VARIABILIDAD MOLECULAR EN AISLAMIENTOS DE Phytophthora nicotianae VAN BREDA DE HAAN
MOLECULAR VARIABILITY IN Phytophthora nicotianae VAN BREDA DE HAAN ISOLATES
Belkis Peteira*, Verónica Toledo** y B. Martínez*
*Grupo de Fitopatología, División de Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. Correo eléctrónico: bpeteira@censa.edu.cu; ** Instituto de Investigaciones del Tabaco. San Antonio de los Baños, La Habana, Cuba
]]>
RESUMEN
La pata prieta, causada por el patógeno Phytophthora nicotianae es una de las enfermedades fungosas importantes que atacan al cultivo del tabaco. Para apoyar de manera racional el programa de mejoramiento del cultivo para la obtención de variedades resistentes se hace necesario el conocimiento de la variabilidad de los aislamientos del patógeno. Las técnicas moleculares son una herramienta útil en tal sentido. Se caracterizaron a través ITS y corte de los fragmentos ITS amplificados y de RAPDs, un grupo de 50 aislamientos representativos de diferentes regiones del país, razas y características culturales. Los ITS rindieron un producto para todos los aislamientos de 850pb y la digestión de los mismos solo fue posible con la enzima MspI. A pesar de obtenerse dos patrones de restricción con 300 y 400pb estas características no se correlacionan con la patogenicidad, ni el origen geográfico, aunque es posible la aplicación de esta técnica para corroborar el diagnóstico de esta especie. Los RAPD demostraron la alta variabilidad molecular de los aislamientos en estudio. Sin embargo, los grupo formados no pudieron correlacionarse con las características seleccionadas.
Palabras clave: Phytophthora nicotianae; caracterización molecular; RAPD; ITS
ABSTRACT
The black shank, caused by the phytopathogen Phytophthora nicotianae is one of the fungal diseases in tobacco crop. To support, in a rational way, the breeding program for the obtaninig of resistant varieties in this crop, it is necessary to know about the variablity in the pathogen isolates. The molecular techniques are a useful tool in such way. By the ITS, the cutting of ITS amplified fragments and RAPDs, 50 representative isolates from different regions of the country, strains and cultural characters, were studied. ITS presents a common product 850bp for all the isolates and their digestion was possible only with the MspI enzyme. Two restriction patterns were obtained with 300 and 400bp, but non of them had a correlation with pathogenicity or geographic origin; however it is posible to use this technique for diagnostic porposes at species level. RAPDs analysis showed a high molecular variation between the isolates tested. The cluster from the molecular data did not correlate with the other characteristics selected.
Key words: Phytophthora nicotianae; molecular characterization; RAPD; ITS
]]> INTRODUCCIÓN
La pata prieta, causada por el patógeno Phytophthora nicotianae es una de las enfermedades fungosas más importantes que atacan al cultivo del tabaco. Con el fin de contribuir al mejoramiento genético de las variedades, como una de las tácticas para el control de la enfermedad, se hace necesario conocer la variabilidad del patógeno en estudio. Las técnicas moleculares, de manera general, pueden ser empleadas como una herramienta útil en la caracterización de los aislamientos circulantes.
Para el género Phytophthora se han empleado diferentes técnicas moleculares con resultados importantes en diagnóstico, taxonomía, filogenia, mapeo de genes, evolución poblacional de diferentes especies y en estudios de variabilidad (1,2).
Entre las técnicas más empleadas se destacan: Polimorfismo del ADN Amplificado al Azar (RAPD, Random Amplified Polymorphic DNA), Polimorfismo de la Longitud de los Fragmentos de Restricción (RFLP, Restriction Fragment Length Polymorphism) y la amplificación del Espaciador Interno Transcripto (ITS, Internal Transcribed Spacer) (3,4,5).
El Instituto de Investigaciones del Tabaco cuenta con una colección de P. nicotianae, la cual ha sido caracterizada desde el punto de vista morfológico, cultural y patogénico. Como parte del completamiento de la caracterización de la colección de este patógeno, en el trabajo se propone como objetivo la caracterización de la variabilidad molecular de un grupo representativo de aislamientos a partir de los marcadores RAPD, los ITS y el corte de los fragmentos amplificados con enzimas de restricción.
MATERIALES Y MÉTODOS
Para el análisis de la variabilidad a nivel molecular fueron seleccionados 50 aislamientos representativos, teniendo en cuenta origen geográfico, raza y patrón cultural (Tabla 1). Los mismos fueron sembrados en el medio de cultivo caldo de papa dextrosa durante 15 días a 27ºC y estático, en oscuridad. Transcurrido este tiempo se cosechó el micelio, se lavó con agua destilada estéril y se secó con papel de filtro. Las muestras fueron conservadas a -20ºC hasta su utilización.
Extracción de ADN total
La extracción de ADN total se realizó por el método descrito por Dellaporta et al. (6). El ADN obtenido fue resuspendido en 100mL de buffer TE 1X (Tris-EDTA) y se conservó a -20ºC. La calidad del ADN se constató por electroforesis en geles de agarosa al 0,8% en solución amortiguadora de corrida TBE 0,5X, a 90 volts y teñidos con bromuro de etidio (5mg.mL-1) y observados en un transiluminador (LKB, Pharmacia). La concentración se estimó por la medición de la densidad óptica a 260nm en un espectrofotómetro Ultrasepec Plus Spectrophotometer Pharmacia LKB, según Sambrook et al. (7).
]]> Condiciones de la PCR con cebadores ITSEl ADN de los aislamientos en estudio se amplificó empleando para ello los cebadores ITS 4 y 5 (ITS 4: 5´- GGA AGT AAA AGT CGT AAC AAG G -3´; ITS 5: 5´- TCC TCC GCT TAT TGA TAT GC -3´). La mezcla de amplificación, con volumen total de 25uL, contenía: 10mM Tris-HCl a pH 8,3; 50mM KCl, 2mM MgCl2 , 0.001% de gelatina, 100uM de cada dNTPs, 10 picomoles de cada cebador, 100ng de ADN genómico aproximadamente y 1U de Taq ADN polimerasa (Amplicen, CENSA). El programa de amplificación empleado fue el siguiente: 1 ciclo de 94ºC durante 3 minutos, 34 ciclos de 94ºC/1 minuto, 42ºC/1,5 minutos y 72ºC/1,5 minutos y un ciclo final de extensión de 72ºC, durante 5 minutos. Los fragmentos amplificados se observaron después de su separación en geles de agarosa al 1,5% en idénticas condiciones a las descritas anteriormente, utilizando como marcador de peso molecular el conocido como 1Kb Plus DNA Ladder (Vitrogen).
Condiciones para los PCR -RFLP
Los fragmentos amplificados por ITS -PCR se digirieron con las enzimas de restricción: EcoR I, EcoR V, Msp I, Hind III, BamH I, Alu I y Hae II , todas provenientes de la casa comercial Sigma, UK. Se tomaron tres aislamientos al azar y se realizaron las digestiones de 15uL del ADN amplificado, siguiendo las especificaciones descritas para cada enzima por el manufacturador en cuanto a la solución amortiguadora a emplear en cada reacción, ajustando la concentración de todas las enzimas empleadas a 2,5U. Las reacciones se mantuvieron a 37ºC en baño de María (Baño Serológico BS02, SIME), toda la noche. Posteriormente, los fragmentos digeridos se separaron en geles de agarosa al 1,5% en solución amortiguadora TBE 0,5X, a 90 volts y se procedió de igual forma a la descrita con anterioridad para su observación. A partir de los resultados de las digestiones, se desarrollaron las reacciones con las enzimas que produjeron fragmentos de corte, para todos los aislamientos en estudio, siguiendo los protocolos descritos previamente.
Condiciones de la PCR con cebadores arbitrarios
La reacción de amplificación se realizó en un volumen total de 25uL que contenía: 10mM Tris-HCl a pH 8,3; 50mM KCl, 2mM MgCl2, 0,001% de gelatina, 100uM de cada dNTPs, 5 picomoles del cebador, 100ng de ADN genómico y 1U de Taq ADN polimerasa (Amplicen, CENSA). El programa de amplificación fue de 45 ciclos de: 1 minuto a 94ºC, 1 minuto a 36ºC y 2 minutos a 72ºC, y un ciclo de 10 minutos a 72ºC y se desarrolló en el termociclador Amplificador Progene. Los productos de la PCR se visualizaron por electroforesis en geles de agarosa al 1,5%, en solución amortiguadora TBE 0,5X (45mM Tris-Borato, 1mM EDTA), a 90 volts y se tiñeron con bromuro de etidio, antes de ser observados al UV.
Se realizó primeramente un experimento con solo dos aislamientos como muestras, para la determinación de los cebadores funcionales. Se utilizaron dos juegos de cebadores: A y F de la firma Operon Technologies. A partir de estos resultados se seleccionaron aquellos iniciadores con amplificación positiva para los aislamientos de Phytophthora estudiados. Los cebadores seleccionados se utilizaron en las posteriores amplificaciones para todos los aislamientos, aplicando para ello idénticas condiciones a las descritas anteriormente.
Los datos de las amplificaciones se organizaron en una matriz binaria, donde el valor 0 se le adjudicó a la ausencia y 1 a la presencia de bandas. Se realizó un análisis de la similitud comparando todas las muestras entre sí, utilizando la fórmula de Nei y Li (8). Con el coeficiente de similitud se calcularon las distancias genéticas y se elaboró un dendrograma, para ambos juegos de cebadores y para cada uno por separado. Se realizaron además diferentes análisis de correspondencia múltiple comparando los resultados moleculares de ambos juegos de cebadores con datos obtenidos en estudios previos de identificación racial, características patogénicas, respuesta al metalaxyl, hospedantes y procedencia de los aislamientos estudiados.
RESULTADOS Y DISCUSIÓN
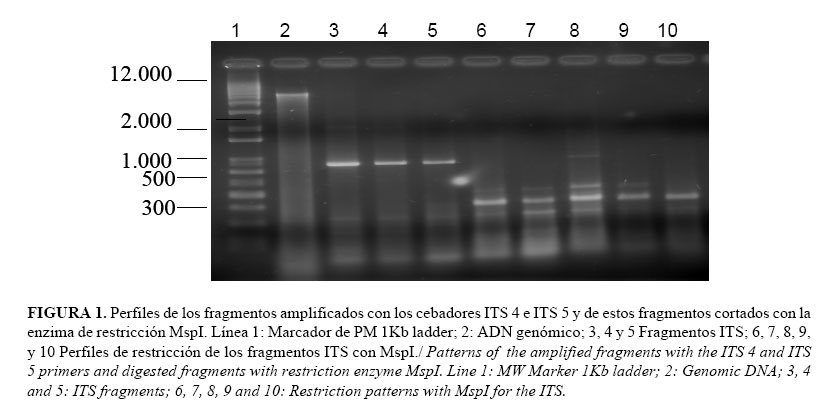
]]> La amplificación de la región ITS a partir de PCR con los cebadores específicos ITS 4 e ITS 5 evidenció gran homogeneidad para todas las muestras analizadas. El fragmento obtenido para todos los aislamientos tuvo un tamaño aproximado de 850 pb (Fig. 1).De las enzimas de restricción utilizadas posteriormente, para digerir el fragmento amplificado, solo rindió producto de corte la enzima Msp I. Los fragmentos amplificados por los cebadores ITS empleados y cortados con la enzima Msp I rindieron dos perfiles diferentes de restricción con diversos fragmentos de tallas de 400 y 300pb (Fig. 1). Sin embargo, estos perfiles tampoco pudieron relacionarse con la patogenicidad ni con el resto de las variables analizadas para los aislamientos estudiados.
Numerosos autores emplean en la actualidad la amplificación de las regiones ITS en estudios de identificación de géneros y especies para diversos organismos. Bonants et al. (9) sugirieron que la detección de patógenos fungosos a través de la PCR es al menos 10 veces más sensible que el ELISA. Además, las técnicas serológicas desarrollas para detectar Phytophthora son casi siempre género-específicas.
La importancia del diagnóstico molecular en el manejo de las enfermedades en diferentes cultivos ha sido enfatizada por Johri et al. (10), debido a que la identificación morfológica es laboriosa y requiere altos niveles de experiencia y puede conllevar a falsas determinaciones.
En el género Phytophthora los ITS han sido ampliamente usados para dilucidar relaciones filogenéticas, diferenciación entre especies y diagnóstico (11, 12, 13). Lee y Taylor (11), estudiaron 27 aislamientos de cinco especies de Phytophthora: P. palmivora, P. megakarya, P. capsici, P. citrophora y P. cinnamomi. A partir de datos referentes a características morfológicas y ecológicas, se pensaba que tales especies estaban relacionadas. El análisis de las secuencias de las regiones ITS I e ITS II demostró relaciones de cercanía entre aislamientos provenientes de cacao de P. capsici y P. citrophora, un linaje común para P. palmivora y P. megakayra y distancias mayores entre P. cinnamomi y el resto de las especies estudiadas. Sin embargo, no se detectó variación intraespecífica o esta fue despreciable.
En el presente caso los aislamientos analizados fueron de la especie P. nicotianae y todos rindieron la banda única anteriormente mencionada (850pb), que por demás coincide con los rangos para el número de bases para la especie encontrada en la literatura (que oscila entre 812 a 860pb, según la región ITS amplificada (14), lo cual confirma la posibilidad del uso de esta técnica para el diagnóstico de esta especie.
Con el objetivo de aumentar la posible variabilidad de las regiones ITS, otros autores digieren las regiones amplificadas con enzimas de restricción. Aún cuando esta técnica es ampliamente utilizada, los resultados más exitosos han sido obtenidos cuando se analizan muestras provenientes de diferentes especies, no así cuando se trata de hallar diferencias a nivel intraespecífico.
Ristaino et al. (3), amplificando las regiones ITS con los cebadores ITS 4 e ITS 5 y posteriormente cortando los amplicones con las enzimas de restricción Rsa I, MspI, Hae III, lograron diferenciar algunas de las especies en estudio. Sin embargo, otras permanecieron inseparables luego de la comparación de los datos obtenidos.
Tripathi et al. (15), trabajaron con los fragmentos ITS amplificados con los cebadores universales ITS 4 e ITS 6 y digeridos posteriormente con la enzima Msp I obteniendo 3 bandas de ~ 410, 390 y 120 pb en aislamientos de P. nicotianae, mientras que se produjeron tres bandas de ~ 400, 300 y 200 pb en P. infestans. Sin embargo, no fue posible encontrar diferencias entre aislamientos de una misma especie.
]]> El hecho de encontrar en el presente estudio casi un 50% del total de los aislamientos para el patrón de 1 banda con 400pb y otro 50% para el patrón de las bandas 300pb, coincide con la escasa o nula variabilidad encontrada por los autores antes mencionados al comparar aislamientos entre sí, procedentes de una misma especie.Otra posibilidad para profundizar en el análisis de las diferencias basadas en las regiones ITS es la secuenciación de los fragmentos amplificados, aunque para el género en estudio hasta el momento no se han obtenido resultados muy alentadores para los estudios de variabilidad intraespecífica.
En el caso de la técnica RAPD, de los 40 cebadores empleados en el análisis, solo resultaron funcionales 14 cebadores. El promedio de loci detectado por cebador fue muy variable, desde 4 loci hasta 12. Se amplificaron un total de 104. De ellos resultaron polimórficos los 104 loci detectados, para un 100% de polimorfismo. Los cebadores OPA 10, OPA 02, OPA 18, OPF 04 y OPF 12 se destacaron por el número de bandas amplificadas.
Los resultados que se observan en el dendrograma obtenido a partir del análisis de los datos moleculares de los RAPDs (Fig. 2), destacan la gran variabilidad existente entre los aislamientos estudiados.
En el dendrograma se muestran tres grupos, cuando se estableció como límite para el corte un 70% de similitud, y dentro de estos se observa una gran variabilidad para la especie analizada, lo cual corrobora los resultados obtenidos en los análisis culturales y patogénicos realizados previamente a este trabajo.
Si interesante resulta conocer la variabilidad dentro de la especie, la posible correlación de la distribución obtenida a partir del análisis de los datos moleculares con otras características de los aislados analizados, puede tener un valor práctico importante. Algunas de las características con las cuales se tratan de correlacionar los resultados moleculares son la patogenicidad, la procedencia de los aislamientos u origen geográfico y el hospedante de donde fueron aislados. En este caso se incluyó en el análisis la respuesta al metalaxyl aún cuando este ejemplo de correlación no se encuentra informado en la literatura para P. nicotianae, aunque sí para otras especies de Phytophthora. Los resultados del análisis de correspondencia muestran que, no hay relación entre la agrupación obtenida a través del análisis de los datos moleculares con los ordenamientos obtenidos para las variables de grupos patogénicos, hospedante, origen ni respuesta al metalaxyl (Tabla 2).
Resultados similares se encontraron cuando estos mismos análisis se efectuaron para los dos juegos de cebadores por separado, especialmente para el juego de cebadores OPA . Aún cuando la tendencia se mantiene para el OPF, se pudiera llamar la atención sobre la correspondencia obtenida para la patogenicidad y el cebador OPF 3 (Tabla 3).
La prueba de Chi cuadrado de independencia mostró que cuando la banda de elevado PM (más de 1000pb), está presente, hay un 66,67% de probabilidad de que el aislado en cuestión pertenezca a la raza 1.
Aún cuando los valores son bajos sería interesante profundizar en este estudio con el cebador OPF 03, analizando un mayor número de aislados de ambas razas y procediendo a la secuenciación del fragmento amplificado para su comparación con otras secuencias para Phytophthora en bases de datos.
Muchos autores han arribado a resultados similares. Los análisis de correlaciones considerando los grupos genéticos en P. capsici obtenidos a partir de los datos moleculares con la virulencia y los diferentes comportamientos al metalaxyl, demostraron la no correlación entre estas variables (16). De igual forma, la variabilidad genética por RADP y los estudios de patogenicidad en P. nicotianae en diferentes regiones de China demostraron una alta variabilidad, aunque los marcadores polimórficos no mostraron asociación con la variación patogénica. Zhang et al. (17), a partir del empleo de esta técnica, demostraron que poblaciones de P. nicotianae provenientes de diferentes regiones en China, fueron variables patogénica y genéticamente y le atribuyen a esto los grandes problemas en la obtención de cultivares resistentes para el control de la enfermedad pata prieta.
]]> La no correlación del agrupamiento basado en los datos moleculares con el resto de las variables estudiadas puede deberse a la naturaleza de este marcador. En el caso de los RAPDs, se muestrean zonas del genoma al azar y por lo tanto pueden o no estar relacionadas con los parámetros antes estudiados. Las zonas que no se corresponden en homología con la secuencia del cebador empleado, quedan sin testar. Es por eso que puede recomendarse ampliar el número de cebadores a emplear o incluso probar con cebadores específicos u otro tipo de marcador, que pudiera estar más estrechamente relacionado con el proceso de patogenicidad.Los resultados obtenidos en este trabajo con la técnica de RAPD pusieron de manifiesto la variabilidad molecular que existe en los aislamientos cubanos de P. nicotianae, aun cuando los grupos establecidos no se correlacionaron con los agrupamientos obtenidos para el resto de los parámetros analizados como patogenicidad, hospedante u origen geográfico.
La amplificación de los ITS puede constituir una herramienta útil para el diagnóstico de la especie dada la estabilidad de la técnica y la obtención de un patrón único, aunque ni esta técnica ni la digestión de estos fragmentos fueron útiles en la diferenciación de aislamientos de una misma especie.
Estos resultados constituyen los primeros informes de variabilidad molecular de P. nicotianae en Cuba e indican la necesidad de realizar estudios más profundos a nivel de secuencia de ADN o la comparación de estos datos con otros tipos de marcadores moleculares, a fin de lograr diferenciación intraespecífica y/o la correlación de los mismos especialmente con la patogenicidad de los aislamientos.
REFERENCIAS
1. Kamoun S. Molecular Genetics of Pathogenic Oomycetes. Eukaryotic Cell. 2003;2(2):191-9.
2. Suvillan MJ. Characterization and management of the race structure of Phytophthora parasitica var. nicotianae. Thesis Ph. D. Phytopathology Department Plant Pathology, North Carolina State University, 148 pags, 2004.
3. Ristaino JB, Madritch M, Trout CL, Parr GPCR. Amplification of Ribosomal DNA for Species Identification in the Plant Pathogen Genus Phytophthora. Applied and Environmental Microbiology. 1998;64(3):948-954.
4. Forster H, Cummings M, Coffey M. Phylogenetic relationships of Phytophthora species based on ribosomal ITS I DNA sequence analysis with emphasis on Waterhouse groups V and VI. Mycol Res. 2000;104:1055-1061.
5. Martin RR, James D, Lévesque CA. Impacts of molecular diagnostic technologies on plant disease management. Ann Rev Phytophatol. 2000;38:207-239.
6. Dellaporta SL, Wood J, Hichs JB. A plant molecular DNA minipreparation, versión II. Plant Mol Biol. 1983;1:19-21.
7. Sambrook J, Fritsch E, Maniatis T. Molecular Cloning. A Laboratory Manual, Second Edition. Cold Spring Harbor Laboratory Press 1989.
8. Nei M, Li WH. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc Natl Acad Sci. 1979;76:5269-5273.
9. Bonants PJM, Hagenaar-de Weerdt M, van Gent-Pelzer MPE, Lacourt I, Cooke DEL, Duncan JM. Detection and identification of Phytophthora fragariae Hickman by the polymerase chain reaction. Eur J Plant Pathol. 1997;103:345-355.
10.Johri JK, Singh HB, Srivastava M. Advances in Plant Disease Management. Udit Narayan, Kumud Kumar y Mukesh Srivastava, Editores, Advance Publishing Concepts, New Delhi, 2000. pp.175-183.
11.Lee SB, Taylor JW. Phylogeny of five fungus-like protoctistan Phytophthora species, inferred from the internal transcribed spacers of ribosomal DNA. Mol Biol Evol. 1992;9(4):636-653.
12.Tooley PW, Bunyard BA, Carras MM, Hatziloukas E. Development of PCR primers from internal transcribed spacer region 2 for detection of Phytophthora species infecting potatoes. Appl Environ Microbiol. 1997;63:1467-1475.
13.Martín FN, Tooley PW. Phylogenetics relationships of Phytophthora ramonum, P. nemorosa and P.
pseudosyringae, three species recovered from areas in California with sudden oak death. Mycol Res. 2003;107:1379-1391.
14.Dias D, Machado MA, Penteado ML, Tragón N, Furtado E. Genotypic diversity among P. parasitica isolates revealed by ITS-5.8S rDNA nucleotide sequences. Summa Phytopathol. 2006;32: 43-48.
15.Tripathi A, Singh R, Raj SK, Singh AP, Johri JK. Molecular identification of Phytophthora nicotianae isolates causing leaf rot of betelvine (Piper betle L.). Current Science. 2003;84(1):22-24.
16.Silvar C, Díaz J, Merino F. Real-Time Polymerase Chain Reaction quantification of Phytophthora capsici in different pepper genotypes. Phytopathology. 2005;95:1423-1429.
17.Zhang XG, Sun WX, Guo L, Yu JF, Chang J. Genetic and pathogenic variation among tobacco black shank strains of Phytophthora parasitica var. nicotianae from main tobacco growing in China. J Phytopathol. 2003;151(5):259-266.
(Recibido 18-4-2008; Aceptado 1-9-2008)
]]>{kind=link}
{kind=link}