
INDUCTION OF EXTRACELLULAR ENZYMES WITH Meloidogyne incognita AND Globodera pallida EGGS
Belkis Peteira*, Ivania Estévez**, S. Atkins**, L. Hidalgo-Díaz*** y B. Kerry**
Grupos de Fitopatología* y Plagas Agrícolas***, División de Protección de Plantas, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, La Habana, Cuba. Correo electrónico: bpeteira@censa.edu.cu; **Nematode Interactions Unit, Rothamsted Research, Harpenden, Herts AL5 2JQ, UK
RESUMEN
La cepa cubana IMI SD 187 del hongo nematofago Pochonia chlamydosporia var. catenulata es un agente de control biológico potencial de nematodos formadores de agallas. Existen informes sobre la acción controladora de los hongos de la especie Pochonia chlamydosporia sobre nematodos de quistes. Sin embargo también se conoce de la especificidad de los aislamientos según su hospedante original. El objetivo de este trabajo fue estudiar el comportamiento de diferentes sistemas enzimáticos en la cepa IMI SD 187, frente a huevos de Meloidogyne incognita y Globodera pallida. Se realizaron los ensayos de inducción con huevos de ambos nematodos en medio líquido y se determinaron los contenidos de proteínas totales y niveles de actividad enzimática de proteasas, quitinasas, lipasas y VCP 1. La dinámica de la inducción de los sistemas enzimáticos estuvo relacionada con las fases del proceso de infección de los huevos de M. incognita en la cepa IMI SD 187. Esta cepa de P. chlamydosporia var. catenulata procedente de nematodos formadores de agallas es capaz de infectar huevos de nematodos de quistes.
Palabras clave: Pochonia chlamydosporia var. catenulata; actividad enzimática; Meloidogyne incognita; Globodera pallida
The Cuban strain IMI SD 187 of the nematophagous fungus Pochonia chlamydosporia var. catenulata is a potential biological control agent for the root knot nematodes. There are reports about the controlling action of these fungi on cyst nematodes. However, isolate specificity according to the original host is also known. The aim of this work was to study the enzymatic performance induced by Meloidogyne incognita and Globodera pallida eggs. The induction assays were carried out in liquid media. The total protein content and the enzymatic activity levels for proteases, chitinases, lipases and VCP 1 were determined. The time course experiment for enzymatic activity induction was related to the infection process of M. incognita eggs by the strain IMI SD 187. The strain IMI SD 187 of P. chlamydosporia var. catenulata isolated from root knot nematodes is able to infect eggs from cyst nematodes.
Key words: Pochonia chlamydosporia var. catenulata; enzymatic activity; Meloidogyne incognita; Globodera pallida
(Recibido 10-2-2009; Aceptado 24-6-2009)
]]>
INTRODUCCIÓN
Pochonia chlamydosporia var. catenulata (Kamyscho ex Barron y Onions) Zare y Gams, en especial la cepa IMI SD: 187, ha mostrado ser un agente de control biológico potencial de nematodos formadores de agallas en Cuba (1). Los estudios realizados con esta cepa han demostrado su robustez, elevada producción de clamidosporas durante el proceso de producción masiva por fermentación sólida, con más de un 93% de viabilidad y valores de colonización de masas de huevos de Meyloidogyne incognita (Koffoi y White) Chitwood y parasitismo de sus huevos de 98 y 70%, respectivamente (2). Algunos informes también clasifican a P. chlamydosporia como agente de control para nematodos formadores de quistes (2).
La especificidad por el hospedante es un aspecto importante a tener en cuenta en la selección de una cepa para su producción e introducción como agente de control biológico (ACB) o la aplicación de una combinación de cepas para el control de un nematodo en particular o mezclas de diferentes especies o géneros, en un agroecosistema dado. Esta especificidad pudiera deducirse a través de la expresión de diferentes sistemas enzimáticos relacionados con el proceso de parasitismo de los huevos o mediante la búsqueda de algún marcador de ADN relacionado con este aspecto.
Pochonia chlamydosporia no escapa a esta aseveración. En un estudio de inducción de proteínas extracelulares, Segers (3), encontró que los huevos de Globodera rostochiensis Woll tuvieron mucho menor efecto en la inducción de VCP1 que los huevos del hospedante susceptible, M. incognita. Este autor argumenta que aunque los huevos de los quistes no fueron liberados artificialmente durante la preparación del medio, los quistes maduros no estaban sellados y era de esperar que el hongo tuviera acceso relativamente fácil a través de las aberturas naturales del quiste.
Teniendo en cuenta el conocimiento que se tiene acerca del proceso de parasitismo y la importancia de la proteasa VCP1, así como el funcionamiento de otras enzimas de carácter hidrolítico, el objetivo de este trabajo fue el estudio del comportamiento de diferentes sistemas enzimáticos, inducidos en tres cepas de P. chlamydosporia, por huevos de dos hospedantes diferentes: M. incognita y Globodera pallida (Stone) Mulvey y Stone.
MATERIALES Y MÉTODOS
Cepas y medios de inducción: ]]>
Las cepas empleadas en este experimento fueron:1. Cepa 309 (P. chlamydosporia var. chlamydosporia, proveniente de M. incognita).
2. Cepa 280 (P. chlamydosporia var. chlamydosporia, proveniente de G. pallida).
3. Cepa IMI SD 187 (P. chlamydosporia var. catenulata, proveniente de M. incognita)
Las tres cepas procedieron de la colección de Rothamsted, Reino Unido, donde habían sido conservadas en medio agar harina de maíz (AHM) a 4°C
y para su uso se subcultivaron en el mismo medio, durante dos semanas, en oscuridad, a 28ºC.
Se evaluaron tres variantes de medio de cultivo a partir de un medio basal que además fue empleado como control:
a) Medio basal: Constituido por 0,03% NaCl; 0,03% MgSO4 x 7 H2O; 0,03% K2HPO4; 0,02% de extracto de levadura (4).
b) Medio basal suplementado con huevos de M. incognita 0.1% (p/v) (3). Se empleó la raza R2:1135 de M. incognita procedente de América del Norte, mantenida sobre Solanum melangena L. (berenjena) en el banco de especies de la estación de Rothamsted Research, Reino Unido. Las masas de huevos equivalentes a la concentración requerida en el medio se colectaron de raíces de plantas infectadas. Posteriormente se desinfectaron superficialmente con Hipoclorito de Sodio al 10% (a partir de la solución comercial) durante dos minutos y por último se procedió a la desinfección de los huevos siguiendo el protocolo descrito por Manzanilla-López (5). Los huevos desinfectados se suspendieron en agua destilada estéril y se añadieron al medio ya dispensado.
c) Medio basal suplementado con huevos de G. pallida 0.1% (p/v) (3). En este caso los huevos de la raza PA1 de G. pallida se obtuvieron de quistes provenientes de muestras de suelo de un campo infestado en el Reino Unido. Para la desinfección superficial se procedió de forma similar a la detallada anteriormente para M. incognita. ]]>
Los pasos correspondientes a la inoculación de todas las variantes de medio, su incubación, toma de muestra, así como la cuantificación de proteínas totales y de las actividades enzimáticas se desarrollaron según los protocolos descritos por Peteira et al. (6). Para cada tratamiento (Basal, M. incognita y G. pallida) y cada cepa estudiada, se realizaron tres repeticiones y tres réplicas, así como tres réplicas para cada determinación de proteínas totales y actividades enzimáticas. Las muestras se tomaron a los tres, cinco y siete días, después de la inoculación de los medios con el hongo. Además, se colectaron y procesaron como control, muestras de los medios sin inocular con el hongo, para eliminar los posibles valores de proteínas totales de fondo.Además, a los siete días se realizaron observaciones bajo microscopio óptico, para todas las variantes, para lo cual se extrajo en cada caso una muestra de 1mL del medio conteniendo huevos del hospedante. La muestra se depositó en un vidrio reloj de 2cm de diámetro, el cual se colocó bajo el microscopio Olympus (ZEISS), empleando aumentos de 20X y 40X para chequear la infestación de los huevos por el hongo.
El diseño del experimento fue completamente aleatorizado. Todos los datos se compararon a través de un Análisis de Varianza Simple y la Prueba de Rangos Múltiples de Duncan (p = 0,05), usando el programa SAS (7). Se tuvo en cuenta las cepas como variable independiente y se analizaron los indicadores bioquímicos en cada tiempo y en el hospedante específico.
RESULTADOS Y DISCUSIÓN
Las concentraciones de proteínas totales para las tres cepas analizadas en el medio suplementado con huevos de M. incognita aparecen en la Figura 1. Es posible observar como a los tres días, la cepa IMI SD 187 es la de mayor nivel de producción de proteínas con valores significativamente superiores respecto a las otras cepas, que muestran niveles estadísticamente similares para este indicador. Al quinto día y séptimo día, la cepa 309 se separa del comportamiento del resto, y alcanza niveles significativamente superiores de síntesis de proteínas al séptimo día. En el séptimo día es posible diferenciar las tres cepas por sus niveles de síntesis, que son estadísticamente diferentes y mayores en las cepas provenientes del hospedante M. incognita, usado en este experimento como agente inductor.
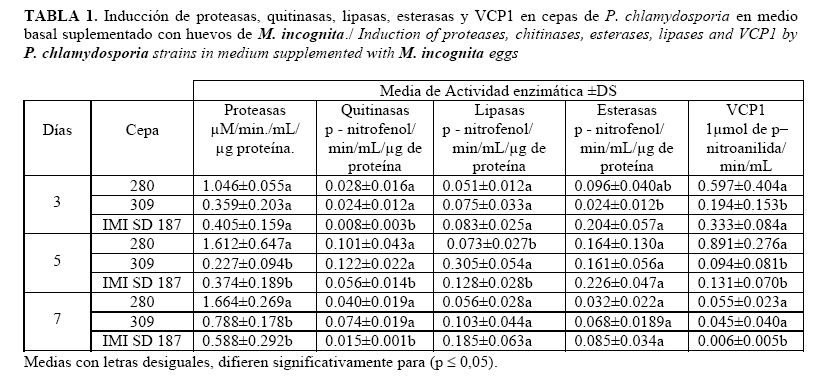
En la Tabla 1 se observa el comportamiento de las actividades enzimáticas para el medio suplementado con huevos de M. incognita. La cepa 280 se caracterizó por presentar los mayores niveles de actividad de proteasa al compararla con las otras cepas, durante todo el tiempo analizado, aunque al tercer día no presenta diferencias significativas con el resto de las cepas. Las cepas IMI SD 187 y 309 (provenientes de M. incognita) presentaron valores inferiores, aunque no difieren significativamente entre ellas.
Las actividades de quitinasas se mantienen con valores similares para las cepas 280 y 309, mientras la cepa IMI SD 187 alcanza los menores valores para esta actividad enzimática. Sin embargo, la cepa más productora de esterasas es la cepa IMI SD 187, aunque sin diferencias significativas con las cepas restantes, excepto al tercer día, donde la producción de esta enzima es significativamente superior a la de la cepa 309. En las lipasas, no se observa ningún comportamiento notable que diferencie estadísticamente a las cepas estudiadas, solo al quinto día se destaca la cepa 309 con valores significativamente superiores a los mostrados por las otras dos cepas en estudio. Es de notar que la enzima VCP1 fue sintetizada en mayores concentraciones por la cepa proveniente de nematodos formadores de quistes (cepa 280). Las diferencias de esta cepa con respecto a las otras se hicieron más notables al quinto día de análisis, donde alcanzó valores significativamente superiores con diferencias significativas respecto a los obtenidos por las cepas provenientes de nematodos formadores de agallas. ]]>
Es posible que las diferencias en la composición y el grosor de las láminas que forman la cubierta protectora de los huevos de M. incognita y de G. pallida, traigan como consecuencia diferencias en los patrones de síntesis de las enzimas necesarias para degradarlas. Por eso, es de suponer que la cepa proveniente de G. pallida (un hospedante más resistente) (3), esté mejor adaptada para llegar al interior del huevo y para esto debe contar con enzimas del mismo tipo, pero más específicas, potentes o nuevas isoenzimas diferentes de aquellas con las que cuentan las cepas provenientes de M. incognita, el cual parece ser un hospedante más susceptible al ataque de Pochonia.Cuando las cepas se incubaron en medio suplementado con huevos del nematodo formador de quistes, G. pallida, se encontró que prácticamente en toda la dinámica estudiada, los valores de proteínas totales para la cepa 280, proveniente de este hospedante, fueron significativamente mayores, al compararlos con las otras cepas (Fig. 2) excepto a los siete días, cuando no difiere de la cepa IMI SD 187.

Los mayores valores de proteínas totales se alcanzaron a los tres días de incubación y disminuyen en el tiempo a partir de este momento. Es posible que para la cepa proveniente de este hospedante (cepa 280), los procesos bioquímicos relacionados con la infección de los huevos se han desarrollado de manera más rápida en las primeras horas de la inoculación y ya a partir del tercer día comienza a disminuir la síntesis de proteínas. Para las otras cepas provenientes de un hospedante diferente, que no mostraron niveles semejantes de síntesis, pudiera significar
que los huevos de G. pallida son difíciles de penetrar con sus enzimas y la síntesis también va en decremento al no constituir un sustrato tan favorable como los huevos de M. incognita.
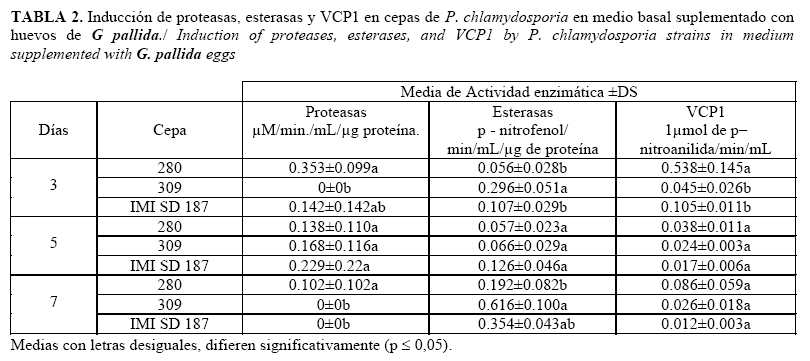
Las actividades enzimáticas de proteasas y VCP1 en este medio mostraron una tendencia similar a la observada en las proteínas totales: niveles máximos de ambas enzimas a los tres días, para luego ir disminuyendo en sus valores de actividad (Tabla 2). A los tres días, la actividad proteasa de la cepa 280 es significativamente superior a la cepa 309 y estadísticamente similar a la cepa IMI SD 187, mientras que para la enzima VCP1 alcanza valores estadísticamente superiores a los de las cepas restantes. A los cinco días no se encontraron diferencias estadísticas entre las tres cepas. Sin embargo, las cepas IMI SD 187 y 309 mostraron mayores niveles de esterasas, especialmente al séptimo día de la dinámica en comparación con la cepa 280, aunque solo la cepa 309 alcanzó valores estadísticamente diferentes. Es de notar que de forma general, los valores de proteasas alcanzados en este medio fueron inferiores a los alcanzados en el medio con huevos de M. incognita. En este medio no hubo inducción de quitinasas ni de lipasas.
Cuando las tres cepas se estudiaron en medio basal (Tabla 3), los valores obtenidos tanto de proteasas como de VCP1 fueron los mayores en comparación con los obtenidos en los otros dos medios. De igual manera, la cepa 280 exhibió los mayores niveles para ambas enzimas.
Por su parte, la cepa IMI SD 187 continúa destacándose por poseer altos niveles de esterasas, con relación a las otras cepas.
El medio suplementado con huevos de M. incognita, fue más favorable para la síntesis de enzimas en las cepas provenientes de este hospedante que el medio suplementado con huevos de G. pallida, donde solo se produjo la inducción de esterasas, VCP1 y proteínas totales.
Es de destacar que el indicador proteínas totales mostró resultados especialmente interesantes. Para la cepa 280, los mayores valores de proteínas totales se alcanzaron en el medio que contenía a su hospedante originario, seguido del que contenía huevos de M. incognita y por último los valores obtenidos en el medio basal. Para las cepas 309 y IMI SD 187 se mantuvo una tendencia similar: sus niveles de proteínas totales fueron superiores en el medio suplementado con huevos de Meloidogyne, seguidos del medio con huevos de G. pallida y de los alcanzados en medio basal. Este comportamiento pudiera estar relacionado con el hospedante de procedencia de las cepas y tal vez con la agresividad de las mismas. Las lipasas, por su parte, no fueron un buen indicador para diferenciar las cepas entre sí por su comportamiento y en el resto de las enzimas tampoco se encontró un patrón específico. ]]>
Anteriormente, Segers (3) realizó un estudio de inducción en la cepa 10 de P. chlamydosporia donde utilizó huevos de M. incognita (0,125%) y huevos de G. rostochiensis (0,1 y 0,5%). Cuando ambos inductores fueron empleados en concentraciones similares, no se encontró efecto inductor para los huevos del nematodo formador de quistes. Solo se alcanzaron valores medibles para este caso cuando se emplearon las concentraciones más altas (0,5%) y aún así, se encontraron los mayores valores de proteasas en el medio con huevos de M. incognita como inductor, lo cual coincide con los resultados obtenidos en este trabajo.Al analizar cada cepa en el medio inoculado con huevos de su hospedante originario, se observa una relación entre el comportamiento de las enzimas estudiadas y los pasos del mecanismo de infección de los huevos por el ACB, especialmente en la cepa IMI SD 187 (Fig. 3).

En el tercer día de incubación, las proteasas comienzan a declinar en sus valores. Esto se pudiera asociar a que en ese momento la síntesis de estas enzimas no es tan apremiante, pues ya se ha degradado la capa más externa de los huevos, la de naturaleza proteica, que precisamente sirve de sustrato a la acción de estas enzimas. Ellas, con su acción, han dejado al descubierto la capa siguiente, la de quitina. Se observa entonces como del día tercero al quinto hay un incremento en la síntesis de quitinasas y lipasas, enzimas que serán necesarias para la degradación de la tercera capa, la más interna, constituida por lípidos y proteínas. Las quitinasas disminuyen su concentración para el séptimo día, donde las lipasas continúan su acción y además se registra un nuevo aumento de las proteasas, que deben actuar junto con las lipasas para degradar la última capa y el contenido del huevo.
Es conocido que las cubiertas de los huevos de nematodos formadores de agallas y de nematodos formadores de quistes poseen la misma estructura básica. Sin embargo, se plantea que el grosor de las tres láminas que las componen, varía considerablemente entre los diferentes géneros de nematodos. La cubierta de los huevos de Meloidogyne es generalmente más delgada que la de los géneros Heterodera y Globodera y esto debe tenerse en cuenta al hablar de la vulnerabilidad al ataque del ACB. Otro factor importante es que los huevos de Meloidogyne son más susceptibles a la lisis enzimática debido a que es de esta forma que los juveniles escapan del huevo (8, 9).
Hasta el presente, los mecanismos que confieren al hongo especificidad frente al nematodo hospedante no están claros. Diferentes autores encontraron a partir del análisis de diferentes aislamientos por PCR-ERIC, que era posible agrupar los materiales estudiados de acuerdo al hospedante, planteando que esto pudiera indicar que hay elementos en la población de P. chlamydosporia que están asociados con la infección de un género particular de nematodos (10, 11, 12). Morton et al. (9) encontraron polimorfismo en la composición aminoacídica de la VCP1, al comparar aislamientos provenientes de diferentes hospedantes. Este polimorfismo se ubica específicamente en la posición 99 donde la glicina encontrada en las cepas provenientes de nematodos formadores de agallas es sustituida por una alanina en las cepas de nematodos formadores de quistes y ocurre en la región de unión al sustrato, lo que pudiera estar relacionado con un efecto de impedimento estérico a la unión de la enzima con el sustrato. El autor concluye que si este polimorfismo altera la unión de la enzima a la prolina (aminoácido que forma parte de la cubierta de los huevos en casi un 30%), se vería afectado el proceso de degradación de la cubierta del nematodo y que la existencia de especificidad por sustrato en la VCP1, pudiera indicar que la estructura en las proteínas de los huevos de los diferentes géneros de nematodos es diferente, lo cual se relaciona con la expresión de diferentes isoformas de VCP1 por diferentes cepas.
Aún cuando las cepas fueron enfrentadas a diferentes hospedantes y ellas mismas mostraron diferente comportamiento, llama la atención que todas fueron capaces de infectar los huevos de ambos géneros de nematodos, como se observa en las fotos (Fig. 4). Las distintas cepas de P. chlamydosporia forman un enrejado de micelio alrededor de los huevos del nematodo y hay producción de conidios y clamidosporas. Esto último es otro aspecto interesante de este estudio ya que se ha declarado que no hay formación de clamidosporas en medio líquido (13). Sin embargo, se detectó la formación de pequeñas "micelas" compuestas por micelio del hongo y los huevos del nematodo atrapados en él, las que no fueron observadas en los experimentos en los cuales fue empleado el medio básico con diferentes suplementos. Estas "micelas" fueron observadas a simple vista y posteriormente bajo microscopio.

]]>
Por lo tanto, sería interesante tratar de encontrar algún compuesto que simule la composición química de los huevos, especialmente alguna proteína que pudiera ser empleada en la producción masiva en medio líquido, como inductor de la formación de las clamidosporas necesarias para la aplicación del hongo en campo. Esto indudablemente resulta beneficioso, pues el proceso de recuperación de las clamidosporas sería más sencillo en un medio líquido a través de un proceso de filtración, que como se hace en la actualidad para la separación de las esporas de la fase sólida.
Los resultados obtenidos en este trabajo confirman los obtenidos por otros autores que han mostrado un comportamiento diferencial de las cepas provenientes de diferentes nematodos hospedantes. Estos son los primeros estudios que se realizan de esta naturaleza para una cepa de la variedad catenulata en el mundo. Se muestra también por primera vez, que la cepa IMI SD 187 autóctona de Cuba es capaz de infectar huevos de G. pallida, por lo cual puede ser empleada también en el control de este nematodo, teniendo como base una caracterización del comportamiento enzimático de esta cepa en presencia de los dos hospedantes estudiados.
REFERENCIAS
1. Hidalgo-Díaz L, Kerry B. Integration of biological control with other methods of nematode management. In: Ciancio A, Mukerji KG (Eds). Integrate management and biocontrol of vegetable and grain crops nematodes. Serie Kluwer, Springer, pp. 28-43, 2007.
2. Montes de Oca N, Arévalo J, Acosta N, Peteira B, Hidalgo-Díaz L, Kerry B. Estabilidad de la cepa IMI SD 187 de Pochonia chlamydosporia var. catenulata (Kamyscho ex Barron y Onions) Zare y W. Gams. Parte I. Indicadores morfológicos, productivos y patogénicos. Rev Protección Veg 2005;20(2):93-101.
3. Segers R, Butt TM, Kerry BR, Beckett A, Feberdy JF. The role of the proteinase VCP1 produced by the nematophagous Verticillium chlamydosporium in the infection process of nematode eggs. Mycol Res. 1996;100:421-428.
4. Tikhonov VE, Lopez-Llorca LV, Salinas J, Jansson HB. Purification and characterization of chitinases from the nematophagous fungi Verticillium chlamydosporium and V. suchlasporium. Fungal Genetics and Biology 2002;35:67-78.
5. Manzanilla-Lopez RH. Studies on the characterization and bionomics of Nacobbus aberrans (Thorne, 1935) Thorne & Allen, 1944 (Nematoda: Pratylenchidae)". PhD Thesis, University of Reading, U.K. 1997.
6. Peteira B, Estévez I, Atkins S, Hidalgo-Díaz L, Montes de Oca N, Kerry B. Estabilidad de la cepa IMI SD 187 de Pochonia chlamydosporia var. catenulata (Kamyscho ex Barron y Onions) Zare y W Gams. Parte II. Indicadores bioquímicos. Rev Protección Veg. 2005;20(2):102-109.
7. SAS Institute Statistical Analysis software SAS. Version 8.02. Cary, NC, USA. 2001.
8. Morton CO, Mauchline TH, Kerry BR, Hirsch PR. PCR _ based DNA fingerprinting indicates host _ related genetic variation in the nematophagous fungus Pochonia chlamydosporia. Mycol Res. 2003;107:198-205.
9. Morton CO, Hirsch PR, Kerry BR. Infection of plant _ parasitic nematodes by nematophagous fungi _ a review of the application of molecular biology to understand infection process and to improve biological control. Nematology. 2004;6(2):161-170.
10.Mauchline TH. Studies into the molecular ecology of the nematophagous fungus Verticillium chlamydosporium. PhD Thesis. University of Reading and IACR Rothamsted, UK. 2001.
11.Morton OC, Hirsch PR, Peberdy JP, Kerry BR. Cloning of and genetic variation in protease VCP1 from the nematophagous fungus Pochonia chlamydosporia. Mycol Res. 2003;107(1):38-46.
12.Mauchline TH, Kerry BR, Hirsch PR. The biocontrol fungus Pochonia chlamydosporia shows nematode host preference at the intraspecific level. Mycol Res. 2004;108(2):161-169.
13.Xingzhong ZL, Senyu YC. Nutritional requirements of Pochonia chlamydosporia and ARF18, fungal parasites of nematode eggs. J Invertebrate Pathol. 2003. Disponible en: www.sciencedirect.com.
]]>
]]>
{kind=link}
{kind=link}
{kind=link}