla 3. Valores de las CL50 y factor de sinergismo (FS), para 3 insecticidas organofosforados, con la utilización del sinergista PB, en Aedes aegypti
]]>
|
|
Pirimifos | ||
|
|
Temefos |
metil | Clorpirifos |
| ]]> SANTIAGO DE CUBA LC50a | 0,0885 | 0,181 |
0,0861 |
|
|
(0,0838-0,0943) |
(0,157-0,205) | ]]>
(0,0786-0,0949) |
| bb | 7,9 (± 1,11) | 4,6 (± 0,42) | 4,2 (± 0,49) |
| FSc | 0,8 | 0,35 | 1,3 |
| ]]> APURE LC50a | 0,0377 |
0,0758 |
0,0296 |
|
|
(0,0337-0,0416) | ]]> (0,0606-0,0839) |
(0,0262-0,0329) |
| bb | 3,9 (± 0,34) | 4,9 (± 1,26) | 3,9 (± 0,38) |
| FSc | 3,9 | 2,7 | ]]> 0,7 |
| ARAGUA LC50a | 0,0420 |
0,151 |
0,0457 |
|
|
(0,0379-0,0461) | ]]> (0,138-0,167) | (0,0402-0,0508) |
| bb | 5,4 (± 0,71) | 5,4 (± 0,46) | 3,9 (± 0,48) |
| FSc | 1,1 | 1,5 | 3,3 |
| ]]> MIRANDA LC50a |
0,0423 | 0,0779 |
0,0358 |
|
|
(0,0373-0,0478) | (0,0716-0,0849) | ]]>
(0,0311-0,0406) |
| bb | 3,2 (± 0,32) |
4,7 (± 0,63) | 2,6 (± 0,24) |
| FSc | 1,3 | ]]> 2,2 | 1,1 |
| TACHIRA LC50a | 0,039 |
0,15 | 0,072 |
|
|
(0,031-0,043) | ]]> (0,11-0,17) |
(0,068-0,073) |
| bb | 6,88 (± 0,61) | 7,46 (± 0,73) |
2,36 (± 0,33) |
| FSc | 0,795 | ]]>
0,65 |
0,79 |
| ROCK LC50a | 0,0223 | 0,0269 |
0,0156 |
|
| ]]>
(0,0191-0,0252) | 0,024-0,029) |
( (0,013-0,018) |
| bb | 3,1 (± 0,24) |
4,2 (± 0,36) | 3,2 (± 0,27) |
| ]]> FSc | 0,6 | 0,29 |
0,43 |
a CL50 en mg/L: 95 % IC están entre paréntesis. b Pendiente de la recta probit-log. La desviación estándar (± DE) está entre paréntesis. c Factor de sinergismo (FS): CL50 del insecticida solo/CL50 del insecticida + sinergista Número de larvas evaluadas: 500 por insecticidas.
A la luz de los resultados obtenidos con ambos sinergistas es sugerente que en la resistencia a pirimifos metil intervengan otros mecanismos de acción tan importantes en Ae. aegypti como GST y gen Kdr. El primero se encontró a una alta frecuencia en SANTIAGO DE CUBA (0,8042), TACHIRA (0,525) y ARAGUA (0,45) (tabla 4).
Tabla 4. Valores de frecuencia para el mecanismo de resistencia mediado por esterasas elevadas y glutation-s-transferasa (GST), observadas en Aedes aegypti de 5 localidades
]]>|
|
ROCKEFELLER |
APURE | ARAGUA |
MIRANDA | ]]>
TACHIRA |
SANTIAGO DE CUBA |
| Mecanismos |
(n) | (n) |
(n) | ]]>
(n) |
(n) | (n) |
| Esterasaa | 0 | 1,0 |
0,42 | ]]>
0,831 | 0,771 | 1,0 |
|
|
(350) |
(285) | ]]> (300) |
(350) | (300) | (285) |
| GSTb | 0 |
0,041 | 0,45 | ]]>
0,043 | 0,525 | 0,80 |
|
|
(288) |
(285) | ]]> (300) |
(150) | (300) |
(285) |
a Actividad de esterasas, medida como densidad óptica. b Actividad de glutation-s-transferasa, medida como actividad específica. (n) El total de individuos evaluados está entre paréntesis.
Se demostró que la actividad de las esterasas elevadas desempeñó un papel muy importante en la resistencia a clorpirifos y a temefos en las cepas estudiadas; este mecanismo se encontraba a alta frecuencia génica (tabla 4).
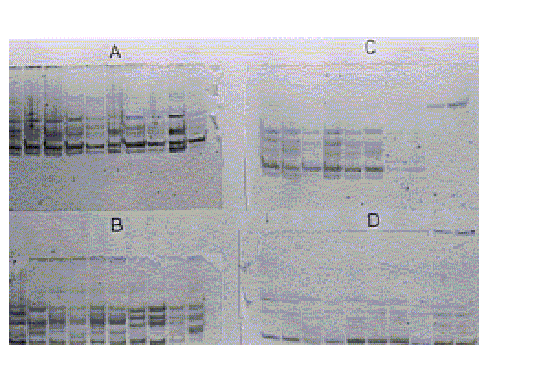
El examen visual de los geles reveló un gran número de bandas de esterasas A y B para estas cepas (fig.1). Por su movilidad electroforética, las bandas fueron enumeradas desde 1 hasta 5, con el correspondiente cálculo de su Rf. Es de destacar que en todas las cepas se observó una banda fuertemente teñida de esterasa, nombrada como esterasa A4 (Esta), con una movilidad relativa de 0,779, que apareció con una alta frecuencia (99 a 100 %), y no se observó en la cepa de ROCKEFELLER (fig. 2). Otras bandas fueron observadas en las 5 cepas pero su intensidad fue menor. El posible papel de esta esterasa en la resistencia a tóxicos organofosforados está aún por demostrar y este aspecto es objeto de investigaciones futuras.
]]>
Gel A: cepa MIRANDA, Gel B: de izquierda a derecha (6 muestras de TACHIRA, 2 de SANTIAGO DE CUBA y 2 de ROCKEFELLER, Gel C: 6 muestras de ARAGUA, 2 de ROCKFELLER y 2 de Culex quinquefasciatus, Gel D: todas las muestras de la cepa APURE.
Fig. 1. Esterasas elevadas en larvas de Aedes aegypti de Venezuela.
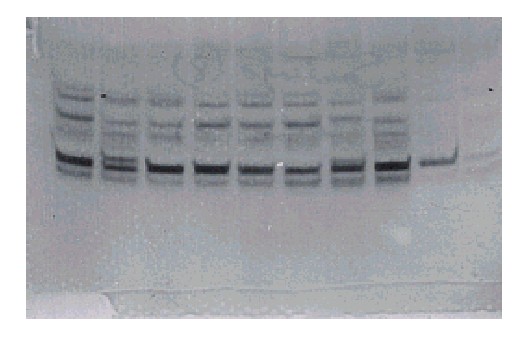
De izquieda a derecha 8 muestras de SANTIAGO DE CUBA y 2 de ROCKEFELLER
Fig. 2. Esterasas elevadas en larvas de Aedes aegypti de Cuba.
Las cepas de Ae. aegypti de Venezuela y Santiago de Cuba mostraron niveles variables de resistencia a los insecticidas organofosforados, temefos, clorpirifos y pirimifos metil. Las cepas de APURE Y SANTIAGO DE CUBA, tratadas en el campo con temefos a 1 ppm, mostraron altos niveles de resistencia a temefos. Sin embargo la selección de una cepa de Ae. aegypti de la India por 20 generaciones con temefos, incrementó la tolerancia a este insecticida solamente de 2 a 3 veces.7
Los altos valores de factor de sinergismo (FS 5) obtenidos con DEF y clorpirifos indicaron que la resistencia a este insecticida estuvo mediada por el mecanismo de esterasas. Altos niveles de resistencia a clorpirifos han sido reportados en poblaciones de Ae. aegypti de Puerto Rico, Santa Lucía y Trinidad.8 Como el clorpirifos no ha sido utilizado para el control de Ae. aegypti en Venezuela, ni en Cuba, es probable que la resistencia detectada a este insecticida sea el resultado de resistencia cruzada de temefos u otros insecticidas organofosforados.9
]]> En el presente estudio, el mecanismo de esterasas elevadas estuvo asociado con la resistencia a temefos, lo que se corroboró por los estudios con sinergistas y las pruebas bioquímicas en la cepa de APURE Y SANTIAGO DE CUBA. Recientemente se reportó el papel importante de la alta actividad de esterasas en una cepa de Aedes aegypti resistente a temefos, que fue seleccionada en el laboratorio por 13 generaciones con este insecticida.10 La asociación del mecanismo de alta actividad de esterasas con la resistencia a temefos y clorpirifos fue reportada en Venezuela9 y Trinidad.11Existen otros mecanismos bien documentados en Ae. aegypti, como son, el gen Kdr basado en la insensibilidad en el sitio de acción y el mecanismo de la elevada actividad de la glutation-s-transferasa (GST, mecanismo de acción metabólica), responsables de la resistencia a DDT.12-14 En los resultados de este trabajo el mecanismo de GST está presente en casi todas las cepas evaluadas, excepto en las cepas de MIRANDA Y APURE. Al parecer, este mecanismo de resistencia no debe ser importante en la resistencia a insecticidas organofosforados en las cepas evaluadas.
En la electroforesis se observó una banda fuertemente teñida, que de acuerdo con su movilidad relativa y su coloración se clasificó como esterasa A4 y cuyo Rf fue de 0,779. Según lo encontrado por Mazzarri (Mazarri MB. Insecticida resistance in two field populations of Aedes aegypti from Venezuela [Tesis de Máster]. University of California, Riverside, California. 1994) en una cepa de Ae. aegypti se mostraba una banda cuya movilidad relativa era de 0,61, que estaba presente en 91 % de los individuos, y fue nombrada esterasa A6. También se clasificó como esterasa A6 la encontrada en adultos de Aedes aegypti procedentes de San Juan de Puerto Rico, con un valor de Rf de 1,00.15 Producto de la nomenclatura de estas esterasas se dificulta la comparación entre ellas.
Es evidente que Ae. aegypti ha desarrollado resistencia a organofosforados, lo que puede resultar en un problema potencial para el control de esta especie. Agentes de control biológico, como el Bacillus thuringiensis var. israelensis pudieran ser seriamente considerados para el control larval en los programas de control de Ae. aegypti. Se hace necesario continuar el monitoreo de la susceptibilidad a insecticidas en las poblaciones de Ae. aegypti para brindar información necesaria que ayude a la toma de decisiones sobre el uso correcto de insecticidas. Lograr programas de control sostenidos, con énfasis en la reducción de focos y el saneamiento ambiental, con el objetivo de utilizar menos insecticidas para disminuir la presión de selección sobre las poblaciones resistentes.
Five strains of Aedes aegypti L. one from Cuba and 4 from Venezuela were evaluated to determine their resistance to organophosphate insecticides (temephos, chlorpiriphos and pirimiphos methyl). In the Venezuelan strains only APURE showed resistance to temephos. In TACHIRA and MIRANDA moderate resistance values were noted (FR50 5 to 10x) for chlorpiriphos and high levels of resistance (FR 10x) to this insecticide were found in ARAGUA. All the Venezuelan strains showed high levels of resistance to pirimiphos methyl. The Cuban strain from Santiago de Cuba revealed moderate resistance to temephos and pirimiphos methyl, but high resistance to chlorpiriphos. The results of the biochemical tests proved the presence of esterase and glutathione-s-transferase at high frequency in almost every strain. By the polyacrilamide gel electrophoresis, a strongly stained band was observed in all the strains with a Rf value of 0.779; it was named esterase A4 and was not seen in the susceptible reference strain. The meaning of this esterase in the resistance to organophosphate insecticides is yet to be determined. Resistance to these insecticides in Aedes aegypti is a serious problem for the control of this species therefore integrated management strategies were proposed to prevent or delay the appearance of this species in Cuba and Venezuela.
Subject headings: AEDES/enzimology; ESTERASES//analysis; INSECTICIDES.
Recibido: 29 de septiembre del 2000. Aprobado: 6 de noviembre del 2000.
Lic. Juan A. Bisset. Instituto de Medicina Tropical "Pedro Kourí". Apartado 601, Marianao 13, Ciudad de La Habana, Cuba.Correo electrónico: ciipk@ipk.sld.cu