Resistencia sistémica inducida contra virus fitopatógenos mediada por la inoculación con la rizobacteria Bacillus spp.
Induced systemic resistance against plant viruses elicited by inoculation with rhizobacteria Bacillus spp.
Blancka Y. Samaniego-Gámez1, Arturo Reyes-Ramírez1, Oscar A. Moreno-Valenzuela2, José M. Tun-Suárez1*
1División de Estudios de Posgrado e Investigación, Instituto Tecnológico de Conkal, C.P. 97345, Conkal, Yucatán, México.
2Unidad de Bioquímica y Biología Molecular de Plantas, Centro de Investigación Científica de Yucatán A.C., Calle 43 No. 130, Colonia Chuburná de Hidalgo, C.P 97200, Mérida, Yucatán, México.
]]>
RESUMEN
Las plantas desarrollan diferentes mecanismos de defensa ante el ataque de patógenos. Después del reconocimiento del patógeno se activan respuestas que incluyen la expresión de genes asociados a defensa, una de ellas es la Resistencia Sistémica Inducida. Diversos estudios demostraron que esta resistencia es independiente de ácido salicílico (AS), pero requiere la señalización de ácido jasmónico (AJ) y etileno (ET). El ácido jasmónico y el etileno transducen los estímulos extracelulares reconocidos por receptores de la célula a un gran número de moléculas blanco que, en completa coordinación, integran respuestas intracelulares altamente específicas al estímulo externo. Este mecanismo, dentro de la planta, puede ser inducido a través de elicitores como Bacillus spp. Existe evidencia de la inducción de mecanismos de resistencia a través de Bacillus spp. contra hongos, bacterias y nematodos fitopatógenos, pero resultan escasos los estudios relacionados con la inducción de resistencia contra virus. En este artículo se analizan las principales vías de señalización de AS, AJ y ET y genes involucrados durante la interacción Bacillus-planta-virus; y se aportan conocimientos sobre los efectos de Bacillus en la reducción de la expresión de síntomas ocasionados por enfermedades de origen viral y las posibilidades de poder ser incluido en un programa de manejo sostenible de hortalizas.
Palabras clave: hortalizas, NPR1, COI1, ERF1.
ABSTRACT
Plants have developed various defense mechanisms against pathogen attack. After recognition of the pathogen, a number of responses including the expression of defense related genes are activated; one of them is the induced systemic resistance. Studies show that this resistance is independent of salicylic acid but requires jasmonic acid signaling and ethylene. Jasmonic acid and ethylene transduce the extracellular stimuli recognized by cell receptors to a large number of target molecules, which in a fully coordinated way, integrate the highly specific intracellular responses to external stimuli. Several studies have shown elicitors like Bacillus spp. to induce this mechanism with in the plant. While many studies show induction of resistance mechanisms by Bacillus spp. against fungi, bacteria, and nematodes, few studies have focused on the induction of virus resistance in the virus-plant-rhizobacteria interaction. In this paper, the main signaling pathways of salicylic acid, jasmonic acid and ethylene, and the expression of genes involved in the interaction Bacillus-plant-virus are discussed. This review contributes to a better understanding of the effects of Bacillus spp. on the reduction of viral disease symptom expression and the possibility of Bacillus of being included into a sustainable vegetable management program.
Key words: vegetables, NPR1, COI1, ERF1.
INTRODUCCIÓN
]]> Las enfermedades atribuidas a virus fitopatógenos son comunes en cultivos agrícolas y, a nivel mundial, ocasionan severas pérdidas económicas anualmente (1,2). Los virus se encuentran también en plantas nativas y constituyen reservorios potenciales para la diseminación de enfermedades virales (3,4,5).En la agricultura, el manejo de virus ha sido, principalmente, a través del control de los insectos vectores, además del uso de semillas libres de virus, la selección negativa, las medidas físicas o barreras y algunos compuestos para prevenir o limitar la infección, como benzotiadiazol, ácido acetil salicílico y quitosano (6,7,8,9). De la misma manera, el uso de plantas con genes de resistencia hacia enfermedades atribuidas a la presencia de virus, también se utiliza para controlar las pérdidas en los cultivos agrícolas (7,10). Actualmente, se informan cientos de genes de resistencia a virus en plantas monocotiledóneas y dicotiledóneas (7,10). Asimismo, esta resistencia puede ser potenciada también por elicitores como patógenos y microorganismos.
Recientemente, se produjo un aumento en el número de productos a base de microorganismos benéficos, como inductores de resistencia a enfermedades (11,12). Algunos de estos microorganismos pueden potenciar la Resistencia Sistémica Inducida (RSI), un mecanismo de defensa que las plantas desarrollan ante el ataque de patógenos.
Diversos estudios demostraron que RSI es independiente del ácido salicílico (AS), pero requiere la señalización del ácido jasmónico (AJ) y el etileno (ET), y no involucra la síntesis de proteínas relacionadas con la patogenicidad (12,13). Los productos AJ y ET transducen los estímulos extracelulares reconocidos por receptores de la célula a un gran número de moléculas blanco que, en completa coordinación, integran respuestas intracelulares altamente específicas al estímulo externo (14,15).
La RSI es potenciada por microorganismos, como las rizobacterias promotoras del crecimiento vegetal (PGPR), en la interacción planta-microorganismo (16). Las rizobacterias como Azozpirillum, Azotobacter, Gluconacetobacter, Pseudomonas y Bacillus, son utilizadas para estimular los mecanismos de RSI (15,17). Algunos estudios demostraron que las PGPR pertenecientes al género Bacillus pueden inducir mecanismos de defensa contra enfermedades producidas por virus en cultivos hortícolas (6,18). El uso de estas rizobacterias, como alternativa biológica para el control de virus en plantas, puede contribuir, en gran medida, al desarrollo de una agricultura sostenible. En este artículo se analizan las principales vías de señalización de AS, AJ y ET y genes involucrados durante la interacción Bacillus-planta-virus.
PARTE ESPECIAL ESTRATEGIAS DE PROTECCIÓN ANTIVIRAL EN PLANTAS
Los virus que infectan a las plantas son, generalmente, de tipo ADN o ARN de cadena simple. Estos fitopatógenos son transmitidos a las plantas a través de semillas, polen o vectores (3,9). El ciclo viral se inicia al penetrar el virus en la célula hospedera. Este comienza con el desensamblaje, la replicación del ARN, la traducción de proteínas, el ensamble, la liberación, el movimiento de célula a célula y a larga distancia (7). La multiplicación y translocación de los virus en las plantas son necesarios para el desarrollo de los síntomas.
A nivel mundial, para disminuir los daños ocasionados por la diseminación de los virus fitopatógenos, se emplearon cultivares resistentes que son desarrollados en los programas de mejoramiento genético (7,8). Para llevar a cabo estos programas existen tres fuentes de obtención de genes para proteger las plantas contra los virus: 1) Genes de resistencia natural, 2) Resistencia derivada del patógeno, y 3) Resistencia conferida por otras fuentes, como son protección cruzada, resistencia derivada de anticuerpos, silenciamiento postranscripcional de genes (sPtG), resistencia mediada por proteínas y péptidos (7).
Con respecto a los genes de resistencia natural, diversos estudios demostraron que la mayoría de los genes de resistencia viral caracterizados en plantas fueron de la clase dominio de unión a nucleótido de repeticiones ricas en leucina (NBS-LRR), que proporciona resistencia dominante monogénica (10).
Con relación a la resistencia conferida por otras fuentes, el mecanismo de protección cruzada se informó por primera vez a comienzos de los años 20 en el siglo pasado, en el cual la planta que ha sido inoculada con una cepa viral no tan severa puede protegerse, posteriormente, de una cepa viral más agresiva (7).
]]> Posteriormente, se desarrollaron mayores avances en el entendimiento de los mecanismos moleculares asociados a la resistencia genética a virus fitopatógenos. A finales de la década de los 90, mediante el mecanismo de silenciamiento génico mediado por ARN, se generaron diversas plantas resistentes a virus (7).Existe un extenso número de investigaciones que señalaron que las plantas perfeccionan y aumentan su capacidad defensiva contra patógenos después de un estímulo apropiado, y este mecanismo de resistencia es regulado por una red de las vías de señalización de las hormonas AS, AJ y ET que inducen la expresión de distintos conjuntos de genes (10,19).
Bajo este contexto, se demuestra que microorganismos como las rizobacterias del género Bacillus pueden inducir resistencia sistémica en plantas, mediante la expresión de genes que participan en la cascada de señalización de AS, AJ y ET contra diversos patógenos, con especial enfoque hacia enfermedades ocasionadas por virus (18).
¿CÓMO FUNCIONA LA RSI DURANTE LA DEFENSA DE LAS PLANTAS?
Para sobrevivir a enfermedades ocasionadas por diversos patógenos, las plantas desarrollan mecanismos de resistencia inducida basados en el reconocimiento del patógeno y una inmediata respuesta de señalización para activar las defensas (19).
En el reconocimiento, la interacción entre un patógeno particular y una especie de planta es específica e invariable. Se inicia la activación de las defensas de las plantas con el reconocimiento en los patógenos de las moléculas efectoras generales o específicas. Inmediatamente después de este reconocimiento comienza la respuesta de la planta con el incremento en la expresión de los genes de defensa llevándose a cabo: a) el fortalecimiento físico de las paredes celulares por la producción de ligninas y la formación de callo, b) se intensifica la producción de fitoalexinas, y c) se induce la producción de proteínas antimicrobianas como las quitinasas, β-1,3-glucanasas o peroxidasas, y las proteínas relacionadas con la patogénesis (PR) (8,20). El aumento en la expresión de los genes relacionados con la defensa está regulado por moléculas de señalización como el ET, el AS y el AJ (8,13).
Las interacciones entre las vías de señalización están mediadas por dos formas de resistencia sistémica, la Resistencia Sistémica Adquirida (RSA, mediada por AS) y la Resistencia Sistémica Inducida (RSI, mediada por AJ y ET). Las investigaciones sobre estos mecanismos de resistencia sugieren que las vías de señalización de los jasmonatos y el etileno suelen trabajar juntos de manera cooperativa, siendo generalmente antagónicos a la vía de señalización de AS (8,13).
No obstante, existe una creciente evidencia sobre que las vías de señalización mediadas por AS, AJ y ET no funcionan de manera independiente (21). Sin embargo, algunas características en las que se pueden diferenciar RSA y RSI son que, en el caso de RSA, esta es inducida por un amplio número de elicitores bióticos o abióticos, induce PR y utiliza vías de señalización que pueden involucrar al AS. Por otra parte, RSI es potencializada por la interacción planta-PGPR, no involucra la síntesis de proteínas PR y la señalización la realiza a través de AJ y ET (6,22).
VÍAS DE SEÑALIZACIÓN Y GENES INVOLUCRADOS EN LA RSI
La RSI mediada por rizobacterias puede activar mecanismos de defensa inducidos en la planta similares a la respuesta en interacciones incompatibles con microorganismos patógenos (20). Las vías de señalización del AJ y el ET actúan inmediatamente después del reconocimiento inicial entre las bacterias benéficas y la planta huésped (8).
]]> Tras la percepción del patógeno, se activan rápidamente una serie de respuestas que incluyen la expresión de genes asociados a defensa (19). Una de estas respuestas de defensa promueve la RSI a través de la síntesis y posterior activación de las rutas del AJ y ET; sin embargo, la interacción entre las vías de señalización puede ser antagónica o de cooperación, lo cual resulta en la enfermedad o resistencia en la planta hacia el patógeno (21). Si la interacción es positiva se activa una cascada de señalización que involucra a diversos genes y factores de transcripción (21,22).VÍA DE SEÑALIZACIÓN MEDIADA POR EL ÁCIDO SALICÍLICO
El AS es una molécula que posee un anillo aromático que lleva un grupo hidroxilo, o su derivado funcional, pertenece a un conjunto de compuestos fenólicos y se produce por un extenso número de organismos procariotas y eucariotas, incluyendo a las plantas (19). Las investigaciones sobre esta molécula sugieren que el AS es una molécula de señalización en la respuesta inmune de la planta, actuando durante el desarrollo de la RSA en plantas dicotiledóneas (8,19,23).
El AS en plantas se sintetiza a partir del metabolito primario corismato, que es convertida en isocorismato por acción de la enzima isocorismatosintasa. El isocorismato a su vez, es convertido en AS por acción de la enzima Isocorismato PiruvatoLiasa (24). Al recibir la señalización, el AS inicia la cascada de señalización de la resistencia sistémica a través de la activación de NPR1, mismo que en estado inactivo está presente en un complejo oligómero citosólico a través de enlaces disulfuro intermoleculares (25). Al iniciar la vía de señalización mediada por el AS, este activa a la tiorredoxina TRX-H5, conduciendo a la reducción en NPR1; por lo tanto, la conversión a monómeros activos que se translocan desde el citosol al núcleo, activan la expresión del resto de genes de defensa (25,26).
VÍA DE SEÑALIZACIÓN DEL ETILENO
La señalización de defensa regulada por ET incluye receptores de dicha molécula, que son capaces de activar una cascada de quinasas proteínas quinasas activadas por mitógenos (MAPK), cuya señal es transmitida al gen insensitivo a etileno-2 (EIN2) y que permite la acumulación del factor de transcripción del gen insensitivo a etileno-3 (EIN3) y promueve al factor de transcripción de respuesta a etileno (ERF1), el mismo que regula la expresión génica en respuesta a patógenos y a la represión de genes de respuesta a herida (24,27).
El factor de transcripción ERF está posicionado en el penúltimo paso en la cascada de señalización de etileno, directamente regulada por la expresión del gen PR, en virtud de la unión de ADN con el codón CCG (GCCGCC) situado en los promotores de genes PR, tales como PRB-1b (PR1), β-1, 3-glucanasa (PR2), quitinasa (PR3), y osmotina (PR5) (22). El gen ERF1 se induce, sinérgicamente, por las vías de señalización del AJ y ET, y la inducción por cualquiera de estas hormonas es dependiente de una vía de transducción de señal para ambas hormonas, lo que indica que en ERF1 puede haber un punto de integración para AJ y ET para la regulación de los genes de defensa en respuesta a los patógenos (13).
Utilizando el modelo Arabidopsis, se observó que la función del etileno fue fundamental frente a la infección ocasionada por el Cauliflower mosaic virus (CaMV) (28). De igual manera, un estudio en mostaza demostró que el etileno, en interacción con la ruta de la oxidasa alternativa mitocondrial (AOX), induce resistencia sistémica al Turnip mosaic virus (TuMV), posiblemente al limitar la infección sistémica del virus y su acumulación en la planta (27).
VÍA DE SEÑALIZACIÓN DEL ÁCIDO JASMÓNICO
La síntesis de jasmonatos ocurre, principalmente, en las hojas, donde los ácidos grasos son liberados de la membrana plasmática de los plastidios por acción de enzimas fosfolipasas y desaturasas. La señalización por jasmonatos, es esencial para la expresión de genes de defensa en plantas contra las plagas (29). Se observó que el gen de la proteína insensible a la coronatina (COI1) es el componente regulador central de la vía de señalización del ácido jasmónico y se requiere para las respuestas de defensa de las plantas (22). La unión de Jasmonil-Isoleucina (JA-Ile) al receptor COI1, en el núcleo celular, promueve la unión de los represores de jasmonatos Dominio-ZIM (JAZ) y su ubiquitinación, resultando así en la liberación del factor de transcripción MYC2 y la transcripción de genes de respuesta a jasmonatos (29).
]]> En la primera década del siglo XXI, se realizaron investigaciones para conocer la expresión del gen COI1 en plantas infectadas con virus fitopatógenos. Particularmente, se observó la expresión de este gen en la interacción tabaco-TMV, al igual que la expresión del gen N en plantaciones de tabaco, que confiere resistencia al TMV; se observó que la supresión del gen COI1 afecta la función del gen N y COI1 es un componente en la respuesta de la resistencia a TMV (30). Recientemente, en plantas de tabaco infectadas con TMV e inoculadas con Bacillus spp. se observó la activación de los genes de NtPR1, NtNPR1 y NtCOI1, generando una RSI modulada por Bacillus spp. (22).INTERACCIÓN ENTRE LAS VÍAS DE SEÑALIZACIÓN DE AJ/ET Y AS
En la última década, diversos estudios demostraron que las interacciones entre la ruta del ácido salicílico y la ruta del jasmonato y etileno pueden tener una cooperación antagónica o una cooperación positiva mutua. A pesar de ello, la cooperación antagónica no es la más común, pero existen algunos informes de dicha actividad (13,21). Por lo tanto, se ha sugerido que la planta activa la red de señalización en dependencia de la naturaleza del patógeno y de su modo de patogenicidad (19). Al respecto, estudios señalan al gen NPR1 como regulador central en la ruta de señalización, tanto de RSA como de RSI (12,22).
El gen NPR1 codifica para una proteína que se transporta al núcleo en la célula durante la respuesta RSA, donde actúa como modulador de la expresión de los genes PR (4). Allí, los cambios redox, que son inducidos como resultado de la acumulación del AS, conectan la señal del AS con NPR1. Este gen interactúa con un grupo de miembros de la familia de tipo cierre de leucina de región básica (bZIP), tipo factores de transcripción (TGAs), que a su vez se unen, específicamente, a los sitios promotores de genes de respuesta al AS como PR-1 (11,26).
El rol del gen NPR1 podría ser diferente en la RSA que en la RSI. Sin embargo, estas diferentes funciones de NPR1 no son mutuamente excluyentes, ya que la simultánea activación de la RSA y la RSI puede aumentar la capacidad de defensa de la planta, comparado con su activación de forma independiente (19). En la ruta de AS, el NPR1 se conecta a una proteína del núcleo, mientras que en la ruta mediada por AJ/ET la proteína es conectada al citosol. Sin embargo, los mecanismos moleculares mediante los cuales el NPR1 realiza esta función en la ruta AJ/ET aún no son muy claros (19,26).
CARACTERÍSTICAS DE Bacillus spp.
Las bacterias pertenecientes al género Bacillus forman parte del nicho ecológico del suelo. Bacillus spp. son bacterias Gram-positivas, aerobios o anaerobios facultativos, catalasa positiva y poseen ciertas características metabólicas que permiten su supervivencia en el suelo, como la presencia de una pared celular con múltiples capas y la formación de endosporas resistentes a factores adversos del ambiente (18).
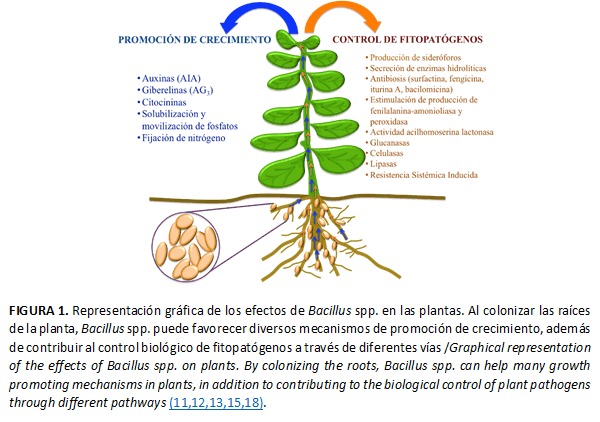
Al colonizar las raíces de la planta, Bacillus spp. estimulan diversos mecanismos de promoción de crecimiento a través de la síntesis de hormonas; aunado a ello, puede favorecer la fijación de nitrógeno, movilización de fosfatos y fosfatasas (16,18).
A su vez, Bacillus spp. puede inducir la producción de metabolitos y enzimas, y la Resistencia Sistémica en la planta(11,12,13,17,18). Las especies de Bacillus son rizobacterias cosmopolitas; sin embargo, diversos estudios indican que las secuencias de las especies de Bacillus dominantes presentes en un suelo, son diferentes a las presentes en las cepas cultivadas en medios sintéticos (11). Algunas de estas especies, como Bacillus subtilis, Bacillus polymyxa, Bacillus pumilus, Bacillus amyloliquefaciens, Bacillus cereus y Bacillus licheniformis se utilizan comercialmente como parte del control biológico hacia enfermedades en cultivos agrícolas (12,13,31).
Aunque un gran número de investigaciones se han utilizado para demostrar la actividad antagónica de Bacillus spp. hacia bacterias fitopatógenas (31,32), oomicetos (33) y hongos fitopatógenos (34,35,36,37,38), recientemente se evaluó su efecto como promotoras de RSI hacia virus fitopatógenos (13).
]]> INDUCCIÓN DE RSI POR Bacillus spp.La colonización de las plantas por las PGPR genera un aumento en su productividad y sanidad al favorecer la absorción de nutrientes, el crecimiento de la planta hospedante y el aumento de la estimulación de las defensas hacia patógenos y plagas, como método de control biológico (11,39).
Estas PGPR contribuyen al control biológico de enfermedades mediante diferentes mecanismos de acción: competencia por un nicho ecológico o sustrato, la formación de compuestos inhibitorios como antibióticos, producción de sideróforos, enzimas detoxificadoras y líticas, así como la RSI a fitopatógenos. Esta protección puede incrementarse al realizar la mezcla de dos o más cepas bacterianas, aumentando su rango de acción a varios hospedantes y contra diferentes fitopatógenos (18). Bacillus spp. son un ejemplo de las PGPR ampliamente utilizadas para el control biológico (Figura 1). Es conocido el efecto de Bacillus spp. como elicitores de RSI en cultivos agrícolas, tales como tabaco, pepino, sandía, jitomate y chile contra enfermedades ocasionadas por hongos, nematodos, bacterias y virus (18). De las especies de Bacillus, se informó a B. subtilis y B. cereus por su capacidad de inducción de RSI hacia fitopatógenos (11).
Después de la colonización de Bacillus spp. en las raíces, se demostró que las cepas inducen un aumento en la formación de auxinas, que resulta en la formación de abundantes raíces laterales, el aumento de la longitud del pelo de la raíz y el aumento de la producción de biomasa de las plantas (13).
Además de promover el crecimiento, B. subtilis también induce resistencia sistémica en las plantas hospedantes a través de la liberación de compuestos volátiles como acetoína y 2,3-butanodiol (7). Tras la activación de RSI, se ha observado una aumento en la producción de peroxidasa, una enzima antioxidante presente en las plantas, así como la activación de ciertos genes de defensa de plantas que codifican para las proteínas fenilalanina amonio liasa (PAL) y PR, y quitinasa, β-1, 3 glucanasa (40). Sin embargo, a pesar de que la mayoría de los estudios sobre la activación de RSI están dirigidos hacia diversos fitopatógenos, ha habido algunos estudios sobre la RSI hacia virus fitopatógenos (11).
MECANISMOS DE INDUCCIÓN DE RSI POR Bacillus spp. CONTRA VIRUS FITOPATÓGENOS
El uso de PGPR para el control de enfermedades basado en la RSI se incrementó en los últimos años, las investigaciones sobre la aplicación de rizobacterias para el control de enfermedades ocasionadas por virus fitopatógenos han sido reducidas (22,41). La particularidad del uso de Bacillus spp. es en relación con la activación de algunos mecanismos de RSI hacia virus fitopatógenos (Tabla 1).
Se han empleado hospedantes pertenecientes a las familias Cucurbitaceae, Solanaceae, Fabaceae, Amaranthaceae y Brassicaceae, infectados con virus de las familias Potyviridae, Bromoviridae, Virgaviridae y Geminiviridae, donde se ha evaluado el efecto de la inoculación de B. pumilus, B. subtilis, B. amyloliquefasciens y B. cereus, aplicadas de manera individual y en mezclas, en condiciones experimentales in vitro, de campo e invernadero. El efecto de Bacillus spp. hacia enfermedades ocasionadas por virus fitopatógenos, solo había sido evaluado a nivel de expresión de síntomas; sin embargo, recientemente se implementaron técnicas moleculares para deducir los mecanismos atribuidos a este fenómeno (22).
A nivel de síntomas, se observó que la inoculación en plantas con Bacillus spp. e infectadas con el virus ocasiona una reducción en la incidencia y severidad, además de un aumento en la altura y el peso fresco de las plantas. Este efecto puede ser atribuido a la RSI mediada por Bacillus spp., a la inhibición de la replicación viral o a su dilución por efectos del crecimiento de la planta por efecto de la inoculación (41). En algunos de estos estudios, el efecto de la inoculación de cepas de Bacillus spp. se evaluó a campo abierto y en condiciones de invernadero; se observó una reducción en la severidad e incidencia de la enfermedad a los 28 y 40 días después de la inoculación con TMV y el Tomato mottle virus (ToMoV), incluso al segundo año de producción, contra el Cucumber mosaic virus (CMV), atribuyendo dicha reducción de la enfermedad a la inducción de RSI por la inoculación con cepas Bacillus spp. en las plantas (15,22,41).
En otro estudio contra CMV se encontró que, al segundo año de producción, los niveles de incidencia y severidad fueron más altos que los observados en el primer año. Esta disminución del efecto de la inoculación con Bacillus spp. puede ser atribuida a la alta presión de inóculo de CMV, además de que las plantas pudieron haber sido infectadas de manera natural con cepas diferentes de CMV, contra las que Bacillu sspp. no fue efectiva (42). Esta variabilidad en el efecto de la inoculación con Bacillu sspp. podría sugerir que la inducción depende de la cepa de la bacteria, la planta hospedante y, en el último de los casos, del patógeno usado en el hospedante específico (11,18). Además, esta resistencia inducida es una respuesta del hospedante y, como tal, está en gran medida influenciada por el genotipo y el medio ambiente (8). Sin embargo, resulta importante detectar la presencia del virus y determinar la acumulación viral en la planta para validar el efecto de Bacillus spp. en las interacciones rizobacteria-planta-virus. Existen métodos confiables para la cuantificación de las partículas virales, como la técnica de Ensayos de Inmunoabsorción Ligados a Enzimas (ELISA), los análisis Western Blot y Southern Blot, la Reacción en Cadena de la Polimerasa-cuantitativa (qPCR), también conocida como Reacción en Cadena de la Polimerasa en tiempo real (PCR-real time) (4,22,23).
]]> Los estudios resaltan que las esporas de Bacillus spp. colonizan las plantas y se mantienen, mayormente, adheridas a la superficie de la raíz y, con menor frecuencia, en la parte inferior del tallo y en las hojas de las plantas (22,46). Después de haber sido expuesta a la asociación raíz-Bacillus spp. las plantas tienden a aumentar sus niveles de resistencia a patógenos, a través de la RSI (11). Investigaciones realizadas en plantas de tabaco inoculadas con Bacillus spp. e infectadas con TMV demostraron la activación de los genes COI1, NPR1, PR-1a y PR-1b (22). En otros estudios se observó que las plantas de tabaco tratadas con Bacillus spp., e infectadas con TMV, presentaron reducción en los niveles de incidencia y severidad, cambios en la acumulación viral, además de un aumento en la producción de PR, de las enzimas peroxidasa, polifenoloxidasa y fenilalanina amonoliasa (46). Estos cambios fisiológicos y bioquímicos que manifiestan las plantas sugieren que la inoculación con especies de Bacillus puede protegerlas contra la infección ocasionada por el virus, al aumentar el fortalecimiento de la pared celular y al restringir la invasión del virus mediante el aumento de resistencia sistémica de la planta y de la acumulación de metabolitos secundarios (22,46).
CONCLUSIONES
En la última década las investigaciones sobre los mecanismos de defensa inducidos por microorganismos contra fitopatógenos han ido en aumento. En estos trabajos se demuestra el efecto de la rizobacteria Bacillus en la interacción con las plantas, principalmente como promotor de crecimiento, y su uso en el control biológico de enfermedades. Sin embargo, el número de informes acerca de la inducción de resistencia contra virus fitopatógenos es reducido. Se observó que, tras la detección del virus, la planta promueve una serie de mecanismos que interactúan entre sí para responder al ataque, como son los relacionados con las vías de señalización del ácido salicílico, ácido jasmónico y etileno. En conjunto, el ácido jasmónico y el etileno inician la cascada de señalización para la resistencia sistémica inducida por la inoculación con Bacillus spp.
El descubrimiento de los genes involucrados en las vías de señalización ha sido un factor determinante para conocer el efecto de Bacillus spp. en la resistencia sistémica inducida. La expresión de genes ERF1, COI1 y NPR1 en las vías de señalización mediadas por ácido jasmónico y etileno puede ser inducida por la presencia de cepas de Bacillus spp.; sin embargo, este fenómeno depende de la cepa, la planta hospedante y el patógeno utilizado en un hospedante determinado. Aunado a la inducción de los mecanismos de resistencia en la planta infectada con virus, se menciona que Bacillus spp. puede participar en inhibición en la replicación viral o en su dilución por efectos de crecimiento de la planta. Las rizobacterias como Bacillus, mediante la inducción de resistencia en las plantas, son una alternativa con potencial para el desarrollo de bioproductos para el manejo sostenible de enfermedades ocasionadas por virus fitopatógenos.
AGRADECIMIENTOS
Agradecemos al CONACYT por la beca para realizar estudios de posgrado de Blancka Y. Samaniego-Gámez. De igual manera, nuestro agradecimiento al Dr. Raúl Enrique Valle-Gough, al Dr. Hernán Villanueva-Alonzo, y al Dr. Esaú Ruiz-Sánchez por la revisión de este escrito.
REFERENCIAS ]]>
1. Gottula J, Lewis R, Saito S, and Fuchs M. Allopolyploidy and the evolution of plant virus resistance. BMC Evolutionary Biology. 2014;14:149.
2. Zamudio-Moreno E, Ramírez-Prado JH, Moreno-Valenzuela OA, and López-Ochoa LA. Early diagnosis of a Mexican variant of Papaya meleira virus (PMeV-Mx) by RT-PCR. Genetics Molecular and Research. 2015;14(1):1145-1154.
3. Tun Suárez JM, Zavaleta Mejía E, Ochoa Martínez DL, Sánchez García P, Soto Hernández M, Cristóbal Alejo J. Incidencia del Virus Mosaico de la Alfalfa en semillas y plántulas de chile habanero (Capsicum chinense Jacq.) en Yucatán, México. Fitosanidad. 2007;11(1):11-14.
4. Villanueva-Alonzo HJ, Us-Camas RY, López-Ochoa LA, Robertson D, Guerra-Peraza O, Minero-García Y, Moreno-Valenzuela OA. A new virus-induced gene silencing vector based on Euphorbia mosaic virus-Yucatan peninsula for NPR1 silencing in Nicotiana benthamiana and Capsicum annuum var. Anaheim. Biotechnology Letters. 2013;35:811–823.
5. Hernández-Zepeda C, Idris AM, Carnevali G, Brown JK, Moreno-Valenzuela OA.b. Preliminary identification and coat protein gene phylogenetic relationships of begomoviruses associated with native flora and cultivated plants from the Yucatan Peninsula of Mexico. Virus Genes. 2007;35:825–833.
]]>
6. Maldonado-Cruz E, Ochoa-Martínez DL, Tlapal-Bolaños B. Efecto del ácido acetil salicílico y Bacillus subtilis en la infección causada por Cucumber mosaic virus en calabacita. Revista Chapingo Serie horticultura. 2008;14(1):55-59.
7. Kobayashi KK, Sekine KT, Nishiguchi M. Breakdown of plant virus resistance: can we predict and extend the durability of virus resistance? Journal of General Plant Pathology. 2014;80(4):327-336.
8. Faoro F, Gozzo F. Is modulating virus virulence by induced systemic resistance realistic? Plant Science. 2015;234:1-13.
9. Hamelin FM, Allen LJS, Prendeville HR, Reza Hajimorad M, Jeger MJ. The evolution of plant virus transmission pathways. Journal of Theoretical Biology. 2016;396(7):75-89.
10. Gururani MA, Venkatesh J, Upadhyaya CP, Nookaraju A, Pandey SK, Park SW. Plant disease resistance genes: Current status and future directions. Physiological and Molecular Plant Pathology. 2012;78:51-65.
]]>
11. Choudhary DK, and Johri BN. Interactions of Bacillus spp. and plants – With special reference to induced systemic resistance (ISR). Microbiological Research. 2009;164(5):493-513.
12. Beneduzi A, Ambrosini A, Passaglia LMP. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genetics and Molecular Biology. 2012;35(4 Suppl):1044-1051.
13. Pieterse CMJ, Zamioudis C, Berendsen RL, Weller DM, Van Wees SCM, Bakker PAHM. Induced Systemic Resistance by Beneficial Microbes. Annual Review of Phytopathology. 2014;52:347–75.
14. Jankiewicz U, Koltonowicz M. The involvement of Pseudomonas bacteria in Induced Systemic Resistance in Plants (Review). Applied Biochemistry and Microbiology. 2012;48(39):244-249.
15. Choi HK, Song GC, Yi HS, Ryu CM. Field Evaluation of the Bacterial volatile derivative 3-Pentanol in priming for induced resistance in Pepper. Journal of Chemical Ecology. 2014;40(8):882-92.
]]>
16. Elsharkawy MM, Shimizu M, Takahashi H, Hyakumachi M. Induction of systemic resistance against Cucumber mosaic virus by Penicillium simplicissimum GP17-2 in Arabidopsis and tobacco. Plant Pathology. 2012;61(5):964–976.
17. Guo S, Li X, He P, Ho H, Wu Y, He Y. Whole-genome sequencing of Bacillus subtilis XF-1 reveals mechanisms for biological control and multiple beneficial properties in plants. Journal of Industrial Microbiology and Biotechnology. 2015;42(6):925-937.
18. Kloepper JW, Ryu CM, Zhang S. Induced systemic resistance and promotion of plant growth by Bacillus spp. Phytopathology. 2004;94(11):1259-1266.
19. Thaler JS, Humphrey PT, Whiteman NK. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Science. 2012;17(5):260-270.
20. García GL, Zeriouh H, Romero D, Cubero J, de Vicente A, Pérez GA. The antagonistic strain Bacillus subtilis UMAF6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate- and salicylic acid-dependent defense responses. Microbial Biotechnology. 2013;6(3):264-74.
]]>
21. Derksen H, Rampitschb C, Daayf F. Signaling cross-talk in plant disease resistance. Plant Science. 2013;207:79-87.
22. Wang S, Wu H, Qiao J, Ma L, Liu J, Xia Y, Gao X. Molecular mechanism of plant growth promotion and induced systemic resistance to Tobacco Mosaic Virus by Bacillus spp. Journal of Microbiology and Biotechnology. 2009;9(10):1250-1258.
23. Luna‐Rivero MS, Hernández‐Zepeda C, Villanueva‐Alonzo H, Minero‐García Y, Castell‐González SE, Moreno‐Valenzuela OA. Expression of genes involved in the salicylic acid pathway in type h1 thioredoxin transiently silenced pepper plants during a begomovirus compatible interaction. Molecular Genetics and Genomics. 2016;291(2):819–830.
24. Kepinski S, Leyser O. SCF-Mediated Proteolysis and Negative Regulation in Ethylene Signaling. Cell. 2003;115(6):647-8.
25. Brosché M, Kangasjärvi J. Low antioxidant concentrations impact on multiple signalling pathways in Arabidopsis thaliana partly through NPR1. Journal of Experimental Botany. 2012;63(5):1849-1861.
]]>
26. Tada Y, Spoel SH, Pajerowska-Mukhtar K, Mou ZL, Song JQ, Wang C, Zuo JR, Dong XN. Plant immunity requires conformational charges of NPR1 via S-nitrosylation and thioredoxins. Science. 2008;321(5891):952–956.
27. Zhu L, Li Y, Li L, Yang J, Zhang M. Ethylene is involved in leafy mustard systemic resistance to Turnip mosaic virus infection through the mitochondrial alternative oxidase pathway. Physiological and Molecular Plant Pathology. 2011;76(3-4):166-172.
28. Love AJ, Laval V, Geri C, Laird J, Tomos AD, Hooks MA. Components of Arabidopsis defense- and ethylene-signaling pathways regulate susceptibility to Cauliflower mosaic virus by restricting long-distance movement. Molecular Plant Microbe Interaction. 2007;20(6):659-70.
29. Garnica VA, Raya GJ, López BJ, Beltrán PE. Señalización del ácido jasmónico y el óxido nítrico durante el desarrollo de Arabidopsis thaliana. Ciencia Nicolaita. 2012;55:28-41.
30. Liu Y, Schiff M, Dinesh KSP. Involment of MEK1 MAPKK, NTF6 MAPK, WRKY/MYB transcription factors, COI1 and CTR1 in N-mediated resistance to tobacco mosaic virus. The Plant Journal. 2004;38(5):800-809.
]]>
31. Berić T, Kojić M, Stanković S, Topisirović L, Degrassi G, Myers M, Venturi V, Fira D. Antimicrobial activity of Bacillus sp. natural isolates and their potential use in the biocontrol of phytopathogenic bacteria. Food Technology and Biotechnology. 2012;50(1):25–31.
32. Zeriouh H, Romero D, Garcia-Gutierrez L, Cazorla FM, de Vicente A, Perez-Garcia A. The Iturin- like lipopeptides are essential components in the biological control arsenal of Bacillus subtilis against bacterial diseases of cucurbits. Molecular Plant Microbe Interactions. 2011;24(12):1540–1552.
33. Solanki MK, Robert AS, Singh RK, Kumar S, Pandey AK, Srivastava AK, Arora DK. Characterization of mycolytic enzymes of Bacillus strains and their bio-protection role against Rhizoctonia solani in tomato. Current Microbiology. 2012;65(3):330–336.
34. Reyes Ramírez A, Ruiz Sánchez E, Yam Chimal C, Dzul Chan M. Selección de Bacillus spp. con actividad antagónica in vitro contra Macrophomina phaseolina (Tassi) Goid. en diferentes medios de cultivo. Fitosanidad. 2011;15(2):117-121.
35. Rodas-Junco BA, Magaña-Sevilla HF, Tun-Suarez JM and Reyes-Ramirez A. Antifungal activity in vitro of native Bacillus sp. strains against Macrophomina phaseolina (Tassi) Goid. Research Journal of Biological Sciences. 2009;4(9):985-989.
]]>
36. Ruiz-Sánchez E, Mejía-Bautista MA, Cristóbal-Alejo J, Valencia-Botín A, Reyes-Ramírez A. Actividad antagónica de filtrados de Bacillus subtilis contra Colletotrichum gloeosporioides. Revista Mexicana de Ciencias Agrícolas. 2014;5(7):1325-1332.
37. Ruiz-Sánchez E, Mejía-Bautista MA, Serrato-Díaz A, Reyes-Ramírez A, Estrada-Girón Y, Valencia-Botín AJ. Antifungal activity and molecular identification of native strains of Bacillus subtilis. Agrociencia. 2016;50(2):133-148.
38. Sosa-Pech M, Ruiz-Sánchez E, Mejía-Bautista MA, Reyes-Ramírez A, Cristóbal-Alejo J, Valencia-Botín A, Gutiérrez-Alonzo O. Actividad antagonista in vitro de aislados de la clase Bacilli de la Península de Yucatán contra cuatro hongos fitopatógenos. Universidad y Ciencia. 2012;28(3):279-284.
39. Wang S, Wu H, Zhan J, Xia Y. The role of synergistic action and molecular mechanism in the effect of genetically engineered strain Bacillus subtilis OKBHF in enhancing tomato growth and Cucumber mosaic virus resistance. Bio Control. 2011;56(1):113-121.
40. Mishra S, KS Jagadeesh, PU Krishnaraj, S Prem (2014) Biocontrol of tomato leaf curl virus (ToLCV) in tomato with chitosan supplemented formulations of Pseudomonas sp. under field conditions. Austalian Journal of Crop Science. 2014;8(3):347-355.
]]>
41. Murphy JF, Zehnder GW, Schuster DJ, Sikora EJ, Polston JE, Kloepper JW. Plant growth-promoting rhizobacterial mediated protection in tomato against Tomato mottle virus. Plant Disease. 2000;84(7):779-784.
42. Zehnder GW, Yao C, Murphy JF, Sikora ER, Kloepper JW. Induction of resistance in tomato against cucumber mosaic cucumovirus by plant growth-promoting rhizobacteria. BioControl. 2000;45(1):127-137.
43. Lee, G. H., Ryu, C.-M. Spraying of leaf-colonizing Bacillus amyloliquefaciens protects pepper from Cucumber mosaic virus. Plant Disease. 2016;100(10):2099-2105.
44. Elbeshehy EKF, Youssef SA, Elazzazy AM. Resistance induction in pumpkin Cucurbita maxima L. against Watermelon mosaic potyvirus by plant growth-promoting rhizobacteria. Biocontrol Science and Technology. 2015;25(5):525-542.
45. El-Borollosy AM, and Oraby MM. Induced systemic resistance against Cucumber mosaic cucumovirus and promotion of cucumber growth by some plant growth-promoting rhizobacteria. Annals of Agricultural Science. 2012;57(2):91-97.
]]>
46. Lian L, Xie L, Zheng L, Lin Q. Induction of systemic resistance in tobacco against Tobacco mosaic virus by Bacillus spp. Biocontrol Science and Technology. 2011;21(3):281-292.
47. Shankar ACU, Nayaka SC, Niranjan-Raj S, Kumar HB, Reddy MS, Niranjanaa SR, Prakash HS. Rhizobacteria-mediated resistance against the blackeye cowpea mosaic strain of bean common mosaic virus in cowpea (Vigna unguiculata). Pest Management Science. 2009;65(10):1059-1064.
48. Zhou W, Zhang L, Zhang B, Wang F, Liang Z, Niu T. Isolation and characterization of ZH14 with antiviral activity against Tobacco mosaic virus. Canadian Journal of Microbiology. 2008;54(6):441-449.
49. Choong-Min R, Murphy JF, Mysore KS, Kloepper JW. Plant growth-promoting rhizobacteria systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. Plant Journal. 2004;39(3):381-392.
50. Murphy JF, Reddy MS, Ryu CM, Kloepper JW, Li R. Rhizobacteria-mediated growth promotion of tomato leads to protection against cucumber mosaic virus. Phytopathology. 2003;93(10):1301-1307.
Recibido: 6/7/2016
Aceptado: 17/1/2017
*Autor para correspondencia: José M. Tun-Suárez. E-mail: jose.tun@itconkal.edu.mx
]]>{kind=link}
{kind=link}