{kind=link}

Expresión de cuatro genes de defensa en dos variedades de tabaco (Nicotiana tabacum L.) frente al virus del mosaico del tabaco (TMV)
Expression of four defenSe genes against tobacco mosaic virus (TMV) in two varieties of tobacco (Nicotiana tabacum L.)
Sandra Pérez*, D. Cabezas*, O. Chacón**, Evelyn Valera*, O. Borras*** y Ondina León****
*Departamento Biología Sanidad. Facultad de Agronomía. Universidad Agraria de La Habana "Fructuoso Rodríguez Pérez" (UNAH). Carretera de Tapaste y Autopista Nacional. San José de las Lajas, La Hababa, Cuba. Correo electrónico: sandra@isch.edu.cu; ** Instituto de Investigaciones del Tabaco. San Antonio de los Baños, La Habana, Cuba; ***Centro de Ingeniería Genética y Biotecnología (CIGB). Ciudad de La Habana, Cuba; ****Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Hababa, Cuba.
]]>
RESUMEN
El virus del mosaico del tabaco (Tobacco Mosaic Virus, TMV) se considera económicamente importante en el mundo. El presente trabajo se realizó con el objetivo de evaluar la expresión de secuencias representativas del genoma de N. tabacum, en dos variedades del germoplasma de tabaco cubano (BHmN y Corojo) con comportamientos contrastantes en la interacción con el TMV. Las muestras de hojas se tomaron a las 0, 48 horas y siete días después de la inoculación para proceder a la extracción del ARN total según el método del Trizol. Los cebadores se diseñaron utilizando secuencias de genes relacionados con la defensa de las plantas frente al ataque del TMV para analizar la expresión de los mismos a través de la RT-PCR. Como resultado se obtuvo que a los siete días, posterior a la inoculación se observa una expresión de la b 1,3 glucanasa en ambas variedades siendo mayor en la resistente. La quitinasa se induce en la variedad resistente a las 48 h pero a los siete días ya no hay expresión. La proteína fosfatasa serina-treonina y la de choque térmico (HSP) se expresan en ambas variedades en los diferentes tiempos probados aunque la expresión es ligeramente mayor en la variedad BHmN. Se observó en este estudio la inducción de la b 1,3 glucanasa en plantas de N. tabacum inoculadas con el TMV asociada posiblemente a mecanismos defensivos como la respuesta hipersensible y la resistencia sistémica adquirida.
(Palabras clave: virus del mosaico del tabaco; secuencias expresadas; tabaco)
ABSTRACT
Tobacco Mosaic Virus (TMV) is an economically important disease infecting crops worldwide. The objective of the present work was to evaluate the expression of representative sequences from N. tabacum genome in two Cuban tobacco varieties (BHmN and Corojo) with a different behavior in the pathogenic interaction. Leaf samples were taken at 0, 48 hours and seven days after virus inoculation to proceed with the total RNA extraction (Trizol method) and the primers were designed using sequences of tobacco defense genes to analyse their expression by RT-PCR. At seven days after inoculation, a b 1,3 glucanase expression was observed in both varieties, which was higher in the resistant one. Chitinase was induced at 48 h in the resistant variety with no expression seven days after inoculation. Phosphatase serine-threonine and the heat shock protein were expressed in both varieties at all the time tested; however the expression was a little higher in BHmN variety. In this study, the induction of b 1,3 glucanase in N. tabacum plants inoculated with TMV was likely to be related to the hypersensible response and the acquired systemic resistance.
(Key words: tobacco mosaic virus; expressed sequences; tobacco)
]]>
(Recibido 22-9-2008; Aceptado 8-12-2008)
INTRODUCCIÓN
El tabaco (Nicotiana tabacum L.) es uno de los productos de mayor demanda en el mundo y es objeto de gran intercambio comercial. En Cuba, la producción tabacalera ocupa un lugar cimero en la economía por ser una de las principales fuentes de ingreso (1).
Este cultivo es afectado por múltiples enfermedades, entre las que se incluyen el moho azul (Peronospora hyoscyami F. sp. tabacina), la pata prieta (Phytophthora nicotianae Breda de Haan) y el mosaico del tabaco. El uso de variedades resistentes se considera una de las tácticas fundamentales para el control de estas enfermedades y por ende los programas de mejoramiento en nuestro país a partir del cruzamiento genealógico o el cultivo de anteras persiguen este fin (2).
El virus del mosaico del tabaco (Tobacco Mosaic Virus, TMV) tiene una amplia distribución en el mundo y se considera económicamente importante. Infecta a más de 199 especies diferentes de plantas pertenecientes a 30 familias (3). En Cuba se detectó a mediados del siglo pasado y provocó importantes pérdidas (más del 50% de la producción) en las provincias de La Habana y Pinar del Río (4).
La comprensión de los procesos fisiopatológicos en la interacción de las plantas con los virus continúa siendo un reto, pues no está esclarecido totalmente como la infección viral influye en la bioquímica y la fisiología de las células y tejidos de las mismas. Un mejor entendimiento de la expresión de los genes de las plantas hospedantes asociados a la interacción de estas con los virus podrá revelar detalles significativos de los mecanismos de respuesta a estos patógenos (5).
Por todo lo planteado se propone como objetivo de este trabajo evaluar la expresión de secuencias representativas del genoma de N. tabacum con el proceso defensivo en genotipos de tabaco (N. tabacum) resistentes y susceptibles al virus del mosaico del tabaco.
]]>
MATERIALES Y MÉTODOS
Para evaluar la expresión de proteínas relacionadas con la defensa, se sembraron semillas de dos variedades cubanas de tabaco (N. tabacum), una resistente al TMV, la BHmN (variedad Burley)(6) y otra susceptible, la Corojo (variedad de tabaco Negro) (7), en una casa de cristal, del Centro Nacional de Sanidad Agropecuaria (CENSA), siguiendo las instrucciones del Instructivo Técnico del Tabaco (8).
Inoculación del TMV
La inoculación se realizó con un aislamiento del TMV procedente de la variedad Corojo sembrada en el Instituto de Investigaciones del Tabaco (IIT). Las hojas infectadas con TMV se maceraron en solución Fosfato de sodio 0,02 M (pH 6) con una proporción de 1:2 (por cada gramo de tejido infectado se aplican 2 mg.mL-1 de solución) y luego la suspensión fue aplicada sobre la superficie de las hojas intermedias (tres hojas como mínimo) de las variedades seleccionadas. Las hojas inoculadas se habían frotado previamente con carborundum. La inoculación se realizó a una temperatura promedio de 25oC y las plantas tenían 30 días.
Las muestras de hojas se tomaron a las 0 h, 48 h y siete días después de la inoculación. Se colectaron tres muestras y tres testigos por cada una de las variedades para proceder a la extracción del ARN total utilizando 1,3 mL de Trizol por cada 100 mg de tejido (9).
Síntesis de la primera cadena de ADNc
La primera cadena del ADNc se sintetizó utilizando el juego de transcripción reversa (10), para lo cual se preparó una mezcla que contenía 1,0 mg.mL-1 de ARN total, solución de la primera cadena 10x 25mM de MgCl2, 10 mM de dNTPs, 40 unidades.mL-1 de RNasin Inh, 0,5 mg.mL-1 de oligo(dT), 25 unidades.mL-1 de la transcriptasa reversa AMV y 4,5 mL de agua desionizada. El programa GeneAmp 9700 (Applied Biosystems, Foster City, CA) se utilizó con un paso de 10 min a 70oC y otro de dos horas a 42oC para la síntesis de la segunda cadena.
Análisis de los niveles de expresión de los genes seleccionados por RT-PCR
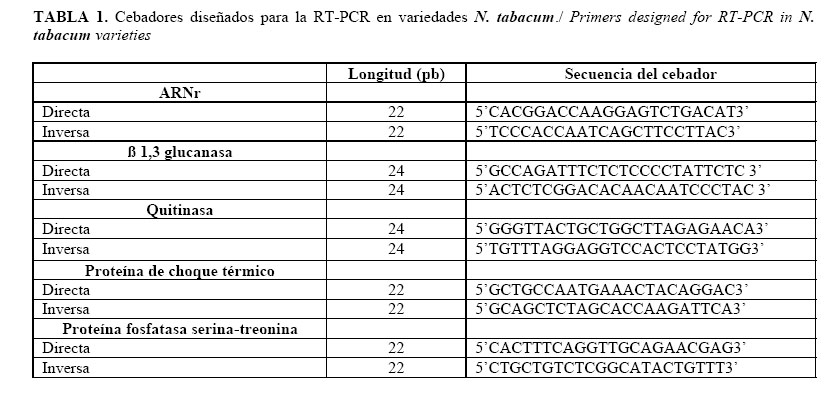
Los cebadores se diseñaron mediante el programa Primer Express 2.00 (Applied Biosystems Software) utilizando cinco secuencias únicas provenientes de una genoteca de ADNc de N. tabacum (11) (Tabla 1). ]]>
Para la RT-PCR se preparó una mezcla que contenía 1 mg.mL-1 de ADNc, solución de la primera cadena 10x, 25mM de MgCl2, 10 mM de dNTPs, 5 unidades.mL-1 de Taq polimerasa y 21,5 mL-1 de agua desionizada. La concentración de cada cebador (directo e inverso) en la mezcla fue de 2 mM. Se realizaron tres repeticiones por cada gen.El programa de amplificación incluyó un ciclo de cuatro min. a 94oC, 30 ciclos de 30 seg. a 94oC, 30 seg. a 58oC, 1,5 min. a 72oC y por último un ciclo de cuatro min. a 72oC. Los resultados fueron visualizados en geles de agarosa al 2% teñidos con bromuro de etidio.
RSULTADOS Y DISCUSIÓN
La inoculación con TMV de dos variedades cubanas de tabaco dio como resultado a las 48 horas lesiones relacionadas con la respuesta hipersensible (Hypersensible Reaction, HR) en la variedad resistente como se observa en la Figura 1.
La reacción incompatible producida en la interacción N. tabacum variedad BHmN-TMV provocó lesiones necróticas que se intensificaron a partir de los siete días posterior a la inoculación. Este resultado se corresponde con los experimentos realizados por Levy et al. (12) donde la infección producida por el TMV en plantas de tabaco que tienen el gen N activaron la HR entre las 40 y las 48 horas después de la inoculación, y las partículas del virus quedaron restringidas inmediatamente en la región rodeada por las lesiones necróticas.
Para analizar la expresión de los genes relacionados con la defensa se realizó la RT-PCR. Los resultados de esta expresión se muestran en la Figura 2. El transcripto que codifica para la b 1,3 glucanasa no se induce a las 48 horas en las dos variedades estudiadas. Sin embargo, a los siete días se observa la expresión tanto en la variedad susceptible como en la resistente, aunque en esta última es mayor. Este resultado podría explicarse porque al aumentar la severidad de las lesiones necróticas los niveles de ácido salicílico se incrementan y esto se relaciona con la mayor expresión de esta proteína pues este compuesto es un inductor de las PR-2. La síntesis de ácido salicílico aumenta después de la HR y esto conlleva a la activación de la Resistencia Sistémica Adquirida (SAR, Systemic Adquirid Resistense) a través de toda la planta, mecanismo que activa un conjunto de genes entre los que se encuentran las PR-2 (13).
 ]]>
]]>
Otros autores, entre ellos Tiryaki y Tunaz (15), plantearon que las PRs se asocian con la iniciación de la SAR. Estas proteínas según van Loon et al. (16) se acumulan después del ataque del patógeno y fundamentalmente las PR-1, PR-2 y PR-5 se relacionan con la respuesta de la SAR correspondiéndose este planteamiento con los resultados de este estudio donde la PR-2 se indujo a los siete días después de la inoculación.
Las investigaciones sobre las b 1,3 glucanasas en la interacción planta-virus son limitadas, por lo que este trabajo es una contribución al conocimiento de las funciones de esta PR en dichas respuestas de defensa.
La quitinasa, se expresa levemente en la variedad resistente a las 48 h, pero a los siete días ya no hay expresión de la misma. Este resultado puede estar indicando su vinculación con la muerte celular, resultado que ha sido informado en esta interacción, para esta proteína y en este tiempo (48 h) (19, 20).
La leve expresión de la quitinasa en la variedad susceptible a las 48 horas puede deberse a su función en la movilización de nutrientes hacia los tejidos dañados por el virus o en la protección contra los daños celulares provocados por el virus (21). A las 48 horas el virus se está moviendo célula-célula por lo que su síntesis se ha completado encontrándose ya en el sistema vascular de la planta.
Elvira et al. (22) en plantas de Capsicum chinense L. infectadas con el virus del moteado del pepino encontraron acumulación tanto de la b 1,3 glucanasa como de las quitinasas en la interacción compatible, lo que indicó una diferencia en la regulación de la expresión debido a la complejidad de la interacción planta-virus.
Los transcriptos que codifican a la proteína fosfatasa serina-treonina y a la proteína de choque térmico (HSP) se expresan en ambas variedades en los diferentes tiempos probados aunque la expresión es ligeramente mayor en la variedad BHmN. La detección de ambas proteínas incluso en el tiempo 0 provee evidencias de que pueden encontrase de forma basal en N. tabacum, lo cual confirma los análisis realizados por Tyrell et al. (23) con la proteína de choque térmico 101 (HSP101) en plantas de Nicotiana benthamiana infectada con TMV donde la expresión de esta proteína no varió en el tiempo.
En la presente investigación es probable que la función de la proteína fosfatasa serina-treonina, al igual que la de choque térmico sea fundamentalmente de señalización y que los genes de resistencia estudiados codifiquen receptores que interactúen con elicitores específicos (como el ácido salicílico) y cuando la señal se percibe por esta y otras proteínas presentes en la planta, los productos de los genes R activen las respuestas de defensa.
En lo referente al transcripto control ARNr 18S mostró similar señal en todos los tiempos y variedades probadas. En las infecciones virales la función biológica que pudiera realizar esta enzima en la respuesta de defensa de N. tabacum frente al TMV aún no esta esclarecida. Algunos autores como Boevink y Oparka (17) consideran que después de los seis días debe comenzar el bloqueo o sellado de los plasmodesmos para evitar que el virus ingrese al sistema vascular y colonice exitosamente la planta. En este momento la inducción de las glucanasas aumenta, pues la acumulación de calosa debe incrementarse para actuar como una barrera protectora y sellar los plasmodesmos y de esta forma limitar el movimiento célula-célula del virus.
La expresión diferencial de la b 1,3 glucanasa tanto en la variedad resistente como en la susceptible a los siete días posterior a la inoculación es similar a los resultados obtenidos por López (18) quién observó inducción de PRs tanto en la variedad susceptible como en la resistente de Saccharum spp. inoculadas con Puccinia melanocephala H & P. Sydow y afirmó que la acumulación de esta proteína no está restringida a las plantas resistentes. Sin embargo, el tiempo de aparición y la magnitud de la inducción puede diferir influyendo en la reacción de defensa. ]]>
Los estudios sobre el genoma del TMV y el uso de los virus como herramientas moleculares, representan vías prometedoras y poderosas de investigación. El presente trabajo profundiza en los conocimientos fisiopatológicos y los resultados obtenidos brindan algunos elementos para caracterizar el germoplasma cubano de tabaco en cuanto a la genotipificación y expresión de genes de defensa ante la infección viral, que pudieran emplearse en los programas de selección asistida por marcadores y en el aislamiento de nuevos genes de interés.
REFERENCIAS
1. Basulto O. Cuba acapara 68% del mercado mundial de tabacos Premium. Granma Internacional, 7 de Septiembre; 2005.
2. García CH. Técnicas especiales de mejoramiento genético en el tabaco (N. tabacum). Cubatabaco. 2003;4(2):75-79.
3. Bagley A. Controlling tobacco mosaic virus in tobacco through resistance; 2001. Master's Thesis. Virginia Polytechnic Institute and State University, Blacksburg, VA (Consultada : 28 jun 2005). Disponible en: http://scholar.lib.vt.edu/ theses/available/etd-01122002-113631/.
4. Espino E, Torrecilla G. El tabaco cubano. Recursos fitogenéticos. Editorial Científico-Técnica, Instituto Cubano del Libro; 1999. p. 39-40.
5. Collazo C, Ramos PL, Chacón O, Borroto CJ, López Y, Pujol M, Toma BP, Hein I, Borrás-Hidalgo O. Phenotypical and molecular characterization of the Tomato Motel Taino virus-Nicotiana megalosiphon interaction. Physiological and Molecular Plant Pathology. 2006;67:231-236.
6. García G, García V, Pino LA, Espino E. `Burley Habana-13' (`BH-13') primera variedad comercial de tabaco obtenida en Cuba por cultivo de anteras con resistencia al moho azul (Peronospora tabacina), pata prieta (Phytophthora parasitica) y al virus del mosaico del tabaco (TMV). Cubatabaco. 1999;1(1):49-54.
7. MINAG (Ministerio de la Agricultura). Instructivo técnico para el cultivo del tabaco. Sedagri/Agrinfor, Ciudad de La Habana; 1998.
8. Torrecilla G, Peñalver N, Gil M, García M, García V, Pino LA, et al. Screening en variedades del banco de germoplasma de tabaco para la búsqueda de fuentes de resistencia a enfermedades. Cubatabaco. 2001;2(2):58-64.
9. Chomczynski P, Sacchi N. Single Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Analytical Biochemistry. 1987; 162.
10.Manual Técnico. Universal RiboClone® cDNA Synthesis System". 2005. Printed in USA. P.13, Promega Corporation. Disponible en: http//www.promega.com.
11.Pérez SA, Cabezas DM, Haitao D. Large-scale identification of ESTs from Nicotiana tabacum by normalized cDNA library sequencing. Contributions to Tobacco Research. 2006;22(2):114-124.
12.Levy M, Edelbaum O, Sela O. Tobacco mosaic virus regulates the expression of its own resistance gene N. Plant Physiology. 2004;135(4):2392-2397.
13.Murphy A, Carr JP. Salicylic Acid Has Cell-Specific Effects on Tobacco mosaic virus Replication and Cell-to-Cell Movement. Plant Physiology. 2002;128:552-563.
14.Verberne MC, Hockstra J, Bol JF, Linthorst JM. Signaling of systemic acquired resistance in tobacco depends on ethylene perception. The Plant Journal. 2003;35:27-32.
15.Tiryaki S, Tunaz H. Systemic acquired resistance: Characterization of genes associated with plant defense response. J Cell and Molecular Biology. 2004;3:9-14.
16.van Loon L, Rep M, Pieterse C. Significance of Inducible Defense-related Proteins in Infected Plants. Ann Rev Phytopathology. 2006;44(4):150-162.
17.Boevink P, Oparka KJ. Virus-Host Interactions during Movement Processes. Plant Physiology. 2005;138:1815-1821.
18.López RL. Bases bioquímicas-moleculares de la respuesta de defensa de la caña de azúcar (Saccharum spp.) a Puccinia melanocephala H & P. Sydow. Tesis para optar por el grado de Doctor en Ciencias Agrícolas. Centro Nacional de Sanidad Agropecuaria (CENSA). La Habana Cuba; 2002.
19.Fritig B, Kauffmann S, Dumas B, Geoffroy P, Kopp M, Legrand M. Mechanism of the Hypersensitivity Reaction of Plants. In: Proceeding of Ciba Foundation Symposium-Plant Resistance to Virus. Online, ISBN: 9780470513569. 2007.
20.Riviére MP, Marais A, Ponchet M, Willats W, Galiana E. Silencing of acidic pathogenesis-related PR-1 genes increases extracellular b 1,3 glucanase activity at the onset of tobacco defense reactions. J Exp. Botany, 2008;1:15.
21.Espinoza C, Medina C, Somerville S, Arce-Johnson P. Senescence-associated genes induced during compatible viral interactions with grapevine and Arabidopsis. J Exp Botany. 2007; 58: 3197-3212.
22.Elvira MI, Galdeano MM, Gilardi P, García-Luque I, Serra MT. Proteomic analysis of pathogenesis-related proteins (PRs) induced by compatible and incompatible interactions of pepper mild mottle virus (PMMoV) in Capsicum chinense L. plants. J Exp Botany. 2008;59(6):1253-1265.
23.Tyrell C, Wang Y, Huang Z, Joanne MY, Jian-Bing F, Steven AW. Tobamovirus infection is independent of HSP101 mRNA induction and protein expression. Virus Research. 2006;121(1):33-41. ]]>