{kind=link}
ARTÍCULO ORIGINAL
Propiedades biológicas de cepas de virus dengue serotipo 2 aisladas durante la epidemia en Santiago de Cuba, 1997
Biological properties of dengue virus serotype 2 strains isolated from the epidemic occurred in Santiago de Cuba in 1997
]]>
Dra. C. Rosmari Rodríguez-Roche,I MSc. Liudmila López Matilla,II Dra. C. Mayling Alvarez Vera,I MSc. Luis Morier Díaz,I Dra. C. María G. Guzmán TiradoI
I Instituto de Medicina Tropical "Pedro Kourí". La Habana, Cuba.
II Centro de Inmunología Molecular. La Habana, Cuba.
Introducción: durante la epidemia cubana de dengue ocurrida en Santiago de Cuba, 1997, se evidenció un incremento de la severidad en el tiempo en términos de proporción de casos graves y muertes por dengue hemorrágico, que pudiera estar dado por la aparición de mutantes de escape a la neutralización, con mayor potencial virulento.
Objetivos: estudiar algunas propiedades biológicas de cepas aisladas en diferentes momentos de la epidemia de Santiago de Cuba en 1997.
Métodos: se estudiaron 9 cepas de DENV-2, se evaluó el efecto citopatogénico y el crecimiento viral en las líneas celulares C6/36 HT y Vero, tamaño de las placas virales, sensibilidad a la temperatura, neurovirulencia en ratones lactantes y la influencia del pH en la unión del virus a la célula, así como en el medio de multiplicación. ]]>
Resultados: las cepas del final de la epidemia mostraron propiedades que las diferencian de las aisladas al inicio, entre las que se encuentran un mayor efecto citopatogénico en células C6/36 HT, títulos virales superiores y una mayor neurovirulencia en ratones lactantes. Por otra parte, en la unión del virus a la célula las cepas del inicio de la epidemia resultaron favorecidas por un pH ácido mientras que a las cepas del final de la epidemia las favoreció un pH básico.
Conclusiones: pudo demostrarse que además de los cambios genotípicos observados previamente, existen diferencias fenotípicas entre cepas de distintos momentos de la epidemia, que pudieran estar asociados con diferencias en cuanto a la adecuación viral de estas y(o) en su potencial virulento.
Palabras clave: dengue, cepas, propiedades biológicas, severidad.
Introduction: during the Cuban epidemic that occurred in Santiago de Cuba in 1997, there was observed increasing severity in the course of time, in terms of proportion of serious dengue haemorrhagic cases and deaths that could be due to the emergence of escape mutants to neutralization with greater virulent potential.
Objective: to study some biological attributes of a group of strains isolated at different points of time during the Santiago de Cuba epidemic in 1997.
Methods: nine DENV-2 strains were studied. The cytopathogenic effect, the viral growth in C6/36 HT and VERO cell lines, the virus plaque sizes, the sensitivity to temperatures, the neurovirulence in newborn mice and the influence of the pH in the union of the virus to the cell as well as in the multiplication medium were all evaluated.
Results: the strains isolated at the end of the epidemic differed from those of the beginning showing increased neurovirulence in newborn mice and higher viral titers and greater cytopathogenic effect in HT C6/36 cells. On the other hand, the virus and the cell union was favored by acid pH when testing strains from the beginning of the epidemic, whereas this union was favored by the basic PH in the strains isolated at the end of the epidemic
Conclusions: the present study managed to show that in addition to the previously observed genotypical changes, there were phenotypical differences among the strains isolated at different points of time in the epidemic; all these aspects may be associated with differences in the viral fitness and/or in the virulent potential of these strains.
]]>
Key words: dengue, strains, biological attributes, severity.
INTRODUCCIÓN
La fiebre del dengue (FD) y la fiebre hemorrágica del dengue/síndrome de choque por dengue (FHD/SCD) se han convertido en un problema creciente de salud en el mundo tropical y subtropical.1 Específicamente en Cuba, han ocurrido epidemias bien documentadas que han facilitado el estudio de esta entidad viral. En 1977, ocurrió una epidemia de FD causada por el virus dengue serotipo 1 (DENV-1), genotipo americano, que afectó a todo el país con más de 500 000 casos reportados;2 4 años más tarde sucede una epidemia de FHD causada por el virus dengue serotipo 2 (DENV-2), relacionado genéticamente con las cepas viejas del genotipo asiático.3 La introducción de este genotipo en Cuba marcó la aparición de la forma más severa de la enfermedad en las Américas, desarrollándose una epidemia sin precedentes en la región. Se reportó un total de 344 203 casos, 10 312 severos con 158 fallecidos, 101 de ellos niños.4
Desde 1982 hasta 1996, se desarrolló una efectiva campaña para el control del Aedes aegypti el principal vector de esta enfermedad, lo cual permitió que no se observaran casos de dengue. Sin embargo, en 1997, el mosquito había reinfestado varias provincias del país, consecuentemente el DENV-2, de nuevo de origen asiático, reemergió en el municipio de Santiago de Cuba. Se diagnosticaron 3 012 casos confirmados serológicamente, de los cuales 205 se clasificaron como casos de FHD y 12 de estos fallecieron.5 En 98 % de los casos graves se comprobó la infección secundaria (DENV-1/DENV-2). Un análisis comparativo de las epidemias de DENV-2 de 1981 y 1997 muestra que la de 1997 resultó más severa, reporte de gran implicación epidemiológica si se tiene en cuenta que ocurrió a 20 años de la infección primaria con DENV-1.6
Las epidemias de 1981 y 1997 tuvieron en común, la asociación de la FHD/SCD con la infección secundaria, la presencia de cepas de origen asiático como agente causal de la infección secundaria y el incremento de la severidad con el transcurso de la epidemia. Teniendo en cuenta estas observaciones en el año 2000, Guzmán y otros, plantearon como hipótesis que ese incremento en severidad podría estar dado por la aparición de mutantes de escape a la neutralización heterotípica.7 La epidemia de 1997 es la más severa que se ha registrado en Cuba hasta la fecha y el Laboratorio de Arbovirus del Instituto de Medicina Tropical "Pedro Kourí" (IPK) cuenta con un banco de cepas de bajo pase en cultivo celular, obtenidas durante el transcurso de esta epidemia. Por lo tanto, se consideró de gran interés realizar una caracterización biológica de cepas aisladas en diferentes momentos de la epidemia, de las cuales se ha realizado previamente la caracterización molecular,8 para conocer si los cambios genotípicos encontrados tienen implicaciones fenotípicas, que pudieran relacionarse con el incremento de severidad observado con el transcurso de la epidemia en el tiempo. ]]>
MÉTODOS
Cepas virales
Para el estudio se utilizaron 9 cepas de DENV-2 aisladas durante la epidemia de Santiago de Cuba, 1997, seleccionándose cepas aisladas en diferentes momentos de la epidemia. Todos los aislamientos virales se obtuvieron a partir de suero de pacientes en fase aguda, con excepción de la cepa 205/97 aislada a partir de muestra de tejido (bazo) obtenida durante la autopsia de un caso fatal. Las cepas utilizadas fueron multiplicadas en la línea celular C6/36 HT (Aedes albopictus, cepa celular del clono C6/36, ATCC CRL 1660). Una vez realizada la cosecha viral se prepararon lotes de todas las cepas, los cuales fueron conservados a - 70 °C y se les determinó el título viral. En la tabla 1 se brinda información sobre las cepas estudiadas.
Estudio del ECP en las líneas celulares C6/36HT y Vero
]]>
Se emplearon tubos plásticos sembrados con células C6/36HT y Vero (riñón de mono verde africano, ATCC CCL-81) que presentaban una monocapa confluente. Se inocularon con las cepas en estudio a una multiplicidad de infección (m) de 0,01. Luego de 1 h de adsorción a 33 °C en el caso de las células C6/36 HT y a 37 °C para las células Vero se eliminó el inóculo, se lavó la monocapa con el medio correspondiente (medio esencial mínimo eagle [MEM] para los cultivos de C6/36 HT y medio 199 para los de Vero) y finalmente se les adicionó 1 mL de medio de mantenimiento que contenía los medios indicados antes y suero de ternera fetal inactivado (STFI) a 2 %. Los tubos de C6/36 HT se incubaron durante 6 d a 33 °C; tiempo durante el cual se observó diariamente el efecto citopatogénico (ECP). En el caso de los tubos sembrados con la línea celular Vero el período de incubación fue de 9 d a 37 °C.
Cinética de crecimiento viral en las líneas celulares C6/36HT y Vero
Se emplearon tubos plásticos sembrados con células C6/36 HT y Vero con monocapa celular confluente. Se inocularon con las cepas en estudio a m= 0,01. Luego de 1 h de adsorción a 33 °C en el caso de C6/36 HT y a 37 °C para Vero se eliminó el inóculo, se lavó la monocapa con 1 mL del medio correspondiente y por último se les adicionó 1 mL de medio de mantenimiento. A diario se tomó un tubo de cada una de las cepas inoculadas, del primero al sexto día posinoculación en la línea celular C6/36 HT y del primero al noveno día posinoculación en la línea celular Vero; el sobrenadante se recogió y se conservó a - 70 ºC, después fue descongelado para ser titulado por triplicado según el método descrito antes, a partir de diluciones seriadas en base 10 (10-1-10-4). Los resultados obtenidos se graficaron enfrentando la media de los logaritmos de los títulos virales (log 10 UFP/mL) contra los días de cosecha con la representación además de la desviación estándar.
Titulación viral
Para la titulación viral se siguió el método descrito por Álvarez y otros utilizando células BHK-21 (riñón de hámster recién nacido, clono 13, ATCC CCL 10).9
]]>
Sensibilidad a la temperatura y tamaño de placas
Los títulos virales se establecieron a 37 y 39 °C, determinándose el porcentaje de reducción del título viral con el aumento de la temperatura para cada cepa viral.9 Las placas virales fueron evaluadas según su tamaño. Se consideraron pequeñas todas aquellas con un diámetro de 1 mm y grandes aquellas con un diámetro > 1 mm. También se tuvo en cuenta la forma de las placas obtenidas, se clasificaron en redondeadas, estrelladas o heterogéneas en el caso que una misma cepa viral tuviera placas redondeadas y estrelladas.
Determinación de la neurovirulencia en ratones lactantes
Las cepas en estudio fueron diluidas en medio 199 e inoculadas por vía intracerebral en ratones lactantes de 24 h de nacidos, a razón de 20 UFP/animal se inocularon 2 familias de 8 a 10 animales cada una, para cada cepa. Los ratones se observaron a diario durante 14 d en busca de cualquier signo de enfermedad o muerte. Las muertes ocurridas en las primeras 24 a 48 h, no fueron consideradas a los efectos del estudio. Se determinó el porcentaje de supervivencia en los días posinfección para cada cepa viral.
Influencia del pH en la multiplicación del virus
]]>
Se estudiaron todas las cepas escogidas mediante diluciones seriadas en base 10 (10-1-10-4); se aplicó el método descrito para la titulación.9 Los títulos virales se realizaron por cuadruplicado para su determinación a pH 6,5; 7; 7,5 y 8; se ajustó el pH del medio con bicarbonato de sodio.
Influencia del pH del medio de inoculación en la unión del virus a la célula
Tubos plásticos sembrados con células C6/36 HT con monocapa celular confluente se inocularon con las cepas en estudio a m= 0,01. Al medio utilizado en la dilución de las cepas se le ajustó el pH con bicarbonato de sodio a pH 6; 6,5; 7 y 7,5. Luego de 1 h de adsorción a 33 °C se eliminó el inóculo, se lavó la monocapa con 1 mL de medio de mantenimiento sin SFBI y finalmente se les adicionó 1 mL de medio de mantenimiento. Los tubos se incubaron durante 2 d a 33 °C. Se tomó un tubo de cada una de las cepas inoculadas a las 24 y las 48 h; se recogió el sobrenadante y se conservó a - 70 ºC. Posteriormente, este sobrenadante fue descongelado para ser titulado por triplicado según el método descrito (259),9 realizando diluciones seriadas en base 10 (10-1-10-4).
Análisis estadístico
Los resultados obtenidos en los experimentos para determinar la influencia del pH en la multiplicación de los DENV así como en la unión de este a la célula fueron analizados mediante el programa STATISTICA, en el primer caso con un ANOVA/MANOVA mientras que en el segundo caso se hizo una prueba t de Student.
]]>
RESULTADOS
ECP en células C6/36 HT
El ECP observado consistió en la formación de sincitios redondeados, con bordes bien definidos y alta refringencia, que degeneraban hacia la desaparición de la estructura de la monocapa. Todas las cepas estudiadas fueron productoras de ECP. A pesar de que el ECP descrito anteriormente fue similar para todas las cepas evaluadas, se pudo observar diferencia en cuanto al momento de aparición y a su intensidad. Entre las cepas de DENV-2, la 13/97 aislada el primer mes de la epidemia a pesar de que su ECP tuvo un comienzo tardío a partir del tercer día posinoculación, ya en el quinto día se observó la formación de sincitios en 75 % de la monocapa celular. En el resto de las cepas la aparición del ECP comenzó a partir del segundo día posinoculación. No obstante a esta similitud en cuanto al día de aparición, se comprobó una mayor intensidad del ECP, en las cepas del final de la epidemia. La cepa 205/97 productora de FHD, aislada durante el séptimo mes de la epidemia, tuvo su máximo ECP desde el quinto día posinoculación, manifestando mayor intensidad que el resto de las cepas.
ECP en células Vero
El ECP fue similar para todas las cepas estudiadas. Este consistió en redondeamiento celular, que conllevó al desprendimiento y la destrucción de la monocapa. Se observa una aparición tardía del ECP, encontrándose un grupo de cepas (inicio y medio) en las que se detecta entre el séptimo y octavo día posinoculación. En el resto de las cepas correspondientes al final de la epidemia, no es hasta el noveno día posinoculación en que es apreciado el ECP. En cuanto a su intensidad, no se encontraron diferencias entre las cepas estudiadas. ]]>
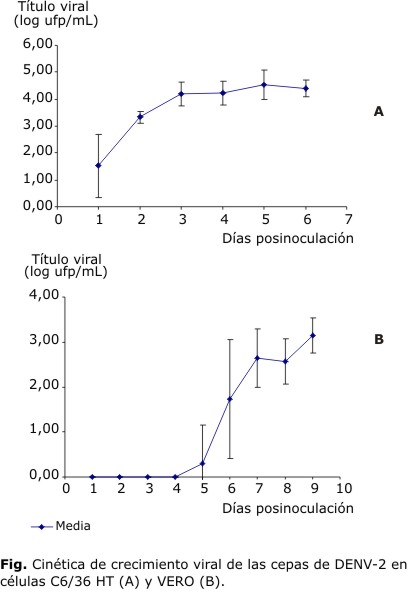
Cinética de crecimiento viral en células C6/36 HT
En la figura se muestra la curva obtenida a partir de la media y la desviación estándar del título infectivo de las cepas de DENV-2 estudiadas por día de cosecha. Para la cepa 205/97 no se detectó título viral a las 24 h de la inoculación como en el resto de las cepas y tuvo una caída brusca al quinto día posinoculación.
Cinética de crecimiento viral en células Vero
Los títulos infectivos de las cepas de DENV-2 en esta línea celular fueron menores que los observados en la cinética de crecimiento viral en C6/36 HT. Los títulos comienzan a elevarse entre el quinto y el sexto día de cosecha. En los días posteriores de la cinética se observa una curva que tiene su máximo en el séptimo día para la mayoría de las cepas, a continuación se ve una caída en el octavo día y una tendencia al aumento en el noveno. Aunque la manera en que aumenta y disminuye el título es similar para todas las cepas y se obtienen curvas semejantes en cuanto a su forma, la marcada diferencia de los títulos obtenidos en los días de cosecha para cada una de las cepas es notable. Por esta razón las medias de los títulos virales muestran una desviación estándar apreciable. Este resultado puede apreciarse en la figura.
]]>
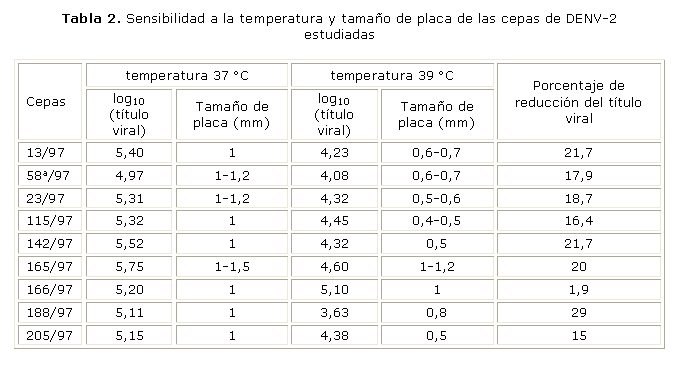
Sensibilidad a la temperatura
De forma general en todas las cepas de DENV-2 el título viral se redujo de un 15-29 % al aumentar la temperatura a 39 °C, con excepción de la cepa 166/97 para la cual el título se redujo solo en 1,9 %. Coincidentemente para esta cepa, no hubo una reducción del tamaño de las placas. Los datos obtenidos de este experimento indican la capacidad de esta cepa (aislada de un paciente con FHD) de multiplicarse a temperaturas más elevadas. En la mayoría de las cepas la reducción del título viral coincidió con una reducción del tamaño de las placas. En la tabla 2 se muestra cómo el aumento de la temperatura afectó el título viral de las cepas en estudio. Las placas observadas para las cepas de DENV-2 fueron en su mayoría heterogéneas.
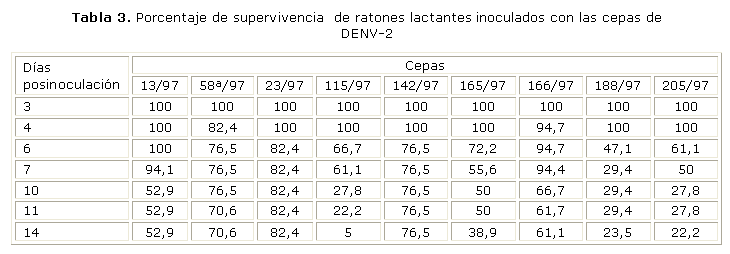
Neurovirulencia en ratones lactantes
La neurovirulencia en ratones se evaluó sobre la base del porcentaje de supervivencia. Los resultados obtenidos se muestran en la tabla 3. La mayoría de las cepas provocaron las primeras muertes a partir del sexto día posinoculación, con excepción de la cepa 13/97 del inicio de la epidemia, que provocó las primeras muertes a partir del séptimo día. Las familias de ratones lactantes inoculadas con las cepas del inicio (13/97, 58a/97 y 23/97), mostraron los mayores porcentajes de supervivencia que oscilan entre 53 y 82 %. Las familias con menor porcentaje de supervivencia, se correponden con las inoculadas con cepas del final de la epidemia: 165/97, 188/97 y 205/97; esta supervivencia osciló entre 22 y 39 %, lo cual indica que este grupo contiene las cepas más neurovirulentas. Es de destacar que la cepa 166/97 de este grupo final constituye una excepción, con un alto porcentaje de supervivencia de las familias inoculadas. Por su parte la cepa 115/97 a pesar de no ser aislada de un caso grave, resultó la más neurovirulenta.
Influencia del pH del medio en la multiplicación del virus
]]>
Para evaluar la influencia del pH en la multiplicación viral, se determinó el título viral de cada una de las cepas crecidas a pH del medio overlay desde 6,5 a 8. Los títulos infectivos de las cepas de DENV-2, no presentaron diferencias con la variación del pH (p= 0,442) aunque tienden a ser mejores a pH 8. Es de destacar que las cepas que alcanzan los mejores títulos virales en todos los pH evaluados (165/97 y 205/97) fueron aisladas de pacientes con FHD.
Influencia del pH del medio de inoculación en la unión del virus a la célula
La influencia del pH en la unión del virus a la célula se medió mediante la titulación viral de los cultivos inoculados, 24 y 48 h posinoculación. Aunque el experimento se realizó ajustando el pH del inóculo a los valores: 6,0; 6,5; 7,0 y 7,5; solo en los valores extremos de pH, ácido (6,0) y básico (7,5) se encontraron diferencias, observándose 2 patrones de comportamiento. Existe un grupo de cepas en las que el pH 6,0 favorece la unión, por cuanto el título viral es detectable a las 24 h posinoculación y otro grupo en el que este pH no es permisivo y se corresponde con las cepas del final de la epidemia. En este segundo grupo, es posible detectar título viral con el pH 7,5. A las 48 h posinoculación se observan diferencias significativas en el título infectivo al variar el pH (p= 0,032). Para todas las cepas los títulos son ligeramente más altos a pH 7,5.
DISCUSIÓN
La circulación de los 4 serotipos del DENV en el mundo se ha incrementado bruscamente en los últimos años, lo que se ha acompañado por un incremento en la diversidad genética de este virus. El análisis filogenético realizado por Zanotto y otros,10 sobre la base de los aislamientos de dengue acumulados en un período de 200 años, reveló un incremento simultáneo en el número de linajes virales en todos los serotipos. Evidentemente, este incremento de la diversidad genética, podría tener también implicaciones fenotípicas, como son: la emergencia de virus antigénicamente diferentes, más virulentos y(o) con cambios en cuanto a su tropismo celular. ]]>
Durante la epidemia cubana de dengue ocurrida en Santiago de Cuba, 1997, se pudo constatar un incremento en la severidad de la epidemia, dado por un aumento cada mes de proporción de casos de FHD/SCD así como la proporción de muertos.7 Se planteó como hipótesis, que este incremento podría estar dado por la aparición de mutantes de escape a la neutralización, capaces de producir las formas más severas de la enfermedad.7 Basados en estas observaciones, Rodríguez-Roche y otros, realizaron la caracterización molecular de cepas aisladas en diferentes momentos de la epidemia, mediante la secuenciación nucleotídica del genoma viral completo. Los resultados obtenidos por estos autores mostraron que existía un claro patrón de evolución, porque los aislamientos del inicio de la epidemia se diferenciaban de los aislados 4 meses más tarde.8Los resultados obtenidos en el estudio de las propiedades biológicas de estas cepas, indican que existe una variabilidad en el comportamiento de los virus aislados en diferentes momentos. Pudo encontrarse que las cepas de DENV-2 son más citopatógénicas en células C6/36 HT que en células Vero. Los DENVs infectan las células diana mediante la unión a varios receptores celulares, muchos de los cuales son aún desconocidos. Algunos reportes como el de Bielefeldt-Ohmann y otros11 muestran que al infectar diferentes tipos de células humanas, el nivel de virus que se une a las células varía ampliamente al comparar diferentes líneas celulares y de modo notable entre cepas virales incluso del mismo serotipo. Existen reportes en la literatura que demuestran una mayor sensibilidad de la línea C6/36 HT, para la multiplicación de los DENVs.12 A este aspecto podría agregarse el hecho de que las cepas del estudio fueron aisladas y crecidas mediante 2 a 3 pases sucesivos en esta línea celular, lo que sin dudas influye de manera notable en su adaptación en las células C6/36 HT, haciendo la línea celular Vero menos permisiva para su multiplicación.
Por otra parte, se encontró una concordancia entre los experimentos de estudio del ECP y la cinética de crecimiento viral. De esta forma las cepas menos citopatogénicas (inicio), mostraron un crecimiento viral más tardío. En el caso de las cepas del final caracterizadas por producir un mayor ECP en células C6/36 HT, se observó una caída de los títulos virales hacia el final de la cinética de crecimiento, lo que se explica por el deterioro del cultivo celular.
En este estudio se han encontrado algunos patrones de comportamiento que dividen las cepas de inicio y final en 2 grupos, como es la tendencia de las cepas del final de las epidemias a ser más citopatogénicas y que el pH ácido o básico favorezca la unión a la célula de un grupo de cepas y otras no, que pudieran tener influencia sobre la patogenia viral. El título infectivo en todas las cepas aumentó con los días de cosecha, pero la cepa 205/97 tuvo una caída brusca al quinto día posinoculación. Eso pudo deberse a que esta cepa al cuarto día de observación presentaba un ECP mucho más intenso que el resto de las cepas, lo cual indica su mayor potencial citopatogénico. Es lógico pensar que con la total destrucción de la monocapa, haya una disminución notable del título infectivo. Este resultado es de especial importancia si se tiene en cuenta que este aislamiento pertenece a 1 caso fatal y que se obtuvo al final de la epidemia. El estudio de la secuencia nucleotídica del genoma completo de la cepa 205/97 determinada recientemente, muestra que la cepa 205/97 acumuló un número considerable de cambios en el gen que codifica la polimerasa del virus. Dentro de los cambios nucleotídicos observados en este gen, uno de ellos se corresponde con una sustitución aminoacídica.8
Aunque no se conocen las implicaciones de estos cambios, podrían estar relacionados con el comportamiento encontrado para esta cepa en cuanto a sus propiedades biológicas. En 1999, Bartholomeusz y otros, encontraron que algunas mutaciones introducidas en el gen de la polimerasa viral del virus de la hepatitis C expresada en baculovirus, podría conllevar a una reducción total de la actividad enzimática, sin embargo, un simple cambio aminoacídico conservado, introducido en otros motivos, podía ocasionar un incremento de alrededor de 50 % en su actividad enzimática.13
Cuando se analizó la sensibilidad a la temperatura de las diferentes cepas se pudo observar que todas fueron afectadas por un aumento de la temperatura. Sin embargo, la cepa 166/97 del DENV-2 aislada de un paciente con FHD, muestra mayor capacidad para crecer a temperaturas más elevadas con una disminución despreciable del título viral y la conservación del tamaño de la placa viral. A pesar de que otras cepas productoras de FHD evaluadas en este estudio, no muestran una alta resistencia a la temperatura, muchos autores consideran esta característica fenotípica como marcador de virulencia. De esta forma las cepas vacunales atenuadas, obtenidas para algunos Flavivirus por pases sucesivos en cultivo de tejidos, muestran una mayor sensibilidad a la temperatura y placas de menor tamaño que el obtenido para las cepas salvajes que le dieron origen.14,15
Otro de los marcadores fenotípicos que indica una mayor virulencia de las cepas, es la neurovirulencia en ratones lactantes. En este sentido, en el presente estudio pudo apreciarse, que en las cepas del final de la epidemia se encuentran los menores porcentajes de supervivencia, lo cual pudiera asociarse con su mayor virulencia. Es significativo que la cepa 115/97 del DENV-2 que no fue aislada de un caso grave muestre el menor porcentaje de supervivencia. De acuerdo con los resultados de la secuenciación nucleotídica, pueden encontrarse 3 cambios nucleotídicos únicos para esta cepa cuando se comparan con el resto de las cepas de DENV-2 secuenciadas.8 Esos cambios se encuentran en la zona NS2B-NS3, aunque no se puede plantear que los cambios observados en la secuencia están relacionados con una mayor neurovirulencia de esta cepa. Shafee y otros reportaron que la serina-proteasa NS3 y su proteína precursora N2B-NS3 del DENV-2, inducen apoptosis en células Vero y se observó un alto porcentaje de apoptosis en células que expresan la proteína precursora. Además, demostraron que algunas mutaciones en la NS3 reducen la capacidad de esta de inducir el fenómeno apoptótico.16
En la literatura consultada, no existen reportes acerca de la influencia del pH en la multiplicación y la unión a la célula de los DENV-2. Un estudio realizado con la cepa A15 del DENV-2 muestra su capacidad de multiplicación, al variar el pH en un rango de 6,5 a 8, que alcanza el mejor título viral a pH 7,5; mientras que la cepa 116/00 del DENV-3 solo produjo placas a pH 7,5 y 8 con el título más alto a pH 7,5.9 De forma similar, en el presente estudio las cepas de DENV-2 presentan título viral en cada valor de pH pero los mejores títulos son obtenidos a pH 8.
El conocimiento acerca de las sustituciones nucleotídicas en el DENV no solo brinda información acerca de su historia epidemiológica, también es crucial para el entendimiento de su proceso de evolución viral. La identificación de cambios genéticos que se puedan relacionar con la expresión de alguna propiedad biológica resulta de gran importancia en la caracterización completa de cepas de dengue (biológica, antigénica y molecular).
En este estudio se han encontrado algunos hallazgos que hablan a favor de cambios en las propiedades biológicas de las cepas con la progresión de las epidemias. Algunos de estos cambios fenotípicos podrían relacionarse con cambios genotípicos. Con el avance de la biología molecular, tenemos a nuestra disposición nuevas tecnologías que permitirán definir el impacto de los cambios genotípicos observados en la evolución de las epidemias y la influencia que tienen en el comportamiento biológico del virus. ]]>
La evolución del virus del dengue ha tenido un gran impacto en la virulencia y en la epidemiología de la enfermedad en todo el mundo. Se han evidenciado diferencias genéticas y antigénicas entre cepas virales de diferente origen geográfico, la falta de modelos animales para reproducir la enfermedad ha hecho difícil demostrar los determinantes genéticos relacionados con la mayor virulencia de algunos serotipos y(o) genotipos particulares.17Leitmeyer y otros, mediante la secuenciación nucleotídica del genoma completo del DENV-2 a partir del plasma de pacientes infectados, tanto con FHD como FD, de diferentes orígenes geográficos, encontraron cambios estructurales que podrían ser determinantes de FHD.18 Recientemente, Cologna y otros, generaron una serie de clonos infecciosos quiméricos de DENV-2, con diferentes combinaciones de las secuencias de los genotipos asiático (productor de FHD/SCD) y americano (productor de FD) que incluyen los cambios encontrados por Leitmeyer y otros.19 Los resultados encontrados por estos autores revelan que para algunos de los virus quiméricos se alteró la morfología de las placas, y que a pesar de que todos eran capaces de replicarse de forma similar en células de mosquito, se encontraron diferencias en cuanto a la salida de ARN viral por célula infectada, cuando los experimentos se hicieron en macrófagos humanos y células dendríticas. Los datos sugieren que los cambios encontrados podrían ser responsables de una disminución de la replicación viral en células blanco humanas o de una mayor virulencia durante la infección.
La variabilidad del virus circulante dentro de una misma epidemia, enmarcada en una región en particular y en un período de tiempo dado, ha sido poco abordada como estudio. Sin dudas, para dar continuidad a este trabajo, deben evaluarse otros modelos que permitan demostrar el papel de los cambios encontrados en la patogénesis viral.
REFERENCIAS BIBLIOGRÁFICAS
1. Gubler DJ. Dengue/dengue haemorrhagic fever: history and current status. Novartis Found Symp. 2006;277:3-16, 16-22, 71-3, 251-3. ]]>
2. Mas P. Dengue fever in Cuba in 1977: some laboratory aspects. PAHO Scientific Publication. 1979;375:40-43.
3. Guzmán MG, Deubel V, Pelegrino JL, Rosario D, Marrero M, Sariol C, et al. Partial nucleotide and amino acid sequences of the envelope and the envelope/nonstructural protein-1 gene junction of four dengue-2 virus strains isolated during the 1981 Cuban epidemic. Am J Trop Med Hyg. 1995;52(3):241-6.
4. Kourí G, Guzmán MG, Bravo J. Hemorrhagic dengue in Cuba: history of an epidemic. Bull Pan Am Health Organ. 1986;20(1):24-30.
5. Kourí G, Guzmán MG, Valdés L, Carbonel I, del Rosario D, Vázquez S, et al. Reemergence of dengue in Cuba: a 1997 epidemic in Santiago de Cuba. Emerg Infect Dis. 1998;4(1):89-92.
6. Guzmán MG, Kourí G, Valdés L, Bravo J, Vázquez S, Halstead SB. Enhanced severity of secondary dengue-2 infections: death rates in 1981 and 1997 Cuban outbreaks. Rev Panam Salud Publica. 2002;11(4):223-7. ]]>
7. Guzmán MG, Kourí G, Halstead SB. Do escape mutants explain rapid increases in dengue case-fatality rates within epidemics? Lancet. 2000;355(9218):1902-3.
8. Rodriguez-Roche R, Alvarez M, Gritsun T, Halstead S, Kourí G, Gould EA, et al. Virus evolution during a severe dengue epidemic in Cuba, 1997. Virology. 2005;334(2):154-9.
9. Alvarez M, Rodriguez-Roche R, Bernardo L, Morier L, Guzmán G. Improved Dengue Virus Plaque Formation on BHK21 and LLCMK2 Cells: Evaluation of Some Factors. Dengue Bulletin. 2005;29.
10. Zanotto PM, Gould EA, Gao GF, Harvey PH, Holmes EC. Population dynamics of flaviviruses revealed by molecular phylogenies. Proceedings of National Academy of Sciences. 1996;93(2):548-53.
11. Bielefeldt-Ohmann H. Analysis of antibody-independent binding of dengue viruses and dengue virus envelope protein to human myelomonocytic cells and B lymphocytes. Virus Res. 1998;57(1):63-79. ]]>
12. Vorndam AV, Kuno G. Laboratory diagnosis of dengue virus infections. In: Gubler D, Kuno G, editors. Dengue and Dengue Hemorrhagic Fever. Wallingford: CAB International; 1997. p. 313-34.
13. Bartholomeusz A, Thompson P. Flaviviridae polymerase and RNA replication. J Viral Hepat. 1999;6(4):261-70.
14. Alvarez M, Guzmán MG, Pupo M, Morier L, Bravo J, Rodriguez R. Study of biologic attributes of Cuban dengue 2 virus after serial passage in primary dog kidney cells. Int J Infect Dis. 2001;5(1):35-9.
15. Barrett AD, Monath TP, Cropp CB, Adkins JA, Ledger TN, Gould EA, et al. Attenuation of wild-type yellow fever virus by passage in HeLa cells. J Gen Virol. 1990;71(Pt 10):2301-6.
16. Shafee N, AbuBakar S. Dengue virus type 2 NS3 protease and NS2B-NS3 protease precursor induce apoptosis. J Gen Virol. 2003;84(Pt 8):2191-5. ]]>
17. Rico-Hesse R. Microevolution and virulence of dengue viruses. Adv Virus Res. 2003;59:315-41.
18. Leitmeyer KC, Vaughn DW, Watts DM, Salas R, Villalobos I, de C, et al. Dengue virus structural differences that correlate with pathogenesis. J Virol. 1999;73(6):4738-47.
19. Cologna R, Rico-Hesse R. American genotype structures decrease dengue virus output from human monocytes and dendritic cells. J Virol. 2003;77(7):3929-38.
]]>
Recibido: 30 de junio de 2011.
Aprobado: 8 de julio de 2011.
Rosmari Rodríguez-Roche. Instituto de Medicina Tropical "Pedro Kourí". Autopista Novia del Mediodía, Km 6 ½. La Habana, Cuba. AP 601 Marianao 13. CP 17100. Fax: 53-7-2046051. Correos electrónicos: rosmari@ipk.sld.cu; liudmila@cim.sld.cu; mayling@ipk.sld.cu; morier@ipk.sld.cu; lupe@ipk.sld.cu ]]>
{kind=link}
{kind=link}