ARTÍCULO DE REVISIÓN
Bases moleculares e importancia clínica de los excepcionales fenotipos de grupo sanguíneo P y PK
Molecular bases and clinical importance of the exceptional phenotypes of blood group P and PK ]]>
Dr. Daniel de La Vega Elena I; Dra. Asa Hellberg II; Dr. Mario Pivetta I; Dr. Miguel Ángel Raillon I; Dr. Sergio Chialina I; Prof. Dr. E. Solis I; Prof. Dr. Martín Olsson II
I Laboratorio de Inmunohematología. Servicio de Medicina Transfusional. Hospital Italiano Garibaldi. Rosario, Argentina.
II Centro de Sangre, Hospital Universitario. Lund, Suecia.
]]>
RESUMEN
Todas las células de los individuos con fenotipos "p" y "Pk" carecen de los antígenos glucídicos de grupo sanguíneo "públicos". Estos individuos forman anticuerpos "naturales" y "regulares" contra las estructuras faltantes (anti-P, anti-PP1 y anti-PP1Pk), que aglutinan y/o hemolisan los glóbulos rojos de todos los individuos, excepto aquellos con el mismo fenotipo. Por lo excepcional de estos cuadros (unos pocos individuos por millón) y su asociación con reacciones transfusionales hemolíticas graves y aborto espontáneo a repetición, estos individuos deben ser identificados y caracterizados.
Palabras clave: grupo sanguíneo, aloinmunización, eritrocitario, glicosiltransferasas, transfusión, gestación.
ABSTRACT ]]>
All the cells of the individuals with phenotypes "p" y "Pk" lack the glucidic antigens of "public" blood groups. These subjects form natural and regular antibodies against the missing structures (anti-P, anti-PP1 y anti-PP1Pk) that agglutínate or hemolyze the red-blood cells of every individual, excepting those with the same phenotype. Due to the exceptional character of these pictures (a few individuals per million) and their association with severe transfusional hemolytic reactions and repeated miscarriages, these subjects should be identified and characterized.
Fenotipos de grupo sanguíneo p y Pk
]]>
Los antígenos de grupo sanguíneo pueden definirse como "un carácter hereditario presente en la cara externa de la membrana plasmática del glóbulo rojo, definido por un aloanticuerpo especifico".Los antígenos glucídicos de grupo sanguíneo Pk, P y P1 están estructuralmente relacionados. Originariamente todos pertenecían al sistema de grupo sanguíneo P (ISBT 003); sin embargo, debido a que su expresión involucra loci y caminos biosintéticos distintos, la Sociedad Internacional de Transfusión Sanguínea reasignó al antígeno P al recientemente creado grupo sanguíneo GLOB (ISBT 028) y al antígeno Pk a la colección GLOB (ISBT 209).1
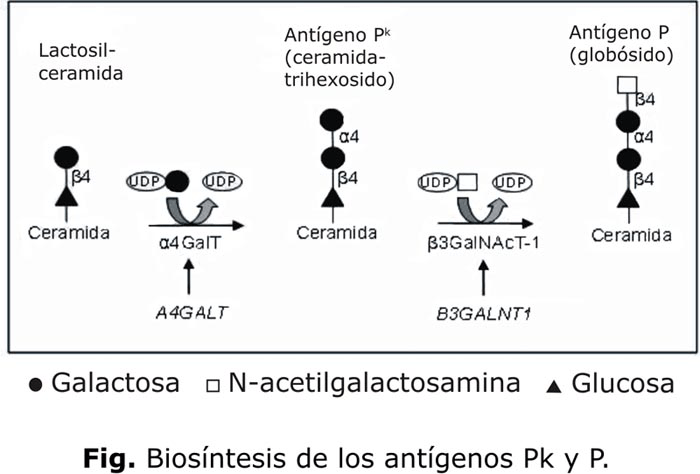
Al igual que para los antígenos del sistema ABO, su expresión se explica bioquímicamente por la presencia del sustrato precursor y de las enzimas (glicosiltransferasas) que catalizan las correspondientes vías sintéticas (figura).
Estos antígenos histohemáticos, presentes en los eritrocitos, plaquetas, linfocitos, fibroblastos, placenta y células uroepiteliales,2-5 se disponen en la población en 5 fenotipos o patrones individuales. En otras palabras, los individuos pueden ser agrupados en los distintos fenotipos dependiendo de los antígenos expresados en sus células (tabla).
Más del 99,999 % de la población pertenece a los fenotipos P1 o P2. Ambos expresan los antígenos públicos P y Pk en sus células y se diferencian entre sí por la expresión del antígeno P1.
Los 3 fenotipos restantes, p, P2k y P1k, son extremadamente raros. Estos individuos se caracterizan por carecer de antígenos presentes en prácticamente toda la población y formar anticuerpos "naturales" y "regulares" contra ellos. Tanto la ausencia de los antígenos como los anticuerpos presentes en todos ellos, son causa de las peculiaridades y complicaciones transfusionales, obstétricas e infecciosas que se describen a continuación.
Bases moleculares de los fenotipos p y Pk ]]>
Las bases moleculares de estos fenotipos excepcionales fueron descritas recientemente. El gen B3GALNT1 (antes llamado B3GALT3) que codifica para la 3-b-N-acetilgalactosaminil-transferasa (P sintetasa) fue clonado en 20006 y se ha demostrado que mutaciones en este gen llevan a los fenotipos Pk (P2k y P1k)7 que tienen en común la ausencia del antígeno P en sus células.
Los individuos con el llamado fenotipo p, carecen de los antígenos P1, P y Pk en sus células.
Este ocurre por la ausencia de actividad de la 4-a-galactosiltransferasa (a4Gal-T o Pk sintetasa), una proteína de membrana de tipo II con 353 aminoácidos,8 encargada cuando funciona de sintetizar al antígeno Pk. El gen A4GALT que la codifica, fue también clonado en 2000 independientemente por 3 grupos.8-10 Cuatro mutaciones en este gen que destruyen la actividad de la enzima,8,9 fueron originariamente reportados. Actualmente se conocen muchas otras mutaciones causales del fenotipo p y otras aparentemente sin efecto en la actividad enzimática de la a4Gal-T.9
La genética que gobierna la expresión del antígeno P1 aún no se ha dilucidado. El gen que codifica para la glicosiltransferasa no se ha clonado y la razón por la que el fenotipo p esta siempre asociado con la ausencia del antígeno P1 tampoco se comprende, aunque existen diferentes teorías. Un modelo sugiere que la misma a4Gal-T es capaz de transferir residuos galactosil a ambos lactosilceramida y paraglobósido, pero para usar este último como aceptor, es necesaria una proteína regulatoria.11 Otro modelo propone un solo gen con 3 alelos: un alelo codificando para la a4Gal-T con lactosilceramida y paraglobósido como aceptores, un alelo con lactosilceramida solo, y el tercer alelo codificando para una forma inactiva de la transferasa.12 Sin embargo, no se halló correlación con la mutación M37V con el fenotipo P1+/_ y no se ha hallado otros polimorfismos en el gen Pk que avalen esta teoría.8,9 Un tercer modelo sugiere que existen 2 enzimas distintas, requiriendo que ambas estén inactivas para causar el fenotipo p.11
Importancia transfusional de los fenotipos p y Pk
]]>
Transfundir a estos individuos es una tarea muy dificultosa, ya que presentan anticuerpos de clase IgM y/o IgG "naturales" y "regulares" contra los antígenos de alta incidencia2 faltantes. Esto significa que en estas personas encontramos siempre anticuerpos plasmáticos contra antígenos públicos sin mediar un episodio inmunizante como una transfusión o embarazos.Frente a la necesidad de una transfusión de glóbulos rojos, deben recibir sangre con su mismo fenotipo o caso contrario, ocurriría una reacción hemolítica transfusional debido a la incompatibilidad inmunohematológica. Estos anticuerpos aglutinan y/o hemolizan los glóbulos rojos de prácticamente toda la población, excepto aquellos con el mismo fenotipo. Su excepcionalidad es tal, que solo 5,8 individuos por millón de habitantes son compatibles con el paciente, sin tener en cuenta otro tipo de compatibilidad inmunohematológica siempre necesaria como el sistema ABO y el RhD, los cuales reducen aún más las posibilidades de encontrar donantes compatibles. Dado lo impracticable de esta aproximación, las opciones son realizar la criopreservación de GR autólogos, donación predepósito para cirugías programadas, búsqueda de individuos compatibles en la familia del paciente (se heredan como rasgos autosómicos recesivos y evidentemente tendremos mayor posibilidades de encontrarlos allí) o solicitar asistencia a bancos de fenotipos raros.
Importancia obstétrica de los fenotipos p y Pk
A pesar que los antígenos Pk y P están bien desarrollados en el feto y el neonato, la enfermedad hemolítica feto-neonatal suele ser de leve a moderada. Una complicación más frecuente y grave para las gestantes con fenotipo p o Pk es la mayor tasa de abortos espontáneos que se observan durante el primer y segundo trimestre. Contrariamente a la baja cantidad de antígeno en el feto temprano, la placenta expresa gran cantidad de estos antígenos y se ha sugerido que es el blanco principal y la razón de los abortos, 5 probablemente en mayor medida relacionado con el componente IgG anti-P.13 Diversas terapéuticas se han propuesto para el manejo de estas gestantes aloinmunizadas: plasmaféresis sola y/o gammaglobulinas endovenosas, pero debido a su baja prevalencia no hay evidencia de la superioridad de una sobre otra.
]]>
Mucho se investiga en este campo, fundamentalmente interesa la función de estos antígenos como receptores para virus y bacterias y la resistencia relativa de los distintos fenotipos a la infección por ellos.
De las más importantes que se citan:
- El antígeno P es un receptor para el parvovirus-B1914 causante del eritema infeccioso en niños (también llamada la quinta enfermedad) y que a veces se complica con anemia aplásica severa. La replicación viral y posterior lisis celular está limitada a los precursores eritroides tempranos que expresan el receptor P15 y se ha reportado que los individuos que carecen de este antígeno parecen ser naturalmente resistentes a dicha infección.16
- La hemoglobinuria paroxística a frigore es una enfermedad más frecuente en niños, donde los glóbulos rojos antígeno P positivos son hemolizados por un efímero auto-antiP.17 Este cuadro generalmente es precedido por una infección viral.
- El virus de la inmunodeficiencia humana (VIH) interacciona con el antígeno Pk (CD77) y el CD4 para entrar a la célula 18 y la expresión de estos antígenos y la resistencia a la infección in vitro es un campo de estudio muy actual.
- Las verotoxinas producidas por Escherichia coli, se unen con el antígeno Pk e inducen apoptosis a través de la vía de las caspasas,19 y podría ser la razón de la severa trombocitopenia vista durante el síndrome urémico hemolítico.8,20
]]> - Los antígenos P1, Pk y P funcionan como aceptores celulares para la Escherichia coli uropatogénica. También la fimbriae de la E coli pielonefritogénica expresa moléculas que se unen con la Pk.21
El interés en los antígenos de grupo sanguíneo de naturaleza glicolipídica es cada vez mayor debido a su relación con patógenos y enfermedades.
Sin embargo, los abortos espontáneos a repetición parecen contrarrestar cualquier efecto beneficioso ganado por la inmunidad frente a patógenos.
Resumiendo, dado la excepcionalidad de este cuadro y su gran importancia clínica, estos individuos deben ser identificados y dentro de lo posible, otorgársele un adecuado soporte transfusional y obstétrico.
REFERENCIAS BIBLIOGRÁFICAS ]]>
1. Daniels GL, Fletcher A, Garratty G. Blood group terminology 2004: From the International Society of Blood Transfusion committee on terminology for red cell surface antigens. Vox Sang 2004;87:304-16.
2. Daniels G. Human Blood Groups. 2 ed. Oxford: Blackwell Scientific; 2002.
3. Klein G, Manneborg-Sandlund A, Ehlin-Henriksson B. Expression of the BLA antigen, defined by the monoclonal 38.13 antibody, on Burkitt lymphoma lines, lymphoblastoid cell lines, their hybrids and other B-cell lymphomas and leukemias. Int J Cancer 1983;31:535-42.
4. Mangeney M, Richard Y, Coulaud D. CD77: An antigen of germinal center B cells entering apoptosis. Eur J Immunol 1991;21:1131-40.
5. Lindstrom K, Von Dem Borne AE, Breimer ME. Glycosphingolipid expression in spontaneously aborted fetuses and placenta from blood group p women. Evidence for placenta being the primary target for anti-Tja-antibodies. Glycoconj J 1992;9:325-9.
6. Okajima T, Nakamura Y, Uchikawa M. Expression cloning of human globoside synthase cDNAs. Identification of beta 3Gal-T3 as UDP-N-acetylgalactosamine: Globotriaosylceramide beta 1,3-N-acetylgalactosaminyltransferase. J Biol Chem 2000;275:40498-503.
7. Hellberg A, Poole J, Olsson ML. Molecular basis of the globoside-deficient P(k) blood group phenotype. Identification of four inactivating mutations in the UDP-N-acetylgalactosamine: Globotriaosylceramide 3-beta-N-acetylgalactosaminyltransferase gene. J Biol Chem 2002;277:29455-9.
8. Iwamura K, Furukawa K, Uchikawa M. The blood group P1 synthase gene is identicalto the Gb3/CD77 synthase gene. A clue to the solution of the P1/P2/p puzzle. J Biol Chem 2003;278:44429-38.
9. Hellberg A, Steffensen R, Yahalom V. Additional molecular bases of the clinically important p blood group phenotype. Transfusion 2003;43:899-907.
10. Keusch JJ, Manzella SM, Nyame KA. Cloning of Gb3 synthase, the key enzyme in globo-series glycosphingolipid synthesis, predicts a family of alpha 1, 4-glycosyltransferases conserved in plants, insects, and mammals. J Biol Chem 2000;275:25315-21.
11. Naiki M, Marcus DM. An immunochemical study of the human blood group P1, P, and PK glycosphingolipid antigens. Biochemistry 1975;14:4837-41.
12. Graham HA, Williams AN. A genetic model for the inheritance of the P, P1 and Pk antigens. Immunol Commun 1980;9:191-201.
13. Cantin G, Lyonnais J. Anti-PP1Pk and early abortion. Transfusion 1983;23:350-1.
14. Brown KE, Anderson SM, Young NS. Erythrocyte P antigen: Cellular receptor for B19 parvovirus. Science 1993;262:114-7.
15. Brown KE, Young NS. Parvovirus B19 infection and hematopoiesis. Blood Rev 1995;9:176-82.
16. Brown KE, Hibbs JR, Gallinella G. Resistance to parvovirus B19 infection due to lack of virus receptor (erythrocyte P antigen). N Engl J Med 1994;330:1192-6.
17. Heddle NM. Acute paroxysmal cold hemoglobinuria. Transfus Med Rev 1989;3:219-29.
18. Fantini J, Hammache D, Pieroni G, Yahi N. Role of glycosphingolipid microdomains in CD4-dependent HIV-1 fusion. Glycoconj J 2000;17:199-204.
19. Kiyokawa N, Mori T, Taguchi T. Activation of the caspase cascade during Stx1-induced apoptosis in Burkitt's lymphoma cells. J Cell Biochem 2001;81:128-42.
20. Cooling LL, Walker KE, Gille T, Koerner TA. Shiga toxin binds human platelets via globotriaosylceramide (Pk antigen) and a novel platelet glycosphingolipid. Infect Immun 1998;66:4355-66.
21. Stapleton AE, Stroud MR, Hakomori SI, Stamm WE. The globoseries glycosphingolipid sialosyl galactosyl globoside is found in urinary tract tissues and is a preferred binding receptor In vitro for uropathogenic Escherichia coli expressing pap-encoded adhesins. Infect Immunol 1998;66:3856-61.
Recibido: 3 de febrero de 2008.
Aprobado: 8 de abril de 2008.
]]>
]]>
Dr. Daniel de La Vega Elena. Virasoro 1249 (2000) Rosario. Provincia de Santa Fe, Argentina. Te/Fax: 0054 -341 - 4858825. e-mail: daniel.delavega@yahoo.comLaboratorio de Inmunohematología. Servicio de Medicina Transfusional. Hospital Italiano Garibaldi, Rosario, Argentina. ]]>
{kind=link}
{kind=link}