CARACTERIZACIÓN BIOQUÍMICA DE AISLAMIENTOS DE Trichoderma spp. PROMISORIOS COMO AGENTES DE CONTROL BIOLÓGICO. II. EXPRESIÓN DE ACTIVIDAD GLUCANASA
BIOCHEMICAL CHARACTERIZATION OF PROMISING ISOLATES OF Trichoderma spp. AS BIOLOGICAL CONTROL AGENTS. II. EXPRESSION OF GLUCANASE ACTIVITY
Ivonne González*, Danay Infante*, Belkis Peteira*, B. Martínez*, Yailén Arias*, Noyma González*, Ileana Miranda** ]]>
*Grupo de Fitopatología y **Grupo de Plagas Agrícolas. Dirección Protección de Plantas. Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: marquetti@censa.edu.cu
RESUMEN
Los hongos pertenecientes al género Trichoderma secretan enzimas hidrolíticas que le permiten penetrar la pared celular de los hongos fitopatógenos. Las glucanasas son enzimas que hidrolizan los b-glucanos de la pared celular de fitopatógenos como Sclerotium rolfsii, Rhizoctonia solani y Pythium sp. El objetivo de este trabajo fue realizar la dinámica de inducción de glucanasas de diez aislamientos de Trichoderma spp. evaluando la actividad enzimática específica de estas enzimas al primer, tercer, quinto y séptimo días, en los sobrenadantes de los cultivos en tres medios líquidos distintos: medio basal, medio basal suplementado con quitina al 0,5% y medio basal suplementado con gelatina al 0,2%. Los aislamientos 1, 13 y 17 alcanzaron los mayores niveles de actividad enzimática específica glucanasa durante la inducción con el medio suplementado con gelatina. El medio basal también resultó ser un buen inductor de estas enzimas.
Palabras clave: Trichoderma; glucanasas; control biológico; caracterización bioquímica.
ABSTRACT
Trichoderma species fungi secret hydrolytic enzymes which allow them to penetrate the cell walls of phytopathogenic fungi. The glucanases are enzymes that hydrolyze the b-glucans of the cell wall of phytopathogens such as Sclerotium rolfsii, Rhizoctonia solani and Pythium sp. The aim of this work was to carry out the dynamics of glucanase induction of ten Trichoderma spp. isolates evaluating the enzymatic activities at the first, third, fifth and seventh days of the cultures in three liquid media: basal medium, basal medium supplemented with chitin 0,5% and basal medium supplemented with gelatine 0,2%. The isolates 1, 13 y 17 reached the highest levels of glucanase specific activity during the induction with the basal medium supplemented with gelatine. The basal medium also resulted to be a good inductor of those enzymes.
Key words: Trichoderma; glucanases; biological control; biochemical characterization.
]]>
INTRODUCCIÓN
Los hongos del género Trichoderma son activos contra una amplia gama de patógenos foliares y del suelo económicamente importantes, y se utilizan exitosamente como bioplaguicidas en aplicaciones tanto en casas de cultivo como en campo (1, 2). Estos hongos emplean el micoparasitismo como uno de sus mecanismos para ejercer el control biológico, penetrando la pared celular del hongo hospedante y utilizando su contenido celular, mediante enzimas hidrolíticas tales como las quitinasas, glucanasas y proteasas, las cuales se inducen, al menos parcialmente, ante el contacto directo con el hospedante (3).
Las glucanasas son enzimas que degradan b-glucanos y se clasifican en dos grandes grupos según los mecanismos que utilizan para hidrolizar el sustrato, identificados por los productos de hidrólisis: (i) las exo-b-glucanasas, las cuales hidrolizan el sustrato por ruptura secuencial de residuos de glucosa desde el extremo no reductor, y (ii) las endo- b-glucanasas, que rompen aleatoriamente los sitios de enlace b de la cadena polisacarídica, liberando pequeños oligosacáridos. La degradación de los b-glucanos por los hongos se acompaña frecuentemente de la acción sinérgica de las endo- y exo-b-glucanasas (4).
Por otra parte, las paredes celulares de los fitopatógenos Sclerotium rolfsii (Sacc), Rhizoctonia solani (Kühn), y Pythium sp. están en su mayoría compuestos de 1,3-b-glucanos y quitina, y algunos como los Oomycetes (ejemplo, Pythium sp.), que poseen los 1,3-b-glucanos, también contienen celulosa en lugar de quitina (5), lo que explica la importancia de estas enzimas en la capacidad antagónica de Trichoderma y por lo tanto en su utilidad como bioplaguicida. El objetivo de este trabajo fue realizar la dinámica de inducción de las enzimas b-1,3-glucanasas de un grupo de aislamientos de Trichoderma candidatos a biofungicidas (6) utilizando tres medios líquidos con diferente efecto inductor: medio basal, medio basal suplementado con quitina al 0,5% y medio basal suplementado con gelatina al 0,2%.
MATERIALES Y MÉTODOS
Para la inducción de las glucanasas, se emplearon los aislamientos de Trichoderma y los medios líquidos referidos por González et al. (7). Se realizó la cuantificación de proteínas totales siguiendo el método descrito por Bradford (8) utilizando el sobrenadante de los cultivos. Para realizar la determinación de la actividad enzimática de las b-1,3-glucanasas se utilizó una variante en microplacas de 96 pozos desarrollada por Zheng y Wozniak (9).
La actividad específica se determinó según la expresión:
]]>Se realizaron tres repeticiones del experimento y tres réplicas para cada determinación de proteínas totales y de la actividad enzimáticas.
Para determinar los aislamientos que presentaron los mayores niveles de actividad específica glucanasa en cada uno de los medios de cultivo se empleó análisis de varianza simple y la comparación entre las medias se realizó haciendo uso de la prueba de rangos múltiples de Duncan en los casos donde existieron diferencias (p<0.05). Para identificar los medios y días de cultivo que favorecen la inducción de la actividad glucanasa se realizó un análisis de componentes principales. Estos análisis se realizaron con el paquete estadístico InfoStat versión 2009 (10).
RESULTADOS Y DISCUSIÓN
Las enzimas gluconolíticas están ampliamente distribuidas entre las plantas superiores, bacterias y hongos y tienen varias funciones fisiológicas en dependencia de su procedencia. Las b-glucanasas fúngicas son reconocidas por realizar un gran número de funciones entre las que se encuentran la movilización de los glucanos de la pared celular y el almacenamiento de carbohidratos en situaciones de escasez de nutrientes (11), la degradación de calosa de plantas por los fitopatógenos, la nutrición de saprófitos, y la participación en el mecanismo de ataque y nutrición del micoparásito (12). Las b-1,3-glucanasas son una de las enzimas claves responsables de la lisis y la degradación de la pared celular y la pared del esclerotio de los hongos (13,14) y de ahí, la importancia de su estudio.
En un trabajo previo realizado por González et al. (7), se emplearon medios de cultivo con diferente efecto inductor de enzimas hidrolíticas, el cual mostró que el medio líquido basal suplementado con quitina era capaz de inducir elevados niveles de actividad quitinasa en los aislamientos de Trichoderma spp. El-Katatny et al. (15) indujeron la producción de quitinasas y b-1,3-glucanasas extracelulares en Trichoderma harzianum Rifai y Pythium oligandrum Drechsler utilizando medios líquidos suplementados con quitina o micelio seco de hongos probados fito-patogénicamente (Fusarium oxysporum Schlechtend.:Fr y Pythium ultimum Trow) como fuente de carbono. Los micelios secos de F. oxysporum Schlechtend.:Fr fueron la fuente de carbono más selectiva para la producción de enzimas por P. oligandrum Drechsler y T. harzianum Rifai.
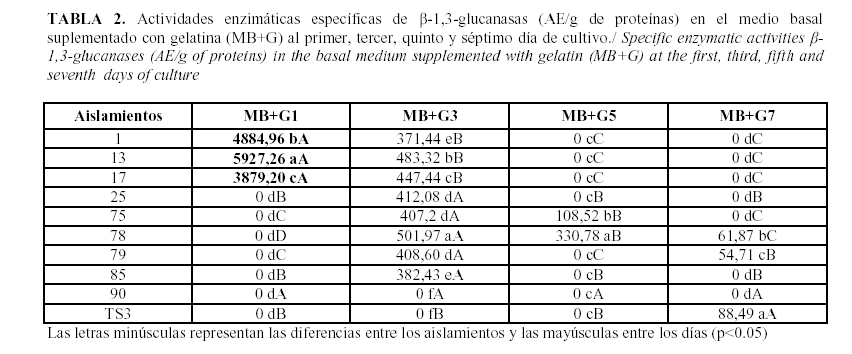
Las actividades enzimáticas específicas para las b-1,3-glucanasas de Trichoderma spp. se comportaron de forma desigual en dependencia de la fuente de carbono empleada (7) (Tablas 1, 2 y 3). Contrario a lo esperado, el medio basal indujo una elevada secreción de esta enzima en los aislamientos 13, 17, 75, 25 y 1 respectivamente (Tabla 1). El medio basal suplementado con gelatina indujo los mayores valores de las actividades enzimáticas de las b-1,3-glucanasas en los sobrenadantes de los cultivos correspondientes a los aislamientos 13, 1 y 17 (Tabla 2). Por su parte, el medio basal que fue suplementado con quitina resultó ser el peor inductor de estas enzimas (Tabla 3).
]]>
El medio a emplear también resulta un factor importante en estos estudios. Huang et al. (2009), realizaron experimentos donde evaluaron la influencia de diferentes fuentes de carbono sobre los patrones de expresión del gen de la endoglucanasa egVIII en Trichoderma viride (Pers, ex Fr.) AS3.3711. Con glucosa (2%) o fructosa (2%) como fuente de carbono, no se detectaron, en ningún momento, transcriptos de egVIII en micelios cultivados. Por otra parte, cuando la sacarosa o el tallo de maíz fueron la fuente de carbono, el RNAm de egVIII alcanzó los mayores niveles a los cuatro días de incubación, mientras que no se detectó señal al primer día. Con celulosa microcristalina, como la fuente de carbono, se observaron resultados similares. Sin embargo, la señal más fuerte se observó con CMC-Na (Carboxi Metil Celulosa-Na) como fuente de carbono: la expresión fue inducida por CMC-Na al primer día, el pico a los cuatro días y decreció después de los cuatro días (16).
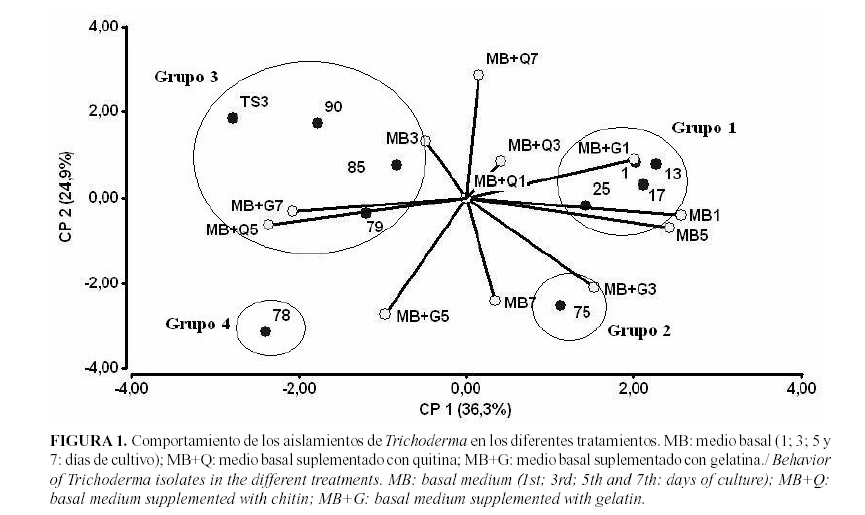
Con dos nuevas variables (componentes principales 1 y 2) fue posible explicar el 61.27% de las asociaciones que se presentaron entre los aislamientos demostrando que el tiempo al cual se alcanza la máxima actividad resultó ser dependiente del medio empleado y del aislamiento estudiado. La primera componente mostró, por su correlación con las variables originales, que algunos aislamientos tenían mayores valores de actividad enzimática glucanasa en los medios basal al primer y quinto día y basal suplementado con gelatina al primer día en contraposición de los que presentaran mejor comportamiento en los medios suplementado con gelatina al séptimo día y suplementado con quitina al quinto día. De manera que los medios suplementados ofrecieron mayor actividad enzimática a medida que pasaron los días. La segunda componente mostró que algunos aislamientos tuvieron mejor comportamiento en el medio basal a los siete días, basal más gelatina al tercer y quinto días contrario a los que obtuvieron altos valores a los siete días en el medio basal suplementado con quitina (Tabla 4).
Estas asociaciones representadas gráficamente mostraron que de forma general, los aislamientos 1, 13 y 17 seguidos del aislamiento 25 presentaron los mayores niveles de expresión de b-1,3-glucanasas, siendo el medio basal suplementado con gelatina junto al medio basal, los más eficaces para realizar la inducción de estas enzimas; ambos durante el primer día de cultivo. El aislamiento 75 también presentó elevados niveles de producción de glucanasas en estos medios pero con la necesidad de esperar hasta el tercer día de cultivo en el medio basal suplementado con gelatina y hasta el séptimo en el medio basal sin suplemento (Fig. 1). Estos resultados tienen estrecha relación con los obtenidos por Martínez et al. (6), los cuales obtuvieron que los aislamientos 17 y 75 eran efectivos para el control de R. solani (Kühn) en campo.
Es importante tener en cuenta que este ensayo solo permite determinar la actividad de las b-1,3-glucanasas, por lo que la producción de b-1,6-glucanasas también pudo influir en la capacidad antagónica de estos aislamientos. Esto podría explicar por qué TS3 mostró bajos niveles de expresión de esta enzima a pesar de ser altamente efectivo en el tratamiento de plantas ornamentales con síntomas de pudrición de la raíz y de la base del tallo, así como de la pudrición de color pardo negruzca, enfermedades asociadas a la presencia de R. solani Kühn, Fusarium spp. y S. rolfsii Sacc, con disminución de la severidad de 100% (17). En la evaluación realizada previamente por González et al. (7) durante la dinámica de inducción de quitinasas, TS3 presentó niveles de actividad bajos de esta enzima, lo que sugiere que la secreción de estas enzimas hidrolíticas no es el mecanismo determinante de la actividad antagónica de TS3 contra los hongos fitopatógenos.
Sin embargo, Liu et al. (18) encontraron que la expresión de genes que codifican para una endoquitinasa, una exoquitinasa y una exo- b-1,3-glucanasa de Trichoderma atroviride Bissett en arroz condujo a un incremento de la resistencia a R. solani Kühn y Magnaporthe grisea (T.T. Hebert) M.E. Barr (18, 19). Estudios con otro agente de control biológico (Beauveria bassiana (Balsamo) Vuillemin), basados en la sobreexpresión de la quitinasa Bbchit1 resultaron en un incremento de la virulencia de este biocontrolador. Los transformantes de B. bassiana (Balsamo) Vuillemin que segregaban una fusión de la proteasa y la quitinasa (CDEP1:Bbchit1), penetraron la cutícula del insecto diana mucho más rápido, con una reducción de la dosis letal media en un 60,5% (20). Los resultados de estos investigadores apuntaron hacia un efecto de sinergismo entre las enzimas hidrolíticas que intervienen en el proceso de la patogénesis, aun cuando una de ellas pudo ser el determinante principal de este atributo fisiológico. De ahí que también se deben tener en cuenta en la evaluación de un aislado las diferentes enzimas hidrolíticas que puedan contribuir según su modo de acción a su eficacia como biocontrolador.
El aislamiento 75 presentó altos niveles de producción de quitinasas en medio basal y en medio basal suplementado con quitina (7) y presenta además altos niveles de glucanasa en idénticas condiciones experimentales, características muy positivas a tener en cuenta según la diana contra la cual pudiera enfrentarse. Esto mostró, por otra parte, que el medio basal garantizó la expresión abundante de ambas enzimas, las cuales son muy importantes para llevar a cabo el micoparasitismo debido a que su acción de conjunto con las proteasas son responsables de la lisis de las hifas de fitopatógenos como R. solani Kühn a través de la digestión de la mayoría de los componentes de la pared celular (21). Los resultados apuntan además hacia la eficacia de los aislamientos 1, 13, 17, 25 y especialmente el 75 (por expresar niveles de actividad quitinasa y glucanasa elevados) como aislamientos promisorios para el control de hongos fitopatógenos.
REFERENCIAS
1. Krause MS, Madden LV, Hoitink HAJ. Effect of potting mix microbial capacity on biological control of Rhizoctonia damping-off radish and Rhizoctonia crown and root rot of poinsettia. Phytopathol. 2001;91:1116-1123.
2. Tondje PR, Roberts DP, Bon MC, Widner T, Samuels GL, Ismaiel A, et al. Isolation and identification of mycoparasitic isolates of Trichoderma asperellum with potential for suppression of black pod disease of cacao in cameroon. Biol Control. 2007;43:202-212.
3. Zeilinger S, Omann M. Trichoderma Biocontrol: Signal Transduction Pathways Involved in Host Sensing and Mycoparasitism. Gene Reg Syst Biol. 2007;1:227-234.
4. Pitson SM, Seviour RJ, Mcdougall BM. Noncellulolytic fungal â-glucanases: Their physiology and regulation. Enzyme Microb Technol. 1993; 15:178-192.
5. De Marco JL, Felix CR. Purification and characterization of a b-glucanase produced by Trichoderma harzianum showing biocontrol potential. Braz Arch Biol Tech. 2007; 50(1):21-29.
6. Martínez B, Reyes Y, Infante D, González E, Baños H, Cruz A. Selección de aislamientos de Trichoderma spp. candidatos a biofungicidas para el control de Rhizoctonia sp. en arroz. Rev Protección Veg. 2008; 23(2):118-125.
7. González I, Infante D, Peteira B, Martínez B, Arias Y, González N, et al. Caracterización bioquímica de aislamientos de Trichoderma spp. promisorios como agentes de control biológico. I: Expresión de actividad quitinasa. Rev Protección Veg. 2010; 25(1):58-63.
8. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem. 1976; 73:248-250.
9. Zheng Y, Wozniak CA. Adaptation of a 1,3 b-glucanase assay to microplate format. Biotechniques. 1997; 22:922-926.
10.InfoStat versión 2009, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. 2009.
11.Kuhn PJ, Trinci APJ, Jung MJ, Goosey MW, Copping LG. Biochemistry of cell walls and membranes in fungi. Berlin/Heidelberg: Springer-Verlag. 1990.
12.Howell CR. Mechanisms employed by Trichoderma species in the biological control of plant diseases: the history and evolution of current concepts. Plant Dis. 2003;87(1):4-10.
13.Cook RJ, Baker KF. The nature and practice of biological control of plant pathogens. Minnesota: American Phytopathological Society. 1983; 539 p.
14.El-Katatny MHA. The activity of b-1,3-glucanase from Trichoderma harzianum in native form and after immobilization on calcium alginate. Arch Phytopathol Pfl. 2008;41(3):175-186
15.El-Katatny MHA, Abdelzaher HMA, Shoulkamy MA. Antagonistic actions of Pythium oligandrum and Trichoderma harzianum against phytopathogenic fungi (Fusarium oxysporum and Pythium ultimum var. ultimum). Arch Phytopathol Pfl. 2006; 39(4):289-301.
16.Huang XM, Yang Q, Liu ZH, Fan JX, Chen XL, Song JZ, et al. Cloning and heterologous expression of a novel endoglucanase gene egVIII from Trichoderma viride in Saccharomyces cerevisiae. Appl Biochem Biotechnol. 2009; 162(1):103-115.
17.Brito R. Tratamiento fitosanitario con Trichoderma sp. para el control de patógenos en la reproducción masiva de plantas ornamentales. Fitosanidad. 2006; 10(2):161-162.
18.Liu M, Sun ZX, Zhu J, Xu T, Harman GE, Lorito M. Enhancing rice resistance to fungal pathogens by transformation with cell wall degrading enzyme genes from Trichoderma atroviride. J Zhejiang Univ Sci. 2004;5:133-136.
19.Djonovic S. Role of two secreted proteins from Trichoderma virens in mycoparasitism and induction of plant resistance. Tesis para optar por el grado científico de PhD. Texas A&M University. 2005.
20.Fang W, Feng J, Fan Y, Zhang Y, Bidochka MJ, et al. Expressing a fusion protein with protease and chitinase activities increases the virulence of the insect pathogen Beauveria bassiana. J Invertebrate Pathol. 2009;102(2): 55-159.
21.Radjacommare R, Venkatesan S, Samiyappan R. Biological control of phytopathogenic fungi of vanilla through lytic action of Trichoderma species and Pseudomonas fluorescens. Arch Phytopathol Pfl. 2010;43(1):1-17.
(Recibido 12-2-2010; Aceptado 29-10-10) ]]>
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}