ARTíCULO ORIGINAL
Representación del desarrollo de la espongiosa primaria por medio de un sistema de reacción-difusión: Una hipótesis sobre el inicio de la formación de hueso inmaduro. Parte 1: Descripción del modelo
Representation of the development of the primary spongiosa by means of a reaction-diffusion system: a hypothesis on the onset of immature bone formation. Part 1: Model description
MSc. Oscar Rodrigo López-VacaI, Ing. Diego Alexander Garzón-AlvaradoII
IFacultad de Ingeniería Mecánica, Universidad Santo Tomás. Bogotá, Colombia. ]]> IIDepartamento de Ingeniería Mecánica y Mecatrónica, Universidad Nacional de Colombia. Bogotá, Colombia.
RESUMEN
Se presenta un modelo bioquímico que predice la formación de la arquitectura de la espongiosa primaria, a partir de la interacción de 2 factores moleculares: VEGF (factor de crecimiento endotelial vascular) y MMP13 (metaloproteinasas 13). Se supone que el MMP13 regula la degradación del cartílago y el VEGF permite la vascularización y el avance del frente de osificación mediante la presencia de osteoblastos. El acople de este conjunto de moléculas se representa mediante ecuaciones de reacción-difusión con parámetros en el espacio de Turing, y se obtiene como resultado un patrón espacio-temporal estable que da paso a la formación de las trabéculas presentes en el tejido esponjoso.
Palabras clave: espongiosa primaria, osificación endocondral, sistemas de reacción-difusión.
ABSTRACT
A biochemical model is presented which predicts the formation of the architecture of the primary spongiosa, based on the interaction of two molecular factors: VEGF (vascular endothelial growth factor) and MMP-13 (metalloproteinases-13). It is assumed that MMP-13 regulates cartilage degradation, and VEGF allows vascularization and the advance of the ossification front through the presence of osteoblasts. The coupling of this set of molecules is represented by means of reaction-diffusion equations with Turing space parameters, and a stable spatio-temporal pattern is obtained which leads to the formation of the trabeculae present in the spongy tissue.
Key words: primary spongiosa, endochondral ossification, reaction-diffusion systems.
]]>
INTRODUCCIÓN
La mayoría de los huesos largos del sistema esquelético de los mamíferos se desarrollan a partir de un proceso denominado crecimiento endocondral.1-4 En este proceso el crecimiento y la elongación de los huesos se logran por la continua adición de cartílago y subsecuente reemplazo por tejido óseo.
Durante el proceso de diferenciación de los condrocitos la composición de la matriz cambia de forma súbita, a través de la producción de otros componentes, como el colágeno tipo X, la expresión de metaloproteínas y la subsecuente calcificación. Al mismo tiempo los vasos sanguíneos invaden el cartílago calcificado trayendo los osteoblastos los cuales construyen el hueso inmaduro. Los condrocitos en el cartílago de crecimiento están sometidos a la influencia del exceso de factores extracelulares incluyendo factores locales sistémicos y solubles, así como componentes de la matriz extracelular. Varios estudios5-8 suministran evidencia que la proliferación de condrocitos en el cartílago de crecimiento está bajo el control de un lazo cerrado local que depende de la ubicación espacial y temporal; y que, principalmente, involucra las señales moleculares sintetizadas por los condrocitos: hormona paratiroidea-relacionada péptida (PTHrP), Indian hedgehog (Ihh), factor de crecimiento transformante (TGFb), proteínas morfogéneticas óseas (BMPs), factor de crecimiento endotelial vascular (VEGF), metaloproteinasas de matriz tipo 9 conocida como gelatinasaB (MMP9) y el factor de transcripción RUNX2. Juntos interactúan entre sí en un bucle retroalimentado, para regular la tasa a la cual los condrocitos dejan la zona proliferativa, diferencian a células hipertróficas y dan paso a la formación de hueso inmaduro.5,9,10
El proceso de osificación y crecimiento endocondral ha sido objeto de estudio desde hace varios años y se han desarrollado diferentes modelos in-silico comprobados mediante reportes histológicos y experimentación in-vivo que han tratado de explicar el proceso de formación de hueso mediante este mecanismo.7-9,11-16 Por ejemplo, Courtin y otros.11 han hecho la comparación entre la secuencia de eventos morfológicos involucrados en la formación de hueso embriónico y las características espacio-temporales autoorganizadas generadas por un modelo de reacción-difusión relacionado con el metabolismo de la mineralización del hueso perióstico. En su trabajo se obtienen las estructuras tridimensionales (mediante simulación computacional) con un estrecho parecido con la arquitectura interna primaria del periostio de los huesos largos. Posteriores trabajos como los de Garzón y otros7 han planteado diferentes hipótesis sobre la interacción de factores mecánicos, celulares, y moleculares que dan paso a la formación de los centros secundarios de osificación en las epífisis de los huesos largos,8,13 el desarrollo y crecimiento de los huesos,14 y la formación de hueso primario.15 En estas hipótesis se plantea que los procesos biológicos y la interacción entre los diferentes factores, pueden ser representados mediante modelos matemáticos en donde la retroalimentación química entre factores moleculares reactivos a través de mecanismos de reacción-difusión pueden explicar los patrones espaciales estables encontrados en algunos sistemas biológicos.
Hasta donde los autores conocen, no se han realizado investigaciones sobre la acción de los diferentes factores celulares, mecánicos y moleculares, sobre el desarrollo y producción de la arquitectura de la espongiosa primaria. Por tanto, en este trabajo se presenta la formulación matemática de un modelo bioquímico que explica el desarrollo de la arquitectura del hueso trabecular, partiendo del supuesto de la interacción de dos factores moleculares, los cuales, a través de un mecanismo reacción-difusión, generan un patrón espacio-temporal estable que da paso a la formación de las trabéculas presentes en el tejido esponjoso primario.
MÉTODOS
a) Hipótesis requerida para el desarrollo de la espongiosa primaria usando sistemas de reacción-difusión ]]>
La hipótesis principal de este trabajo se basa en la formación de la espongiosa primaria a partir del tejido cartilaginoso epifisial mediante la interacción de 2 señales moleculares que se difunden e interactúan en la matriz extracelular cartilaginosa. Para esto se supone la existencia de un sistema de reacción-difusión de 2 moléculas primordiales, como son el VEGF y la MMP13, que puede conducir a un patrón estable en el tiempo e inestable en el espacio similar a los patrones presentes en la estructura del hueso trabecular durante la osificación endocondral.La presencia de MMP13 liberado por los condrocitos hipertróficos permite la degradación de los componentes de la matriz cartilaginosa (colágeno y agrecanos) y da paso a la invasión vascular y del frente de osificación.10,17,18 Esta invasión vascular es facilitada por la presencia del VEGF expresado por los condrocitos hipertróficos. Esto supone que al existir MMP13 y VEGF en todas las regiones del cartílago epifisial, en aquellas zonas en donde se encuentren altas concentraciones de VEGF se producirá un oportuno control de la invasión de células endoteliales, osteoclastos, condroclastos y osteoblastos en el desarrollo la osificación primaria.19 De igual forma, en donde exista una alta concentración de MMP13 se degradará completamente el cartílago, dando origen a la arquitectura del hueso trabecular. Por lo tanto, se puede suponer que en el desarrollo de la arquitectura de la espongiosa primaria debe existir una regulación del MMP13 por parte del VEGF (mecanismo inhibidor) de forma tal que se detenga la degradación del cartílago y empiece la invasión del frente de osificación.
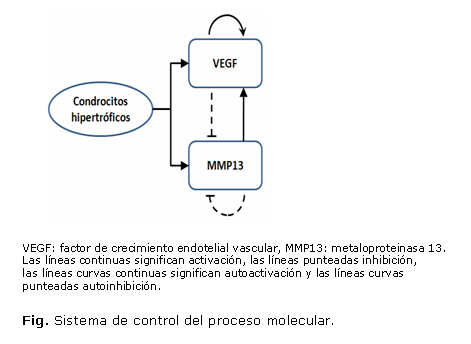
b) Descripción del modelo
El proceso regulatorio que se propone en este modelo está basado en un sistema activador-sustrato. La figura muestra la relación de las señales moleculares producidas por la hipertrofia de los condrocitos. El proceso indica que existe un lazo de control entre el VEGF (factor activador) y el MMP13 (sustrato), donde el VEGF se autoactiva e inhibe la producción de MMP13 deteniendo el proceso de degradación y dando paso a la mineralización de la matriz de cartílago remanente.19 Por otro lado, se supone que el MMP13 se autoinhibe pero activa la producción de VEGF; a este lazo se le conoce como sistema de retroalimentación positiva. El VEGF ayuda a la invasión vascular y traerá consigo las células osteogénicas que permiten la construcción de la espongiosa primaria. Se ha observado que este proceso genera trabéculas regulares y patrones altamente repetibles durante el proceso de osificación, similares a los hallados en los patrones de Turing.20
El mecanismo regulatorio es modelado por medio de ecuaciones de reacción-difusión, donde el término «de reacción» (síntesis de factores extracelulares solubles) es considerado dependiente de la concentración de los reactantes y de la presencia de los condrocitos hipertróficos. De acuerdo con esto, la hipótesis se sustenta en que el origen de los patrones presentes en la espongiosa primaria podría corresponder, desde un punto de vista matemático, a los patrones que ocurren en el espacio de Turing cuando 2 reactantes químicos interactúan.
La definición de las relaciones indicadas en la figura puede ser cuantificada por medio de ecuaciones que establecen cambios locales de los factores extracelulares solubles, y en la concentración de sangre:
]]>
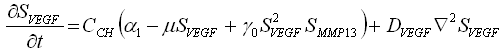 (1a)
(1a)
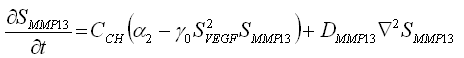 (1b)
(1b)
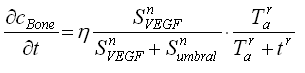 (1c)
(1c)
donde ![]() es la concentración de condrocitos hipertróficos
es la concentración de condrocitos hipertróficos ![]() y
y ![]() representa las concentraciones de VEGF y MMP13, respectivamente. Los restantes son parámetros del modelo:
representa las concentraciones de VEGF y MMP13, respectivamente. Los restantes son parámetros del modelo: ![]() y
y ![]() son términos que cuantifican la producción de cada factor molecular por los condrocitos hipertróficos; m es una constante que cuantifica la inhibición de la producción de VEGF por su exceso;
son términos que cuantifican la producción de cada factor molecular por los condrocitos hipertróficos; m es una constante que cuantifica la inhibición de la producción de VEGF por su exceso; ![]() regula la interacción no lineal entre la concentración de MMP13-VEGF cuantificando la concentración o inhibición de cada factor molecular;
regula la interacción no lineal entre la concentración de MMP13-VEGF cuantificando la concentración o inhibición de cada factor molecular; ![]() y
y ![]() son los coeficientes de difusión de VEGF y MMP13, respectivamente.
son los coeficientes de difusión de VEGF y MMP13, respectivamente.
En la interpretación biológica de las anteriores ecuaciones el término ![]() representa la activación no lineal de
representa la activación no lineal de ![]() (producción de VEGF por la presencia de MMP13) y el consumo no lineal de
(producción de VEGF por la presencia de MMP13) y el consumo no lineal de ![]() (por la presencia de VEGF). La ecuación (1c) representa la activación de la tasa de producción de hueso por la presencia de altas cantidades de VEGF, la cual se regula a medida que transcurre el tiempo. En esta ecuación h es una constante que regula la producción de hueso en el tiempo;
(por la presencia de VEGF). La ecuación (1c) representa la activación de la tasa de producción de hueso por la presencia de altas cantidades de VEGF, la cual se regula a medida que transcurre el tiempo. En esta ecuación h es una constante que regula la producción de hueso en el tiempo; ![]() representa el valor de la concentración de VEGF con la cual comienza el proceso de osificación.
representa el valor de la concentración de VEGF con la cual comienza el proceso de osificación. ![]() es el tiempo requerido para el proceso de calcificación del cartílago y
es el tiempo requerido para el proceso de calcificación del cartílago y ![]() representa el tiempo que limita la producción de hueso.
representa el tiempo que limita la producción de hueso.
]]>
DISCUSIÓN
En este trabajo se presentó la formulación de un modelo biorregulatorio basado en un conjunto de ecuaciones de reacción-difusión para predecir la formación de la arquitectura de la espongiosa primaria. La aplicación del modelo de reacción-difusión con parámetros en el espacio de Turing es un área de constante trabajo y controversia en biología. Garzón y otros,8,14,21,22 Courtin y otros11 y Cramping y otros23 utilizaron en sus trabajos modelos de reacción-difusión para simular diferentes procesos biológicos, y obtuvieron como resultado que el empleo de estos sistemas puede llegar a explicar muchos fenómenos biológicos complejos en donde la formación de patrones sea una variable constante.
La hipótesis presentada plantea que el origen de la espongiosa primaria es internamente controlado por células cartilaginosas, esto se logra a través de 2 bioquímicos reactantes, VEGF y MMP13. Estos factores no son los únicos que actúan en la osificación endocondral, existen otros tantos, entre los que se cuentan Ihh, PTHrP, Runx2, BMP,5,9,10,24 y que probablemente influyen de igual forma en la formación de hueso trabecular. Hasta la fecha se ha hecho un gran esfuerzo para entender completamente el papel de cada una de estas sustancias, cómo interactúan y qué procesos regulan. Es posible que VEGF y MMP13 no sean los factores que controlan completamente el proceso de osificación endocondral, pero la existencia de un mecanismo activador-sustrato garantiza una alta estabilidad para el desarrollo de este proceso biológico.
En el desarrollo del modelo se asumió, para las condiciones iníciales, que el factor activador es liberado por los condrocitos hipertróficos, al igual que el sustrato, sin embargo, el tipo de inestabilidad espacial obtenida es independiente de las condiciones iníciales. No obstante este modelo es muy estable y robusto con respecto a las condiciones iníciales y el rango de parámetros.
REFERENCIAS BIBLIOGRÁFICAS
1. Carter DR, Beaupré GS. Skeletal Function and Form Mechanobioloy of Skeletal Development. Aging and Regeneration. New York: Cambridge University Press; 2001.
2. Shapiro F. Developmental Bone Biology. Pediatric Orthopedic Deformities. 2002;953.
3. Shier D. Hole's Human Anatomy & Physiology. Boston: McGraw-Hill; 2001.
4. Roberts W. Bone development and function: genetic and environmental mechanisms. Seminars in Orthodontics. 2004;10:100-22.
5. Provot S, Schipani E. Molecular mechanisms of endochondral bone development. Bioch. Biph R Biochem Bioph Res Co . 2005;328:658-65.
6. Goldring MB, Tsuchimochi K, Ijiri K. The Control of Chondrogenesis. J Cell Biochem . 2006;44:33-44.
7. Garzón-Alvarado D, Garcia Aznar J, Doblaré M. The early bone epiphysis formation: a numerical simulation. J Biomech. 2006;39:S642.
8. Garzón-Alvarado D, Garcia Aznar JM, Doblaré M. Appearance and location of secondary ossification centres may be explained by a reaction-diffusion mechanism. Computers in biology and medicine. 2009;39:554-61.
9. Brouwers JEM, van Donkelaar CC, Sengers BG, Huiskes R. Can the growth factors PTHrP, Ihh and VEGF, together regulate the development of a long bone? J Biomech . 2006;39: 2774-82.
10. Mackie EJ, Ahmed YA, Tatarczuch L, Chen KS, Mirams M. Endochondral ossification: how cartilage is converted into bone in the developing skeleton. Int J Biochem Cell B. 2008;40:46-62.
11. Courtin B, Perault AM, Staub JF. A Reaction-Diffusion Model for Trabecular Architecture of Embryonic Periosteal Long Bone. Complexity International. 1997;4:1-17.
12. Blumer MJF, Longato S, Fritsch H. Structure, formation and role of cartilage canals in the developing bone. Ann Anat. 2008;190:305-15.
13. Garzón-Alvarado DA, Peinado Cortés LM, Cárdenas Sandoval RP. A mathematical model of epiphyseal development: hypothesis on the cartilage canals growth. Comp Meth Biomech Biomed Eng. 2010;13:765-72.
14. Garzón-Alvarado, DA, García-Aznar JM, Doblaré MA. Reaction-diffusion model for long bones growth. Biomech Mod Mechan 2009;8:381-95 ().
15. Peinado Cortés LM, Vanegas Acosta JC, Garzón Alvarado DA. A mechanobiological model of epiphysis structures formation. J. Theor Biol. 2011;287:13-25.
16. Van Donkelaar CC, Huiskes R. The PTHrP-Ihh feedback loop in the embryonic growth plate allows PTHrP to control hypertrophy and Ihh to regulate proliferation. Biomech Mod Mechan. 2007;6:55-62.
17. Krane SM, Inada M. Matrix metalloproteinases and bone. Bone. 2008;43:7-18.
18. Troeberg L, Nagase H. Proteases involved in cartilage matrix degradation in osteoarthritis. BBA Protein and Proteomics. 2011;1824:133-45.
19. Filvaroff EH. VEGF and bone. J Musc Neur Inter. 2003;3:304-7; discussion 320-1.
20. Turing AM. The Chemical Basis of Morphogenesis. Biol Sc. 1952;237:37-72.
21. Garzón-Alvarado D, Ramírez Martinez AM. A biochemical hypothesis on the formation of fingerprints using a turing patterns approach. Theor Biol Med Mod. 2011;8:24.
22. Garzón-Alvarado D, Velasco M, Narváez-Tovar C. Self-Assembled Scaffolds Using ReactionDiffusion Systems: a Hypothesis for Bone Regeneration. J Mech Med Biol. 2011;11:231.
23. Cramping EJ, Maini PK. Reactiondiffusion models for biological pattern formation. Meth Appl Mech. 2001;8:415-28.
24. Carlevaro MF, Cermelli S, Cancedda R, Descalzi Cancedda F. Vascular endothelial growth factor (VEGF) in cartilage neovascularization and chondrocyte differentiation: auto-paracrine role during endochondral bone formation. Journal of cell science. 2000;113:59-69. Pt 1.
]]>
Recibido: 5 de septiembre de 2011.Aprobado: 20 de septiembre de 2011.
Oscar Rodrigo López-Vaca. Facultad de Ingeniería Mecánica, Universidad Santo Tomás. Bogotá, Colombia. Correo electrónico: osrolova@gmail.com ]]>