El ácido siálico N-acetilado es inmunogénico e induce anticuerpos protectores contra Neisseria meningitidis.
De-N-acetylated sialic acid is immunogenic and elicits antibodies that are protective against Neisseria meningitidis.
Gregory R. Moe*, Becca A. Flitter, Jessica Y. Ing, Tamara S. Bhandari+, and Hardeep Kaur
Center for Immunobiology and Vaccine Development, Children's Hospital & Research Center Oakland, Oakland, CA 94609
Current address: +Department of Biomedical Sciences, University of California, San Diego, San Diego, CA.
*Corresponding author: Mailing address: 5700 Martin Luther King Jr. Way, Oakland, CA 94609
Abstract
Recently, we showed that monoclonal antibodies (mAbs) that are reactive with derivatives of polysialic acid containing de-N-acetylated neuraminic acid (Neu) residues are protective against N. meningitidis group B strains (Moe et al. 2005, Infect Immun 73:2123; Flitter et al., in preparation). In addition, we found that fully de-N-acetylated PSA (i.e. poly alpha 2,8 Neu) conjugated to tetanus toxoid (DeNAc) elicits IgM and IgG antibodies of all subclasses in mice that bind to group B strains, activate human complement deposition, are protective in an infant rat model of meningococcal bacteremia and are bactericidal against group C strains (Moe et al, in press). We show here that anti-DeNAc mAbs, DA1 and DA2 (both IgM), are reactive with polysaccharides containing Neu, bind to group B, C, W135 and Y but not X strains grown in chemically defined media (CDM). However, when the group X strain is grown in CDM supplemented with human plasma, DA2 binds. Also both mAbs mediate bactericidal activity against B, C, W135, and X strains with human complement. The esults suggests that N. meningitidis express and/or acquire zwitterionic de-N-acetyl sialic acid antigens that can be the target of protective antibodies.
Keywords: Neisseria meningitidis, polysialic acid, de-N-acetyl sialic acid.
Introduction
Currently, there is no vaccine that can prevent disease caused by all strains of Neisseria meningitidis group B bacteria (MenB). Group B strains express a capsular polysaccharide consisting of poly alpha 2,8 N-acetyl neuraminic acid (poly 2,8 Neu5Ac), which is chemically identical to polysialic acid (PSA) expressed in human tissues.
Recently, we showed that monoclonal antibodies (mAbs) produced in mice by immunization with PSA derivatives containing de-N-acetyl neuraminic acid residues (NeuPSA) conjugated to a carrier protein are protective against MenB but do not cross-react with human PSA antigens (1).
]]> In the present study, we show that the murine mAbs, DA1 and DA2 (IgM) produced using a vaccine consisting of completely de-N-acetylated PSA conjugated to tetanus toxoid (DeNAc) bind to MenB, C, W135, X, and Y strains and are bactericidal with human complement against MenB, C, W135, and X strains. The mAbs have different fine antigenic specificities but both are reactive with a variety of glycans that contain Neu.The results suggest that most meningococcal strains regardless of capsular group express glycans containing Neu and that vaccines eliciting such antibodies may protect against meningococcal disease without eliciting antibodies that are cross-reactive with human PSA antigens.
Materials and methods
Preparation of de-N-acetylated colominic acid-tetanus toxoid conjugate vaccine (DeNAc)
Colominic acid (100 mg, Sigma-Aldrich, Saint Louis, MO) was de-N-acetylated in 2M NaOH at 100°C in the presence of sodium borohydride (10 mg) 6hrs. The pH as adjusted to 8 with 2M HCl, the supernatant solution was dialyzed against water (1kDa Spectrum Spectra/Por* 7 dialysis membrane; Fisher Scientific) and lyophilized.
De-N-acetylated colominic acid was oxidized with sodium periodate (5 µmol) and 20 mg of polysaccharide (approximately 1 equivalent of periodate for every 10 residues) in 0.1M sodium acetate buffer, pH 6.5. Ethylene glycol (100 µl of a 10% (v/v) solution in water) was added to destroy any remaining periodate and the solution was dialyzed and lyophilized as described above.
Oxidized de-N-acetylated colominic acid (10 mg) was conjugated to tetanus toxoid (TT, 5 mg) by reductive amination in PBS buffer with 5 mg of sodium cyanoborohydride (Sigma-Aldrich). The vaccine preparation was dialyzed (15kDa cutoff) against PBS buffer, sterile filtered (0.22 µm filter), aliquoted and stored at -80 °C until used.
Production of the mAbs. A female CD1 mouse (6-8 wk old, Charles River Laboratories, Wilmington, MA) was immunized with three doses of DeNAc (2 µg of de-N-acetyly sialic acid) as described previously (1). Three days after the final dose, the mouse was sacrificed and hybridomas were produced as described previously (2). All animal experiments were approved by the Institutional Animal Care and Use Committee. MAbs were selected based on reactivity with poly de-N-acetyl colominic acid-BSA conjugate in ELISA and ability to activate complement.
Binding Assays. Antibody binding to meningococcal strains was measured by flow cytometry as described previously (2) except that the bacteria were cultured in chemically defined media (CDM) or CDM supplemented with 5% human plasma from a donor who lacks bactericidal antibodies against the test strain and has been depleted of IgG. FITC-conjugated anti-mouse IgM, G, and A (Zymed, South San Francisco, CA) or IgG subtype-specific FITC-conjugated anti-mouse IgG1, IgG2a, IgG2b, and IgG3 (Bethyl Laboratories, Montgomery, TX) were used to detect bound antibodies. Positive control anticapsular mAbs were a gift from Dr. Jo Anne Welsch of CHORI and the negative control IgM was from Southern Biotech (Birmingham, AL).
]]> Serum bactericidal assay (SBA). The ability of the antisera to mediate bacteriolysis in the presence of exogenous human complement was measured by the serum bactericidal assay as described previously (2) except that Dulbecco's buffered saline was used instead of Gey's buffer. In one experiment testing DA2, the group W135 strain M9262 was grown in CDM supplemented with 5% heat-inactivated human plasma from a donor that lacks intrinsic bactericidal activity against the test strain and was depleted of IgG (>99% removed) by Protein G affinity chromatography. SBA was evaluated in plasma from the same donor (65% v/v) with or without leukocytes. The mAbs were purified by ammonium sulfate precipitation, dialysis, and size exclusion chromatography (ToyoPerl HW65F, Sigma-Aldrich, Saint Louis, MO). The mAbs were eluted in a buffer consisting of 2 mM arginine, pH 7, containing 20 mM sucrose and 0.0001% (v/v) Tween 20. Fractions containing IgM were combined, lyophilized and stored at -80°C until used. Antibody concentrations after resuspending the lyophilized mAb water were determined by capture ELISA (Southern Biotech).Results and discussion
The chemical structure of MenB capsular polysaccharide (MBPS), poly alpha 2,8 N-acetyl neuraminic acid is identical to human polysialic acid (PSA) and E. coli K1 capsular polysaccharide (colominic acid). MBPS is poorly immunogenic in mice even when conjugated to a carrier protein. However, fully de-N-acetylated colominic acid (i.e. poly alpha 2,8 neuraminic acid) conjugated to tetanus toxoid (DeNAc) elicited high titers of IgM and IgG antibodies of all subclasses in mice that bind to group B strains, activate human complement deposition, and are protective in an infant rat model of meningococcal bacteremia (1). In addition, the anti-DeNAc serum was bactericidal against a group C strain (1). One possible explanation for the difference is that zwitterionic polysaccharides have been shown to be processed and presented on MHC II molecules and, therefore, may stimulate helper T cells (3, 4). In contrast, sialylated glycans attenuate the activity of B cell receptors (BCRs) by interacting with a negative regulator of BCR signaling, CD22 (5). Importantly, the DeNAc antiserum was not reactive by ELISA with MBPS (1), showing that the DeNAc vaccine did not elicit autoreactive anti-PSA antibodies. The lack of cross-reactivity is not surprising since de-N-acetylated colominic acid is chemically (zwitterionic versus anionic), physically (solution structure and aggregation properties), and immunologically (1) quite different than PSA.
To investigate further the expression of de-N-acetyl neuraminic acid (Neu)-containing antigens by meningococcal strains from other capsular groups under various culture conditions and the potential for anti-Neu antigen antibodies to protect against disease, we produced mAbs using the DeNAc vaccine.
The mAbs DA1 and DA2 that were elicited by the DeNAc vaccine were selected for the studies described here because they showed reactivity in ELISAs with the nominal de-N-acetyl colominic acid antigen and several other Neu-containing polysaccharides. Also, as described below, the mAbs exhibited SBA against multiple meningococcal strains from different capsular groups with human complement. In retrospect, however, the screening method, which was based on reactivity with the nominal DeNAc antigen and ability to activate human complement turned out to be poorly conceived as only IgM mAbs were selected.
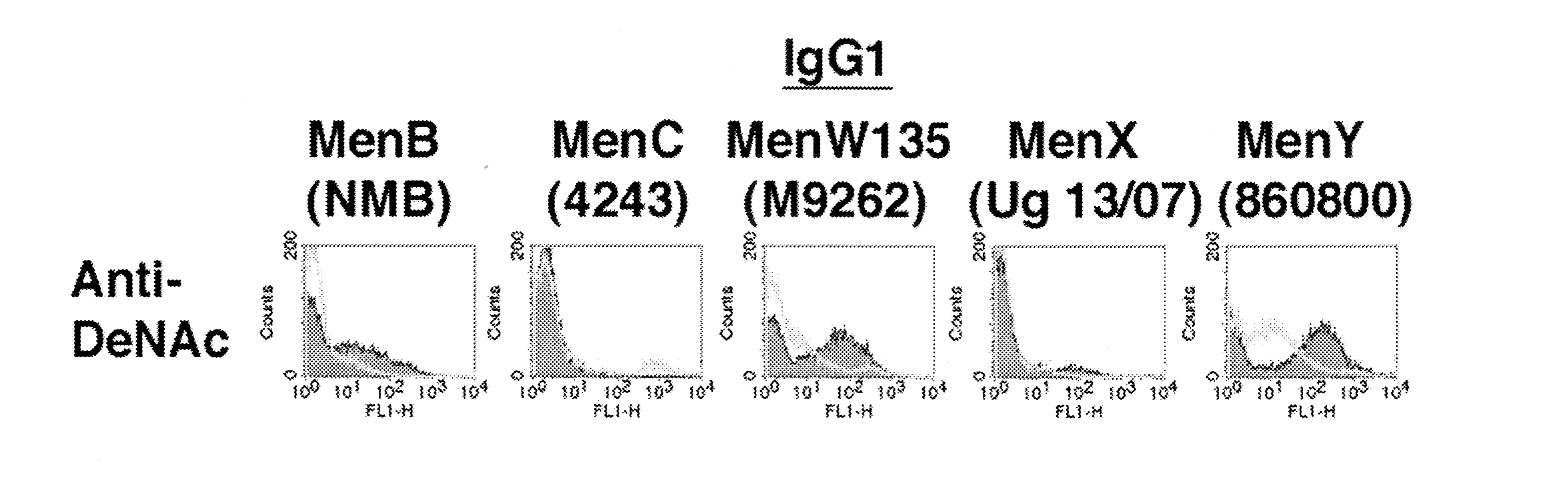
Recently, we observed that, with the exception of group C and X strains, the IgG component of the immune response to the DeNAc vaccine showing the greatest binding to meningococcal strains from multiple capsular groups was highly skewed to subclass IgG1 antibodies. For example, IgG1 antibody from the DeNAc antisera binding to MenB, C, W135, X, and Y strains compared to the carrier tetanus toxoid only antisera is shown in Figure 1.
Since mouse IgG1 is a poor activator of complement, it is possible that the screening strategy limited the ability to identify IgG antibodies reactive with Neu-containing antigens. Interestingly, antibodies elicited by the DeNAc vaccine that were reactive with MenC strains were mainly IgG2a, IgG2b, and IgM (data not shown), which may explain why the DeNAc antisera mediated bactericidal activity against C strains (1). As shown below, anti-Neu antigen mAbs do bind to and mediate SBA against X strains when the bacteria are cultured in media that contains a source of sialic acid. Again, the polyclonal anti-Neu antigen antibody responses that bind to group X strains grown in human serum are predominantly IgG1 (data not shown).
mAb binding to live N. meningitidis strains. We have observed recently that binding of several mAbs reactive with Neu- containing antigens (SEAM 2, SEAM 3 (6); DA1 and DA2) to meningococci can be affected by culture conditions such that rich media containing animal byproducts or human serum can result in a decrease or increase in binding, respectively (Flitter et al, in preparation and Figure 2). Therefore, we cultured the bacteria in CDM so that only endogenously produced Neu-containing antigens could be expressed.
As shown in Figure 2, both mAbs bind to MenB, C, W135 and Y strains but not the MenX strain when the bacteria are cultured in CDM. The capsular operons of MenB, C, W135 and Y strains contain genes encoding all of the enzymes necessary to produce Neu5Ac and, therefore, sialylated glycans (7).
In contrast, the MenX strain cannot produce Neu5Ac endogenously. However, when the MenX stain was cultured in CDM that was supplemented with 5% human plasma, both mAbs showed binding to the MenX strain. Supplementing CDM with CMP-Neu5Ac (8) did not result in reactivity with the MenX strain grown in CDM (data not shown). Interestingly, antibody binding decreased to all the strains grown in CDM supplemented with human serum including the positive control anticapsular mAbs.
]]> Since DA1 and DA2 are reactive with Neu-containing glycans but not the corresponding Neu5Ac derivatives (G. R. Moe, unpublished), the result suggests that N. meningitidis strains may express a sialic acid de-N-acetylase and that sialic acid antigens expressed on the bacterial surface can be acquired by strains that do not produce endogenous sialic acid.Currently, the only known mechanisms for the production of sialyl glycans in N. meningitidis is by de novo biosynthesis (reviewed in (9)) or sialylation of lipopolylsaccharide by exogenous host CMP-Neu5Ac (8). N. meningitidis do not express Neu5Ac transporters and are not known to have the ability to scavange sialic acid from host sialylglycans (9). Other
mechanisms such reverse sialylation of host sialylglycans (10) or specific lectins for binding host sialylglycans are possible but have not been demonstrated in N. meningitidis.
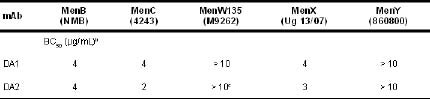
Serum bactericidal activity (SBA). The mAbs DA1 and DA2 were tested for the ability to mediate SBA against MenB, C, W135, X, and Y strains in the presence of human complement. As shown in Table 1, DA1 and 2 showed SBA against MenB, C, and X strains but not W135 or Y strains. One possible reason for the lack of activity against W135 and Y strains is that culturing the bacteria in Muller-Hinton media results in a decrease in DA1 and DA2 binding (data not shown).
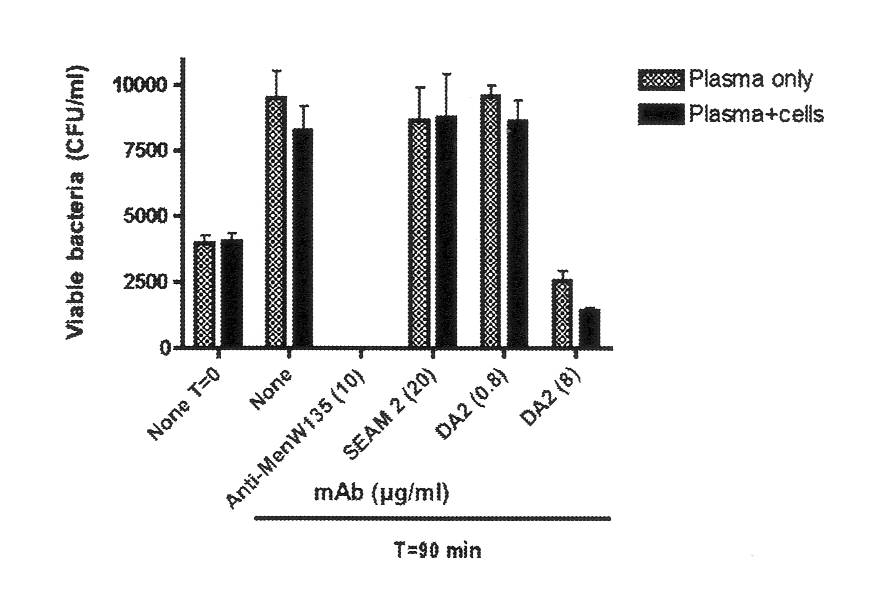
Although it has not yet been possible to repeat the SBA experiments for strains representing all capsular groups cultured in CDM or CDM supplemented with human serum, Figure 3 shows the results for DA2 against the same MenW135 strain (M9262) when the bacteria were cultured in CDM supplemented with 5% human plasma and the SBA assay was performed in human plasma without (65% plasma) or with leukocytes (45% plasma). As shown in Fig 3. DA2 was bactericidal against M9262 at a mAb concentration of 8 µg/mL but not 0.8 µg/mL and mediated increased cytotoxicity in the presence of phagocytic cells.
The positive control anticapsular mAb W3 was also protective but SEAM 2, which does bind to MenW135 strains cultured in CDM supplemented with human plasma (Flitter et al, in preparation) did not exhibit SBA. The results call attention to the importance of evaluating SBA of anti-Neu antigen antibodies under conditions that simulate bacteremia in humans as closely as possible. For reasons that are unclear, culturing meningococci in media containing animal byproducts has a detrimental effect on the expression of Neu-containing antigens.
Tabla 1. Serum bactericidal activity of DA1 and DA2 against N. meningitidis strains with human complementa.
]]>
aThe bacteria were cultured in Muller-Hinton media, which we now know affect the expression of Neu-containing antigens by meningococcal strains.
bSerum bactericidal titers were defined as the mAb concentration resulting in a 50% decrease in colony forming units (CFU) per mL after 60 minutes incubation of bacteria in the reaction mixture, compared to the control CFU per mL at time 0. Typically, bacteria incubated with the negative control antibody and complement showed a 150 to 300% increase in CFU/mL during the 60 minutes of incubation.
cSee Figure 3 for SBA results for DA2 when strain M9262 was cultured in CDM supplemented with 5% human serum.
Figure 1. Anti-DeNAc serum subclass IgG1 antibody binding to meningococcal strains from different capsular groups as determined by flow cytometry. Unfilled histograms are fluorescence of control anti-TT serum and filled histograms anti-DeNAc serum detected with FITC-conjugated anti-mouse IgG1 secondary antibody. The antisera were tested at a dilution of 1:10.
Figure 2. Flow cytometric analysis of binding of the mAbs DA1 and DA2 to meningococcal strains from different capsular groups. For each strain, there are two sets of data. The top row in each set is for bacteria cultured in CDM alone and the bottom row is for bacteria cultured in CDM supplemented with 5% human plasma as described in the Materials and Methods. Unfilled histograms are secondary antibody binding to the bacteria in the presence of an irrelevant IgM mAb (10 µg/mL). Filled histograms show fluorescence of bacteria in the presence the indicated mAb (1 µg/mL).
Figure 3. Effect of anti-Neu antigen mAbs on the viability of MenW135 strain M9262 in human plasma without or with leukocytes. Shaded bars indicate viability in plasma only in the presence of controls or test mAbs and filled bars in plasma with leukocytes at T=0 and T=90 minutes. The error bars represent the standard deviation of three replicate determinations from each assay. mAb concentrations are indicated in parentheses.
]]>
Conclusion
In summary, the data presented here shows that N. meningitidis strains express Neu-containing antigens or possibly antigens that are cross-reactive with anti-polyNeu antibodies under conditions that simulate growth in human blood. The latter possibility appears unlikely since the mAbs were not reactive with a group X strain that lacks the capability for de novo sialic acid biosynthesis when cultured in CDM but were positive for binding when CDM was supplemented with human plasma. The putative Neu-containing antigens are of considerable interest since all strains regardless of capsular group appear to express them, particularly when grown in the presence of human plasma. The expression of many antigens that are important for meningococcal pathogenesis are regulated by iron starvation (11), but thus far we have not observed any effect of removing iron on bacterial binding of several mAbs that are reactive with Neu-containing glycans (Flitter et al, in preparation). Currently, little is known about the expression of Neu-containing glycans in any organism and the role of such antigens in the pathogenicity of meningococci remains to be determined. However, the results presented here and elsewhere (1) clearly show that Neu-containing vaccines are immunogenic and elicit antibodies that bind to and mediate protective responses against MenB, C, W135, and X strains.
Acknowledgements
This work was supported by grant RO1 AI64314 from the National Institute of Allergy and Infectious Disease of the National Institutes of Health, Wellstat Vaccines, Inc., and the family of Jennifer Leigh Wells. The research described here was conducted in a facility constructed with support from Research Facilities Improvement Program Grant Number CO6 RR-16226 from the National Center for Research Resources, National Institutes of Health. G.R.M. has pending patent applications on the vaccines described in this report, Wellstat Vaccines, Inc. has provided research support and the technology has been licensed for development from Children's Hospital & Research Center at Oakland. G.R.M. could potentially receive royalty benefits.
References
]]>1. Moe GR, Bhandari TS, and Flitter BA. Vaccines containing de-N-acetyl sialic acid elicit antibodies protective against Neisseria meningitidis group B and C. J. Immunol. 2009. [in press].
2. Moe GR, Zuno-Mitchell P, Lee SS, Lucas AH, and Granoff DM. Functional activity of anti-Neisserial surface protein A monoclonal antibodies against strains of Neisseria meningitidis serogroup B. Infect Immun 2001;69:3762-71.
3. Cobb BA, and Kasper DL. 2005. Zwitterionic capsular polysaccharides: the new MHCII-dependent antigens. Cell Microbiol 7:1398-1403.
4. Cobb BA, Wang Q, Tzianabos AO, and Kasper DL. Polysaccharide processing and presentation by the MHCII pathway. Cell 2004;117:677-87.
5. Courtney AH, Puffer EB, Pontrello JK, Yang ZQ, and Kiessling L.L. Sialylated multivalent antigens engage CD22 in trans and inhibit B cell activation. Proc Natl Acad Sci U S A 2009;106:2500-05.
6. Granoff DM, Bartoloni A, Ricci S, Gallo E, Rosa D, Ravenscroft, et al. Bactericidal monoclonal antibodies that define unique meningococcal B polysaccharide epitopes that do not cross-react with human polysialic acid. J Immunol 1998;160:5028-36.
7. Swartley JS, Ahn JH, Liu LJ, Kahler CM, and Stephens CM. Expression of sialic acid and polysialic acid in serogroup B Neisseria meningitidis: divergent transcription of biosynthesis and transport operons through a common promoter region. J Bacteriol 1996;178:4052-59.
8. Parsons NJ, Andrade JR, Patel PV, Cole JA, and Smith H. Sialylation of lipopolysaccharide and loss of absorption of bactericidal antibody during conversion of gonococci to serum resistance by cytidine 5'-monophospho-N-acetyl neuraminic acid. Microb Pathog 1989;7:63-72.
9. Severi E, Hood DW, and Thomas GH. Sialic acid utilization by bacterial pathogens. Microbiology 2007;153:2817-22.
10. Chandrasekaran EV, Xue J, Xia J, Locke RD, Matta KL, and Neelamegham S. Reversible sialylation: synthesis of cytidine 5'-monophospho-N-acetylneuraminic acid from cytidine 5'-monophosphate with alpha2,3-sialyl O-glycan-, glycolipid-, and macromolecule-based donors yields diverse sialylated products. Biochemistry 2008;47:320-30.
11. Perkins-Balding D, Ratliff-Griffin M, and Stojiljkovic I. Iron transport systems in Neisseria meningitidis. Microbiol Mol Biol Rev 2004;68:154-171.
{kind=link}
{kind=link}
{kind=link}
{kind=link}