J. Álvarez*, J. Miguel Cotes**, M. Marín*
* Laboratorio de Biología Celular y Molecular, Facultad de Ciencias, Universidad Nacional de Colombia, Calle 59A No 63-20, Medellín, Colombia. Email: mamarinm@unal.edu.co; **Departamento de Ciencias Agronómicas, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Calle 59A No 63-20, Medellín, Colombia
RESUMEN
La virosis del tomate de árbol en Colombia es causada por un complejo de al menos siete virus, que incluyen miembros de los géneros Potyvirus, Cucumovirus, Polerovirus y Tobamovirus, entre otros. La propagación de este frutal se realiza por semilla sexual, por lo cual es fundamental garantizar su sanidad, especialmente de virus como PVY y PLRV, que han sido informados y pueden ser transmitidos por esta vía. En la investigación se realizaron los análisis filogenéticos de aislados de PVY y PLRV obtenidos a partir de muestras de semilla sexual y plántulas de vivero de tomate de árbol procedentes de diferentes regiones de Colombia. También se desarrollaron técnicas de hibridización molecular con sondas dirigidas al gen de la cápside, como herramientas de diagnóstico para el género Potyvirus y la especie PLRV. El análisis filogenético de secuencias para PVY, agrupó los aislados de plantas de vivero con aquellas informadas en tejido foliar de plantas adultas, no asociándose con ninguno de los grupos que representan las razas conocidas de PVY. En el caso de PLRV, los aislados secuenciados se asociaron indistintamente con los obtenidos de cultivos de papa en diferentes países. La técnica de dot-blot presentó niveles de detección muy similares a pruebas de ELISA, pero no así la metodología de tissue-printing, que detectó un 15% menos de las muestras que resultaron positivas mediante ELISA. Este trabajo evidencia la necesidad del desarrollo de programas de certificación de semilla para este frutal en latinoamérica. ]]>
Palabras clave: ELISA; hibridización molecular; PLRV; PVY; Solanum betaceum.ABSTRACT
Tamarillo virus disease is caused in Colombia by a complex of at least seven viruses, including some members of the genera Potyvirus, Cucumovirus, Polerovirus and Tobamovirus. The propagation of this fruit crop is made by sexual seed; therefore, it is important to warranty its health, especially because PVY and PLRV can be transmitted through this way. In this research, the phylogenetic analysis of PVY and PLRV isolates was carried out. They were detected in sexual seeds and plant seedlings of tamarillo obtained from different Colombian provinces. Additionally, the molecular hybridization techniques with probes for the capsid gene were developed as diagnostic tools for the genus Potyvirus and the species PLRV. The phylogenetic analysis of PVY sequences grouped the isolates obtained from seedlings with those previously reported from leaf tissue of adult plants, but not with any of the clades representing the known PVY strains. In the case of PLRV, the strains sequenced were associated to isolates of potato crops from different countries. Tests of dot-blot showed similar levels of detection to conventional ELISA, but not to the tissue-printing method, which detected a 15% less than positive samples detected by ELISA. This study shows the urgent need to develop seed certification programs for this crop in Latin America.
Key words: ELISA; molecular hybridization; PLRV; PVY; Solanum betaceum.
(Recibido 26-3-2011; Aceptado 20-5-2011)
INTRODUCCIÓN
El cultivo del tomate de árbol (Solanum betaceum Bosh) se extiende en Colombia aproximadamente a 7 500 ha distribuidas en 18 departamentos del país (1). Durante el período 1992-2003 el área sembrada de este cultivo presentó un aumento de 6,1% anual; sin embargo en los últimos años esta tendencia se ha revertido en departamentos como Antioquia y Cundinamarca, donde la extensión del cultivo se ha reducido principalmente por diversos problemas fitosanitarios (2,3). La virosis es una de las causas que limitan, ya que no sólo reduce la cantidad y calidad de fruta producida, sino también el período productivo de los árboles. Recientemente, se ha detectado la presencia de un complejo viral asociado a este cultivo en Colombia, que incluye al menos dos potyvirus (Potato virus Y - PVY y Tamarillo leaf malformation virus - TaMLV), además de las especies Alfalfa mosaic virus (AMV), Cucumber mosaic virus (CMV), Potato leaf roll virus (PLRV), Tomato mosaic virus (ToMV) y Tomato ringspot virus (ToRSV) (2,3,4,5). La virosis se caracteriza por una sintomatología compleja que incluye la presencia de mosaicos con ampollamientos, deformaciones foliares y cambios en la apariencia de flores y frutos, bandeado de venas, grabados no geométricos en hojas y frutos, amarillamiento de venas, manchas aceitosas, anillos necróticos y bronceado del tejido foliar. Sin embargo, la etiología de cada uno de estos síntomas no se ha identificado plenamente (2,3,6).
Los estudios de los mecanismos de transmisión de virus en tomate de árbol se han abordado muy tangencialmente, al basarse fundamentalmente en la manifestación de síntomas y no en el uso de técnicas altamente sensibles como las serológicas y/o moleculares. En algunos trabajos en Colombia y otros países se ha encontrado que algunos de los potyvirus asociados al cultivo son transmitidos en forma mecánica, por propagación asexual y por insectos vectores de la familia Aphididae (7,8,9,10), siendo en Colombia informadas las especies Macrosiphum euphorbiae y Aphis gossypii como transmisoras de CMV y PVY en diferentes regiones del país (11). Recientemente, a partir de estudios serológicos en plántulas germinadas de tomate de árbol bajo condiciones de invernadero, se encontró que los virus PVY y PLRV eran transmitidos por semilla sexual (12), lo cual representa el primer informe mundial de la transmisión de un polerovirus por este medio.
El cultivo del tomate de árbol es propagado por los agricultores mediante semilla sexual, por lo que es necesario continuar con los estudios de transmisión de virus a través de este mecanismo, así como el desarrollo de metodologías que ofrezcan herramientas de detección viral más precisas a los productores de semilla, distribuidores de material de siembra y a los organismos de sanidad vegetal de los estados, con miras a la generación en Latinoamérica de semilla certificada de tomate de árbol libre de patógenos virales. En este estudio se evaluaron las relaciones filogenéticas de los aislados de PLRV y PVY obtenidas de semilla sexual y plántulas de vivero, a partir de análisis de secuencias del gen de la cápside. Adicionalmente, se establecieron metodologías de hibridización tipo dot-blot y tissue-printing, que pudieran ser empleadas en forma rutinaria para el diagnóstico de PLRV y potyvirus, realizándose comparaciones con las técnicas de ELISA y RT-PCR. En este caso se estimó más conveniente establecer dichas metodologías de hibridización para potyvirus, del cual hace parte el PVY, de manera que se ampliará el rango de detección a otros miembros de dicho género encontrados en el cultivo como TaLMV y Tamarillo mosaic virus (TaMV).
MATERIALES Y MÉTODOS
Aislados virales ]]>
Las muestras de semillas y plántulas de vivero utilizadas en este estudio, procedieron de material almacenado y colectado en los departamentos de Antioquia, Cundinamarca, Nariño y Putumayo como resultado de un estudio de evaluación serológica de incidencia de virus en material de siembra de tomate de árbol en Colombia (12). Para esta investigación, se evaluó mediante RT-PCR la presencia de los virus PLRV y PVY en al menos una muestra de semilla y de plántulas de vivero por cada región bajo estudio de los cuatro departamentos (Antioquia: Carmen de Viboral, La Unión-Sonsón, Entrerríos y Santa Rosa-Yarumal; Cundinamarca: Anolaima, Granada y San Bernardo; Nariño: Córdoba; Putumayo: Colón). Para evitar contaminación externa con pulpa de fruta, todas las muestras de semillas fueron tratadas con jabón antibacterial, Tween-20 e hipoclorito de sodio al 12% por 15 min, siguiendo el protocolo de Álvarez (12).Extracción de ácidos nucleicos
Debido a que los tejidos de tomate de árbol presentan gran cantidad de inhibidores de las reacciones enzimáticas, fue necesario utilizar tres metodologías de extracción de ácidos nucleicos: ARN total mediante kit comercial; extracción de ácidos nucleicos totales e inmunocaptura post-ELISA. De esta forma, cuando no era posible la detección de un virus con el kit comercial, se repetía el proceso con el segundo método y finalmente se procedía con la inmunocaptura.
Extracción de ARN total. Se utilizó el kit RNeasy plant mini de Qiagen (California, EEUU), partiéndose de la maceración con nitrógeno líquido de 100 mg de semilla o de tejido foliar, utilizando 450 ìL de buffer RLT (incluido en el kit) y 4,5 ìL de â-mercaptoetanol.
Extracción de ácidos nucleicos totales. Se partió de 100 mg de semilla o tejido vegetal macerándose con nitrógeno líquido y adicionándose 450 ìL del buffer de extracción (50 mM Citrato de sodio pH 8,5; 0,1% Triton X-100; 5 mM DTT y 10 mM 2-â-mercaptoetanol) siguiendo el protocolo de Ivars et al. (13).
Inmunocaptura post-ELISA. Para esta metodología se realizaron pruebas de DAS-ELISA con anticuerpos para PVY y PLRV de la compañía Agdia (Indiana, EEUU), siguiendo las instrucciones del fabricante. En aquellas muestras que resultaron positivas, se procedió a liberar las partículas virales con 70 ìL de buffer de liberación (Tris-HCl 10 mM pH 8,0; 1% Triton X-100) e incubando a 70°C durante 10 min (14).
RT-PCR y secuenciación
Las reacciones de RT-PCR se realizaron con el kit Qiagen OneStep RT-PCR, con 5 µL de ARN, 0,6 µM de cada cebador específico para un fragmento del gen que codifica para la proteína de la cápside viral de 480 pb para PVY (PVY-F: 5'-ACGTCCAAAATGAGAAT GCC-3', PVY-R 5'-TGGTGTTCGTGATGTGACCT-3', ubicados entre las posiciones 8717 y 9196 del genoma viral) y de 336 pb para PLRV (PLRV-F: 5'-CGCGCTAACAGAGTTCAGCC-3', PLRV-R: 5'-GCAATGGGGGTCCAACTCCAACTCAT-3', ubicados entre las posiciones 3670 y 4005 del genoma viral) (15,16), 1X de Buffer de enzimas, 400 µM de dNTPs y 2 µL de mezcla de enzimas (transcriptasa reversa Omniscript, transcriptasa reversa Sensiscript y Polimerasa de ADN HotStarTaq). El programa de amplificación consistió de una fase inicial de síntesis de la primera cadena a 50°C por 30 min, seguido de la activación de la Taq polimerasa a 95°C por 15 min y 35 ciclos a 94°C por 1 min, 52°C por 1 min, 72°C por 1,30 min y una extensión final de 10 min a 72°C.
Los amplicones del tamaño esperado, fueron purificados directamente del gel mediante el kit QIAquick Gel Extraction (Qiagen), para proceder a su secuenciación en ambos sentidos en un secuenciador ABI Prism 3730xl (PE Applied Biosystems).
Las secuencias obtenidas en ambos sentidos fueron editadas mediante los programas BioEdit 6.0.6 y Chromas 1.45, confirmándose su identidad con genes virales por comparación con el GenBank, mediante BLASTn. Esta información sirvió de base para realizar análisis filogenéticos con los aislados reportados por Jaramillo (3) en tejido foliar de plantas adultas con virosis y con secuencias de virus obtenidos en otros hospedantes y regiones del mundo. Para esto se alinearon las secuencias mediante el programa Clustal W y la matriz generada fue utilizada para obtener un árbol filogenético basado en Máxima Parsimonia con el programa PAUP 4.0b (17). El soporte de la topología interna de los dendrogramas se determinó por análisis de bootstrap con 1 000 remuestreos (18). Los números de accesión de las secuencias utilizadas en el análisis se presentan en las Figuras 1 y 2. ]]>
Detección de virus mediante hibridización molecularSe evaluaron las metodologías dot-blot y tissue-printing para detectar los virus PLRV y potyvirus en semilla (dot-blot) y plántulas de vivero (dot-blot y tissue-printing). El procedimiento se estableció a partir de la detección en 20 muestras de plántulas tomadas aleatoriamente de un vivero comercial y 20 muestras de semillas obtenidas en cultivos del Oriente de Antioquia. El experimento basado en dot-blot estuvo acompañado de evaluaciones previas mediante pruebas ELISA y RT-PCR para todas las muestras. Con el fin de comparar los niveles de detección obtenidos entre dichas pruebas, se calculó el Coeficiente de concordancia Kappa, mediante:
![]() ,
,
donde:
 y
y ![]() y
y ![]() indican los porcentajes observados correspondientes a la tabla de dos vías entre los dos métodos evaluados. Similarmente, se obtuvieron los limites de confianza mediante la siguiente expresión:
indican los porcentajes observados correspondientes a la tabla de dos vías entre los dos métodos evaluados. Similarmente, se obtuvieron los limites de confianza mediante la siguiente expresión:
 ,
,
donde: ![]() es el cuantil de la distribución normal estándar correspondiente a un nivel de confianza y el error estándar del valor estimado del kappase obtuvo mediante:
es el cuantil de la distribución normal estándar correspondiente a un nivel de confianza y el error estándar del valor estimado del kappase obtuvo mediante:

]]>
Las pruebas de ELISA utilizaron anticuerpos específicos de la compañía Agdia para PLRV y para los miembros del género Potyvirus, y fueron realizados bajo los formatos de DAS y ACP-ELISA. Los resultados colorimétricos fueron cuantificados en un equipo Multiscan (Labsystem, Finlandia). Cada prueba incluyó un control positivo (suministrado por el fabricante en forma liofilizada) y un control negativo. Los pozos considerados con reacción positiva fueron aquellos en los cuales la lectura de absorbancia a 405 nm presentaba un valor mínimo del doble de la lectura obtenida en el control negativo, siguiendo el criterio de Matthews (5). Por otra parte, las pruebas de RT-PCR se realizaron tal como se describió anteriormente, pero en lugar de utilizar cebadores específicos para el gen de la cápside de PVY, se emplearon los cebadores PNBF1 (5'-GG(GCT)AA(CT)AATAGTGG(AGCT)CAACC-3') y PCPR1 (5'-GGGGAGGTGCCGTTCTC(AGT)AT(AG) CACCA-3', ubicados entre las posiciones 8125 y 9197 del genoma viral ) que amplifican un fragmento de 1050 a 1200 pb de las regiones Nib y cápside viral, en los miembros del género Potyvirus (20).
Similarmente, esta evaluación se realizó para la prueba de tissue-printing, comparándose sus niveles de detección con respecto a las metodologías dot-blot y ELISA.
Hibridización dot-blot. Esta metodología se basó en los procedimientos desarrollados por Ivars et al. (13) y Mas y Pallas (19). Se partió de 100 mg de tejido (semilla sexual o tejido foliar) macerándose en 500 ìL de buffer de extracción (50 mM Citrato de sodio pH 8,5; 0.1% Tritón X-100; 5 mM DTT y 10 mM 2-â-mercaptoetanol) para posteriormente centrifugar por 5 min a 6 000 rpm. El sobrenadante generado fue aplicado directamente a la membrana o alternativamente se evaluó la purificación de ácidos nucleicos totales de la misma manera descrita anteriormente. Posteriormente, se tomaron 10 ìL para cada tipo de extracto y se adicionó un volumen de SSC 10X (1,5 M NaCl; 0,15 M Citrato de sodio pH 7,0) incubándose por 15 min a 60°C. A partir de ambos tipos de extracto se tomaron 4 ìL y se depositaron sobre una membrana de nylon Hybond N+ (Amersham, Ingleterra), fijando mediante exposición a luz UV por 2 min a 50 000 ìJ.cm2. Las membranas fueron prehibridizadas por 2 h a 68°C para potyvirus, y a 64°C para PLRV, en el buffer de hibridización (SSC 5X, Solución Denhards 5X; 0,5% SDS) suplementado con 50% de formamida y 100 ìg.mL-1 de ADN de esperma de salmón.
La sonda fue preparada utilizando el kit de marcaje: Biotin DecaLabel DNA Labeling Kit (Fermentas, Lituania), en el cual se adicionaron 10 ìL del producto de PCR específico para PLRV y para potyvirus, siguiendo las instrucciones del fabricante. La sonda se desnaturalizó a 95°C y fue adicionada al buffer de hibridización durante la noche, para proceder al lavado de la membrana a temperatura ambiente así: dos lavados por 5 min con SSC 2X suplementado con 0,1% de SDS, dos lavados por 10 min con SSC 1X con 0,1% de SDS y cuatro lavados de 5 min cada uno con SSC 0,1X y 0,1% de SDS.
Finalmente, la detección se realizó mediante el uso del kit Biotin Chromogenic Detection (Fermentas), que utiliza streptovidina conjugada a fosfatasa alcalina, siguiendo las instrucciones del fabricante.
Tissue-printing. Esta metodología se basó en los protocolos desarrollados por Mas y Pallas (19), a partir de la impresión de pecíolos cortados de plántulas de vivero directamente sobre membranas de Nylon Hybonnd N+ (Amersham), exponiéndose a luz UV por 2 min a 50 000 ìJ.cm2 en un transiluminador, para proceder a la prehibridización por 2 h a 58°C para potyvirus y a 54°C para PLRV, siguiendo los procedimientos indicados para dot-blot.
RESULTADOS
]]> RT-PCR y secuenciación
Las pruebas de RT-PCR permitieron obtener amplicones del tamaño esperado para los virus PLRV y PVY, los que fueron confirmados como asociados a los genes de la cápside mediante secuenciación. La amplificación de la región de 336 pb de la cápside de PLRV fue obtenida en 22 de las muestras, 19 de semillas procedentes de los cuatro departamentos bajo estudio, una muestra de plántula del vivero de Carmen de Víboral y de dos plántulas germinadas en el invernadero a partir de semillas obtenidas en Anolaima. Para el caso de PVY, se obtuvieron tres amplicones de 480 pb que procedían de muestras de plántulas de vivero del municipio de Carmen de Víboral, no pudiéndose obtener de otras regiones ni de muestras de semillas, a pesar de los múltiples ensayos realizados a partir de diferentes metodologías de extracción de ácidos nucleicos y condiciones de RT-PCR.
Análisis filogenético para PLRV
El análisis filogenético para PLRV utilizó 321 posiciones, 238 de las cuales fueron constantes, 69 variables pero no informativas y 14 informativas para el análisis de máxima parsimonia. El Dendrograma generado presentó un Índice de Consistencia (CI) de 0,92, Índice de Retención (RI) de 0,84 y un Índice de Homoplasia (HI) de 0,08 y relacionó todos los aislados de PLRV en un solo grupo soportado por un valor de bootstrap del 100% y diferenciado de la secuencia de CYDV-RPV utilizada como grupo externo (Outgroup) (Fig. 1). El grupo de PLRV, incluyó indistintamente aislados obtenidos de cultivos de papa de Colombia y de diferentes países como India, China, Perú e Irán con los ocho aislados secuenciados a partir de semillas obtenidas de cultivos de los cuatro departamentos bajo análisis, así como de un aislado de plántulas de viveros y uno obtenido de la germinación de semillas mantenidas bajo invernadero.
La matriz de comparación generada a partir de dichas secuencias y las de referencia, indicó un alto nivel de identidad para la región parcial del gen de la cápside de PLRV, siendo en todos los casos superior al 96%. Los niveles de identidad entre los aislados de PLRV de tejido foliar de plantas adultas de tomate de árbol obtenidas por Jaramillo (3) y las generadas en esta investigación fueron superiores al 98% en todos los casos, así como lo fueron éstas con el aislado 4 Col obtenida de un cultivo de papa del municipio de la Unión (Antioquia) por Gil et al. (21). La identidad de los aislados de PLRV con aquella de CYDV-RPV fue cercana al 77%.
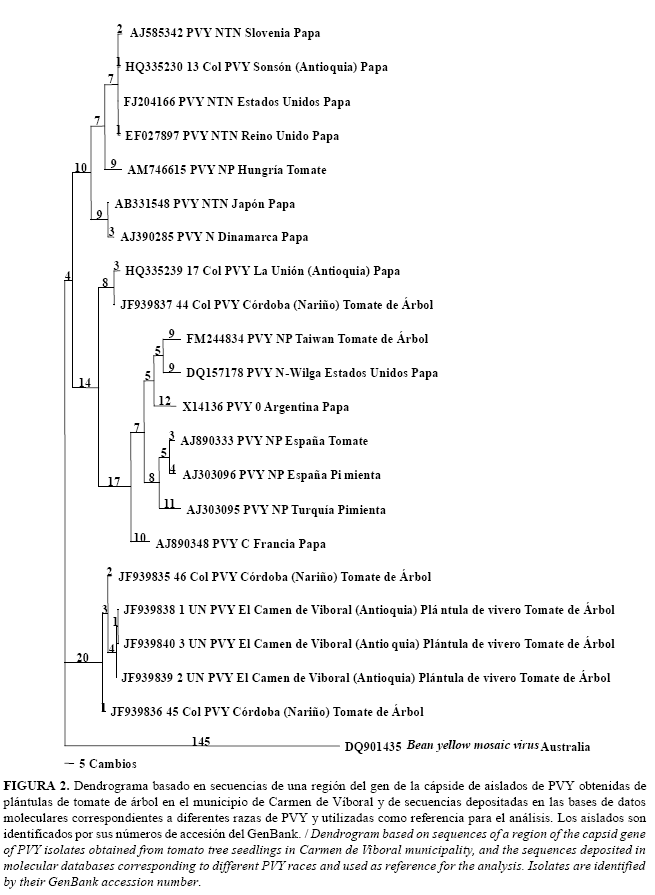
Análisis filogenético para PVY
El análisis filogenético para PVY utilizó 480 posiciones, 272 de las cuales fueron constantes, 111 variables pero no informativas y 98 informativas para el análisis de máxima parsimonia. El Dendrograma generado presentó un CI de 0,74, RI de 0,80 y HI de 0,25 y separó los tres aislados secuenciados en este estudio y procedentes de plántulas de vivero del municipio de Carmen de Víboral con dos aislados (45 y 46 Col), recientemente caracterizadas por Jaramillo (3) de tejido foliar de plantas del municipio de Córdoba (Nariño). Este grupo estuvo fuertemente soportado por un valor de Bootstrap de 100% y a pesar de presentarse relacionado con la especie PVY, se ubicó separado de los aislados de referencia de las diferentes razas de PVY (PVYO, PVYC, PVYN, PVYNTN y PVYN-Wilga, PVY-NP) (Fig. 2).
La matriz de comparación indicó un nivel de identidad superior al 98% para la región parcial del gen de la cápside de PVY de los tres aislados de plántulas de tomate de árbol y aquellas de tejido foliar de plantas con virosis en cultivos de Córdoba (Nariño); sin embargo, dichos niveles de identidad tan sólo alcanzaron valores del 86-87% con respecto a las secuencias representativas de las razas PVYO, PVYC y PVY-NP de tomate de mesa y pimentón de Europa y de 90-91% con aislados de las razas PVYN y PVYNTN. Así mismo, la identidad de los aislados de plántulas de tomate de árbol con respecto a un aislado secuenciado a partir de papa por Gil et al. (21) y de un aislado de tejido foliar de tomate de árbol obtenido por Jaramillo (3) fue del 91%. La identidad de los aislados de PVY obtenidos en este estudio con respecto a la secuencia de BYMV utilizada como grupo externo fue de 65%.
Detección de virus mediante hibridización molecular
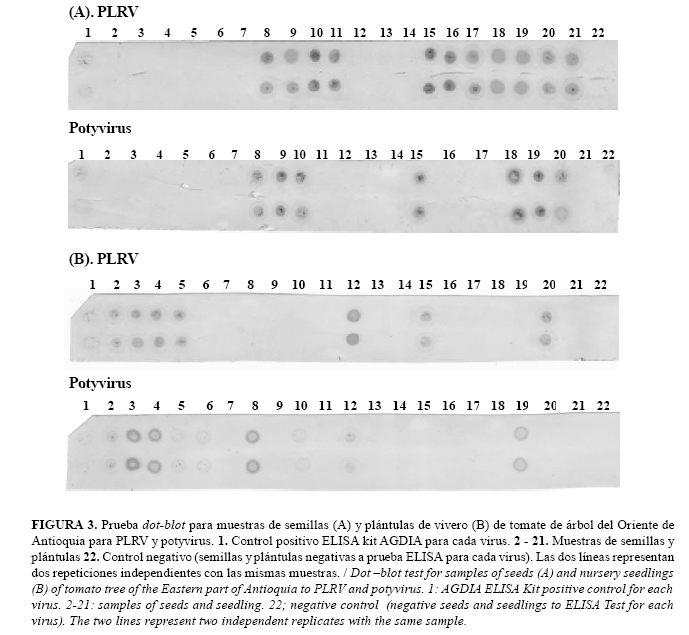
Los procedimientos aplicados para la realización de las metodologías de hibridización molecular tipo dot-blot y tissue-printing condujeron al establecimiento de ambas técnicas para la detección de PLRV y potyvirus, a partir de material de siembra de tomate de árbol, aunque la lectura del revelado de las membranas fue más objetiva para el caso de dot-blot, pues la prueba de tissue-printing puede generar distorsiones en las regiones impresas y mayores niveles de fondo no deseado (background) que dificultan las lecturas finales. En la Figura 3 se presentan los resultados de las pruebas dot-blot para las 20 muestras de semillas evaluadas y de plántulas de vivero, en las que se detectó la presencia de potyvirus en siete y nueve de las muestras respectivamente, mientras que potyvirus se detectaron en 11 muestras de semillas y siete de plántulas. Adicionalmente, es importante indicar que las dos repeticiones realizadas en la membrana para cada muestra coincidieron 100%. Por su parte, la prueba de tissue-printing detectó la presencia de potyvirus en seis plántulas y de PLRV en cuatro muestras (Fig. 4), pero la coincidencia entre ambas repeticiones no fue completa, por las dificultades que se mencionaron para determinar el resultado de la prueba en algunas de las muestras. ]]>
Al comparar los niveles de detección en semilla sexual de tomate de árbol de los virus PLRV y potyvirus logrados con las pruebas de hibridización contra las pruebas ELISA y RT-PCR, se encontró un alto nivel de concordancia entre la metodología de dot-blot y la prueba de ELISA con un coeficiente de Kappa de 1 y 0,89 para la detección de PLRV y potyvirus, respectivamente (Tabla 1), detectándose mediante dot-blot potyvirus enuna proporción adicional del 5%. Por otro lado, la concordancia de estas pruebas con la RT-PCR resultó muy baja o inexistente (Tabla 1), posiblemente debido al efecto de los inhibidores enzimáticos.
Las comparaciones realizadas a partir de las pruebas de tissue-printing indicaron niveles medios de concordancia entre esta metodología y ELISA ó dot-blot, con valores de coeficientes de Kappa que oscilaron entre 0,63 y 0,88 para la detección de PLRV y potyvirus con respecto a pruebas de ELISA y de 0,63 y 0,69 cuando se comparó tissue-printing con dot-blot (Tabla 2). En ambas situaciones la prueba de tissue-printing
detectó el PLRV en un 15% menos de plántulas con relación a las otras dos técnicas bajo estudio. Similarmente, la detección de potyvirus fue realizada en 5 y 15% más de plántulas cuando se utilizaron pruebas de ELISA y dot-blot, respectivamente, que cuando se usó tissue-printing.
DISCUSIÓN
En los últimos años, la enfermedad denominada virosis del tomate de árbol ha venido tomando gran importancia en Colombia, posicionándose como el problema fitosanitario más limitante en la producción de este frutal (2,3,6). Álvarez (12) mediante pruebas serológicas y moleculares determinó la transmisión en esta planta de los virus PLRV y PVY vía semilla sexual, lo cual representa un aspecto epidemiológico fundamental a ser tenido en cuenta, por cuanto los agricultores y viveristas de países latinoamericanos utilizan este medio como base para el establecimiento de sus cultivos y para la comercialización de plántulas de tomate de árbol.
]]>
Por otra parte, el análisis de las relaciones filogenéticas de PVY reforzó lo encontrado recientemente por Jaramillo (3) a partir de aislados obtenidas de tejido foliar con virosis en plantas adultas, en el sentido que algunos de los aislamientos de PVY asociados al tomate de árbol conforman un grupo independiente de las razas tradicionales del virus caracterizadas en papa (PVYN, NTN, O, Z, W, C) y en otros cultivos como pimentón y tomate de mesa (PVY-NP) (24,25); lo cual sugiere que posiblemente los aislados de PVY en tomate de árbol constituyen una variante patogénica no caracterizada hasta ahora en esta especie viral. Al igual que para el caso de PLRV, de gran interés resultaría la realización de pruebas de patogenicidad cruzada de aislados de PVY de diferentes razas de papa y de otras especies solanáceas sobre tomate de árbol y viceversa. Igualmente, sería necesario secuenciar una mayor porción del genoma de este virus, especialmente completar la secuencia de todo el gen de la cápside viral y obtener las secuencias de HC-Pro y NIa, pues en estos genes se han detectado puntos de recombinación asociados a los genotipos PVYNTN y PVYN-Wilga, que causan necrosis de tubérculos de papa y son serológicamente relacionadas con las razas PVYN y PVYO (24,25).
Desafortunadamente, en este trabajo la obtención de un mayor número de secuencias de aislados de PVY no fue posible, a pesar de la utilización de diferentes metodologías de extracción de ácidos nucleicos y de la variación de las condiciones de la RT-PCR. La ausencia de amplificación o la generación de fragmentos inespecíficos es una situación frecuente en los estudios que utilizan la técnica de PCR o RT-PCR, como resultado de la inhibición enzimática de algunos extractos vegetales o a la falta de especificidad de los cebadores, que depende en buena medida del rango de variación de los genotipos considerados al momento de su diseño, lo que en sentido práctico siempre representará una restricción cuando se presentan variantes virales no antes caracterizadas (26). Futuros trabajos deben contemplar este aspecto e incluso proponer metodologías alternativas que conduzcan al diseño de cebadores específicos adaptados a los genotipos virales que se presentan en los cultivos de tomate de árbol de Colombia y otros países latinoamericanos, como es el caso aquí presentado para PVY, cuyas secuencias presentaron divergencias superiores al 9% con respecto a los aislados representativos de las razas clásicas de dicho patógeno.
Por otra parte, la prueba de dot-blot resultó muy eficiente para la detección de potyvirus y de PLRV, siendo similar en sus niveles de detección a las pruebas de ELISA para PLRV, e incluso, alcanzando niveles superiores a la técnica de ELISA para el caso de potyvirus, con un 5% y 10% de mayor detección en material de semilla y de plántulas de vivero, respectivamente. Sin embargo, durante la estandarización de la metodología de dot-blot fue evidente la necesidad de realizar una extracción total de ácidos nucleicos, pues en aquellos ensayos donde se aplicaba a la membrana el macerado directo de tejido (datos no mostrados), era muy bajo o inexistente el nivel de detección alcanzado. Esta metodología es una excelente alternativa a la RT-PCR, que se ve fuertemente afectada por inhibidores enzimáticos presentes en los tejidos de tomate de árbol (2,3,12), lo que conduce a requerimientos de tratamientos adicionales de purificación de ácidos nucleicos como por ejemplo kits comerciales o lavados adicionales con fenol:cloroformo.
La prueba de tissue-printing no arrojó un resultado satisfactorio debido a la alta presencia de fuertes señales basales y a la subjetividad consiguiente que se requiere en la evaluación. Sin embargo, ya que este tipo de metodologías donde se establece impresión de tejidos sobre membrana puede ser muy informativa con respecto a la ubicación del virus en los diferentes tejidos, se recomienda continuar con su proceso de mejoramiento. En nuestro caso, los resultados de las pruebas con plantas de vivero, indicaron que la impresión de pequeños pecíolos de plántulas de apenas dos meses podría dar evidencia de la presencia de ambos grupos de virus (PLRV y potyvirus) en dichas plantas, aunque no fue posible identificarse claramente su ubicación sobre un tejido en particular.
Se espera que los resultados presentados en esta investigación, sirvan de base para el diseño de herramientas de diagnóstico asintomático y de material de siembra de virus en cultivos de tomate de árbol de países latinoamericanos cultivadores de este frutal, de manera que se emprendan acciones para la generación de materiales certificados libres de virus.
AGRADECIMIENTOS
Esta investigación fue financiada por Colciencias (proyecto 1118-405-20317) y DIME-UNALMED (proyecto 20101007745) convocatoria Bicentenario 2009. Se agradece a la Prof. Luz Estela Lagos de la Universidad de Nariño, por el apoyo en la colección de muestras en Nariño y Putumayo.
]]>
REFERENCIAS1. Ministerio de Agricultura y Desarrollo Rural (MADR). Cadena productiva de la papa en Colombia. En: http://www.minagricultura.gov.co; consulta: Junio de 2009.
2. Ayala M, González P, Gutiérrez P, Cotes JM, Marín M. Caracterización serológica y molecular de potyvirus asociados a la virosis del tomate de árbol en Antioquia (Colombia). Acta biol Colomb. 2010; 15:143-162.
3. Jaramillo M. Análisis serológico y molecular de virus asociados al cultivo de tomate de árbol (Solanum betaceum) en Colombia. Medellín, Facultad de Ciencias, Universidad Nacional de Colombia. Tesis de Maestría en Biotecnología; 2009. 151 pp.
4. Gil JF, Ayala M, González P, Marín M. Identificación de potyvirus en cultivos de tomate de árbol (Solanum betaceum Cav) en Antioquia mediante detección serológica. Revista Politécnica. 2009; 8:112-120.
5. Matthews REF. Diagnosis of plant virus diseases. Boca Ratón, FL: CRC Press. 1993; 374p.
6. Rodríguez V, Jaramillo M, Lagos LE, Gutiérrez P, Marín M. Identificación serológica y molecular de virus del tomate de árbol (Solanum betaceum) en cultivos de Córdoba (Nariño, Colombia). Rev LaSallista. 2011; 8:15-30.
7. Vizuete B, Insuasti ML, Ochoa J, Ellis M. Biological and serological characterization of tree tomato virus diseases in Ecuador. INIAP, Ohio State University. 1990; 3 p.
8. Eagles R, Gardner R, Forster R. Incidence and distribution of six virus infecting Tamarillo (Cyphomandra betaceum) in New Zealand. N.Z. J Crop Hortic Sci. 1994; 22:453-458.
9. Bernal, JA, Saldarriaga A, Zapata JL. Evaluación de la transmisión del virus del tomate de árbol. Informe final proyecto CORPOICA-PRONATTA. 1998. 55p.
10. Betancourth C, Goyes R, Bravo DA. Caracterización biológica de un virus del tomate de árbol (Solanum betaceum Send) en el departamento de Nariño. Fitopatol Colomb. 2003; 2:7-10.
11. Martínez J, Cotes JM, Marín M. Detección serológica y molecular de virus en áfidos asociados a cultivos de tomate de árbol con síntomas de virosis en Antioquia y Nariño (Colombia). Rev Fac Ciencias Básicas. 2010; 6:182-197
12. Álvarez J. Caracterización serológica y molecular de virus asociados al material de siembra de tomate de árbol (Solanum betaceum Bosh) en Colombia. Bogotá, Facultad de Ciencias, Universidad Nacional de Colombia. Tesis de Maestría en Microbiología; 2010.
13. Ivars P, Alonso M, Borja M, Hernández C. Development of a non-radioactive dot-blot hybridization assay for the detection of Pelargonium flower break virus and Pelargonium line pattern virus. Eur J Plant Pathol. 2004; 110:275-283.
14. Kogovsek P, Gow L, Pompe-Novak M, Gruden K, Foster GD, Boonham N, et al. Single-step RT real-time PCR for sensitive detection and discrimination of Potato virus Y isolates. J Virol Methods. 2008; 149:1-11.
15. Singh RP, Kurz J, Boiteau G, Bernard G. Detection of Potato leafroll virus in single aphids by the reverse transcription polymerase chain reaction and its potential epidemiological application. J Virol Methods. 1995; 55:133-143. ]]>
16. Nie X, Singh RP. A novel usage of random primers for multiplex RT-PCR detection of virus and viroid in aphids, leaves, and tubers. J Virol Methods. 2001; 91:37-49.
17. Swofford DL. PAUP: Phylogenetic analysis using parsimony (* and other methods). Versión: 4. Sinauer Associates. Sunderland, EEUU; 1998.
18. Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985; 39:783-791.
19. Más P, Pallas V. Non-isotopic tissue-printing hybridization: a new technique to study long-distance plant virus movement. J Virol Methods. 1995; 52:317-326.
20. Hsu YC, Yeh TJ, Chang YC. A new combination of RT-PCR and reverse dot blot hybridization for rapid detection and identification of potyviruses. J Virol Methods. 2005; 128:54-60. ]]>
21. Gil JF. Diagnóstico y caracterización molecular de virus asociados al cultivo de la papa en Colombia, con énfasis en el virus mop-top (PMTV, Pomovirus). Medellín, Facultad de Ciencias, Universidad Nacional de Colombia. Tesis de Maestría en Biotecnología; 2009.
22. Guyader S, Ducray DG. Sequence analysis of Potato leafroll virus isolates reveals genetic stability, major evolutionary events and differential selection pressure between overlapping reading frame products. J Gen Virol. 2002; 83:1799-1807.
23. Plchova H, Cerovska N. Molecular analysis of Potato leafroll virus isolates from the Czech Republic. Virus Genes. 2009; 39:153-155.
24. Ogawa T, Tomitaka Y, Nakagawa A, Ohshimab K. Genetic structure of a population of Potato virus Y inducing potato tuber necrotic ringspot disease in Japan; comparison with North American and European populations. Virus Res. 2008; 131:199-212.
25. Singh R, Valkonen T, Gray SM, Boonham N, Jones RAC, Kerlan C, et al. Brief Review Discussion paper: The naming of Potato virus Y strains infecting potato. Arch Virol. 2008; 153:1-13. ]]>
26. Li R, Mock R, Abad J, Hartung J, Kinard G. A reliable and inexpensive method of nucleic acid extraction for the PCR-based detection of diverse plant pathogens. J Virol Methods. 2008; 154:48-55.
]]>
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}