REPORT
Production of highly polymerized bacterial levan in two eukaryotic hosts of biotechnological interest
Producción de levana bacteriana altamente polimerizada en dos hospederos eucariontes de interés biotecnológico
]]> Alexander Banguela1, Luis E Trujillo1, Juan G Arrieta, Raisa Rodríguez1, Enrique Pérez2, Carmen Menéndez1, Ricardo Ramírez1, Merardo Pujol1, Carlos Borroto1, Lázaro Hernández1
1División de Plantas, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, PO Box 6162, Playa, CP 10600, La Habana, Cuba.
2Centro de Ingeniería Genética y Biotecnología de Sancti Spíritus, Cuba.
ABSTRACT
Bacterial levan [β(2,6)–linked polyfructan] has potential applications in the food, bio-energetic, medical, pharmaceutical, and other industries. The lack of technically and economically feasible large-scale production systems limits the commercial exploitation of levan. Gluconacetobacter diazotrophicus secretes a levansucrase (LsdA, EC 2.4.1.10) that synthesizes high levels of levan and fructooligosaccharides from sucrose. This bacterium is not attractive for the cost-effective production of LsdA. In this research, we used Pichia pastoris and Nicotiana tabacum as hosts for LsdA production and direct levan synthesis, respectively. The recombinant yeast constitutively expressing the lsdA gene acquired saccharolytic capacity and secreted LsdA to a yield 9-fold higher than the value reported for the natural host. The occurrence of N-glycosylation in the yeast-produced LsdA did not affect the catalytic efficiency, substrate specificity, or product profile compared to the native non-glycosylated enzyme. This finding prompted us to express the lsdA gene in vacuoles, the most physiologically appropriate compartment to direct levan formation within the plant cell. Constitutive expression of LsdA fused to the vacuolar targeting signal of an onion fructosyltransferase allowed the accumulation of highly polymerized levan (above 104 fructose residues) in mature tobacco leaves where the polymer represented between 10 and 70% (w/w) of total dry weight. The latter value is the highest reported in the literature for a levan-producing transgenic plant grown in soil. No drastic physiological changes were observed in tobacco plants with levan yields up to 30% (w/w) in leaves. The polymer production remained stable in the plant progenies pointing for potential application in biotechnology.
Keywords: Levan, transgenic tobacco, Pichia pastoris, levansucrase, N-glycosilation, catalytical properties, Gluconacetobacter diazotrophicus, fructan.
Levana bacteriana es un homopolímero β (2,6) soluble de unidades de fructosa, con aplicación en las industrias alimentaria, médico-farmacéutica, bioenergética y otras. La carencia de un sistema productivo factible, técnica y económicamente, impide la explotación comercial de levana a gran escala. La bacteria Gluconacetobacter diazotrophicus secreta una levanasacarasa (LsdA, EC 2.4.1.10) que sintetiza altos niveles de levana y fructooligosacaridos a partir de la sacarosa. Por ser el hospedero nativo poco atractivo para la producción masiva de LsdA, se investigaron la levadura Pichia pastoris y la planta Nicotiana tabacum como nuevas fuentes. La levadura P. pastoris recombinante expresó constitutivamente LsdA y adquirió actividad sacarolítica, e incrementó el rendimiento de LsdA en 8 a 9 veces con respecto a G. diazotrophicus. La N-glicosilación de LsdA en un hospedero eucarionte no afectó la eficiencia catalítica ni el rendimiento de fructanos en comparación con la enzima nativa. Ello permite la obtención de LsdA glicosilada activa en las vacuolas de las plantas, compartimento celular más apropiado para esos fines. La expresión constitutiva de LsdA fusionada con la señal de localización vacuolar de la fructosiltransferasa de cebolla permitió la acumulación de levana altamente polimerizada (más de 104 residuos de fructosa) en hojas de tabaco, entre 10 y 70% del peso seco total. Este último es el máximo valor descrito para una planta transgénica crecida en suelo, productora de levana. La ausencia de alteraciones fisiológicas drásticas en clones que acumularon hasta 30% de levana, y la producción estable del polisacárido en sus progenies, remarcan su potencial agronómico.
]]> Palabras clave: Levana, tabaco transgénico, Pichia pastoris, levanasacarasa, N-glycosilación, propiedades catalíticas, Gluconacetobacter diazotrophicus, fructano.
INTRODUCTION
Bacterial levan is a sucrose-derived polysaccharide consisting of above 10 000 fructose residues connected predominantly by β(2,6) linkages. This polyfructan finds applications in the food, medical, pharmaceutical, bioenergetic, and other industries [1]. The acid or exo-type enzymatic hydrolysis of levan results in highly pure fructose syrups of high demand as sweeteners, while the intact polymer is a low-calorie fiber with prebiotic effect demonstrated in humans. Levan as a fructose source is also a preferred feedstock for the production of dimethylfuran for liquid fuels. This polyfructan is attractive as a blood plasma volume extender and it was recently reported to have antitumor and antiviral effect [2]. Due to its high degree of polymerization and good solubility in water, bacterial levan is appropriate for industrial use as an emulsifying and encapsulant agent in glues, textile coatings, cosmetics and detergents. Currently, there is not a commercial technology for levan production by either natural or recombinant way.
The endophytic bacterium Gluconacetobacter diazotrophicus secretes a constitutively-expressed levansucrase (LsdA, EC 2.4.1.10) that converts sucrose into fructooligosaccharides (FOS) and levan [3]. The low expression levels of the lsdA gene in G. diazotrophicus, the limited cell density in bioreactors, and the technological constraints derived from the polysaccharides causing high dense culture supernatants, make the native bacterium inadequate for the cost-effective production of LsdA. As alternative approaches, we have studied here the expression of the lsdA gene in two model eukaryotic hosts of biotechnological interest: the non-saccharolytic yeast Pichia pastoris and tobacco (Nicotiana tabacum), a plant that does not synthesize fructans naturally.
The main objectives of this research were: 1) production and secretion of N-glycosylated LsdA in P. pastoris and evaluation of the enzyme yield and catalytic performance; 2) direct synthesis of levan by recombinant LsdA into cellular vacuoles of tobacco leaves and evaluation of its influence in plant physiology; 3) the development of useful genetic constructs, methodologies, and tools to use yeast and plants as bioreactors for the production of transgenic levan and other fructans of commercial interest.
The working strategy included the following steps: 1) expressing the lsdA gene in P. pastoris fused to a secretion signal peptide under the control of the methanol-inducible AOX1 promoter or the constitutive GAP promoter; 2) characterizing biochemically the LsdA secreted by the transgenic yeast; 3) comparing the recombinant glycosylated LsdA and the native unglycosylated enzyme in terms of catalytic efficiency, substrate specificity and product profile; 4) expressing the lsdA gene fused to the vacuolar localization signal peptide of an onion fructosyltransferase under the control of the constitutive CaMV35S promoter in transgenic tobacco plants; 5) quantifying the fructan levels in leaves, stems, roots, flowers and seeds of different transgenic lines; 6) isolating and determining the composition, size and structure of fructans from mature leaves; 7) elucidating the physiological influence of levan accumulation during the growth and development of transgenic plants; 8) determining the transgene stability and levan yield in T1 and T2 progenies.
]]> RESULTS AND DISCUSSION
Constitutive and methanol-inducible expression of the lsdA gene in P. pastoris
P. pastoris lacks endogenous saccharolytic activity; therefore, it is an ideal host to study heterologous expression of fructosyltransferases. In this sense, the lsdA gene fused to either the alpha-factor signal peptide or the acid phosphatase signal peptide, both from Saccharomyces cerevisiae, was expressed in P. pastoris under the control of either the methanol-inducible AOX1 promoter or the constitutive GAP promoter. The two expression systems allowed the secretion of active LsdA although efficiency differed [4-6]. In the induced (AOX1) system, 81% of the total levansucrase activity was localized into the periplasmic space and the rest in the culture medium. In the constitutive (pGAP) system, the recombinant enzyme was totally released to the culture medium.
The final LsdA yield of 100 mg/L achieved in the culture supernatant of one constitutive clone was 8-9 times higher that the value of 12 mg/L reported for the natural host G. diazotrophicus [7]. The enzyme productivity of the constitutive expression system (2.2 mg/h·L) duplicated that of the induced system (1.1 mg/h·L).
LsdA secreted in P. pastoris is a glycoprotein with unaltered catalytic performance
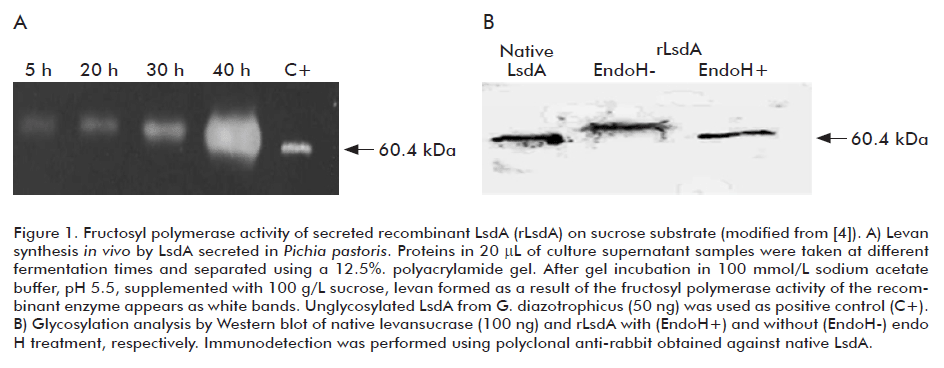
Figure 1A shows that LsdA secreted into the culture medium synthesized levan in vitro as a result of its fructosyl polymerase activity over the substrate sucrose. The migration of recombinant LsdA in SDS-PAGE corresponded to a protein of molecular weight above the theoretical mass of 58 kDa. Three potential N-glycosylation sites were identified on the mature LsdA sequence. Digestion with endoglycosidase H resulted in higher electrophoretic mobility of the recombinant enzyme in relation to the undigested control (Figure 1B), confirming the incorporation of N-linked oligosaccharides during the enzyme passage through the yeast secretory pathway [4]. Contrary to this result, the Bacillus subtilis levansucrase under its own signal peptide or fused to the S. cerevisiae acid phosphatase signal sequence was unable to enter the endoplasmic reticulum of the yeast and the enzyme accumulated inactive in its precursor form in the cytoplasmic membrane [8].
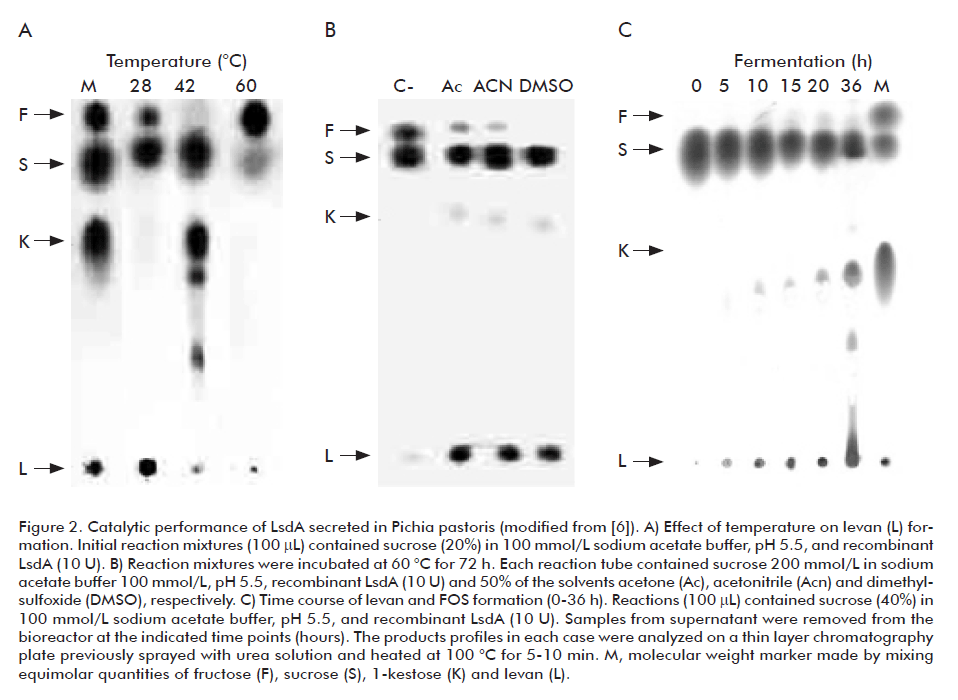
There are multiple reports of variations in the kinetic properties and substrate specificity of bacterial enzymes expressed in yeast due to the occurrence N-glycosylation, a posttranslational modification most commonly absent in prokaryotic proteins. For this reason, we studied in detail the biochemical properties of the glyco-LsdA produced in P. pastoris. The kinetic parameters Vmax (1.67 mM/min), KM (6.63 mM), kcat (57.1 x 103 min-1) and kcat/KM (8.6 x 103 mM-1 min-1) determined for the sucrose hydrolysis reaction of recombinant LsdA showed no significant differences with the corresponding values of the native enzyme [6]. Similarly, the N-linked mannose chains had no influence on the substrate specificity of the enzyme and its behavior towards pH and temperature. The levan yield (polymerization reaction) reached the maximal value at 28 °C (Figure 2A), while the release of free fructose from sucrose (hydrolysis reaction) became the predominant reaction when the incubation temperature was raised to 60 °C. By contrast, the reaction at 60 °C in a water-deficient environment (Figure 2B) favored the formation of levan with high polymerization degree [6]. Glycosylated LsdA showed a slight increase in thermostability [4]. This fact could be of relevance for the industrial operation of the recombinant enzyme.
The use of cane sugar (50-100 g/L) as carbon source to grow the P. pastoris clones constitutively expressing lsdA did not cause cell toxicity [5]. A time-course analysis of the carbohydrate composition in culture supernatant samples revealed the maximum fructan content of 59% (w/w) at the end of the fermentation (Figure 2C) when levan counted for 35% (w/w) of the total sugars. The glucose released from sucrose was consumed by the yeast during growth and it was hardly represented in the total sugar composition.
Constitutive expression of the lsdA gene and vacuolar localization of the enzyme in transgenic tobacco plants
Tobacco is a model plant widely used as a bioreactor for the production of foreign proteins, including enzymes of industrial interest. This is a naturally non-fructan crop with constant availability of sucrose in vacuoles, the cell compartment recommended for the production and accumulation of bacterial levan In vacuoles, the synthesized polyfructan would remain sequestered, minimizing its potential disruptive effects [9]. LsdA is expected to become a glycoprotein during transit to the vacuole in the tobacco cells. Since N-glycosylation did not alter the catalytic properties of LsdA secreted by P. pastoris, this modification should not compromise the enzyme folding and activity in plants either.
]]> In this study, the lsdA gene was fused to the vacuolar targeting signal of the onion sucrose:sucrose 1-fructosyltransferase (1-SST) and expressed in tobacco plants under the control of CaMV35S constitutive promoter. Unlike the non-transformed tobacco (NT), the high-molecular fructan was detected in leaves of three transgenic clones (Figure 3A). This polyfructan was also present in extracts of roots, stems and flowers, but not in seeds (results not shown). Northern blot analysis revealed vast amounts of lsdA transcripts in tobacco leaves of the clone with higher levan yield (Figure 3B). The recombinant polymer extracted from leaves contains more than 104 fructose residues as determined by acid hydrolysis (Figure 3C). Incubation of clarified leaf extracts from transgenic plants with sucrose produced FOS (1-kestose) and levan (Figure 3D). This result indicates that the recombinant LsdA in the cell vacuoles maintained the same product profile of the native enzyme, despite the fact that FOS were not detected in the in-vivo tests assayed on different plant organs, including mature leaves [10].Levan accumulation is proportional to the leaf age and the occurrence of phenotypic alterations in the transgenic plant
As shown in Figure 4A, levan content increased in the leaves of the transgenic clones with plant growth. In older leaves, levan contents of 10 ± 1, 30 ± 1.5, and 70 ± 2.5 mg/g fresh weight (10, 30 and 70% dry weight) were reached for clones 2, 6 and 3, respectively. This latter value is the largest reported for fructan production in leaves of transgenic tobacco plants and it is only surpassed by the so called fructan beet, a transgenic sugar beet plant expressing the Helianthus tuberosus 1-SST that accumulated fructans up to 90% (w/w, dry weight) in the storage root [11]. The transgenic tobacco plants did not show any visible phenotypic alteration during growth in greenhouse for the first 30 days. From this point, older levan-accumulating leaves bleached gradually from the tip toward the base and became rigid to the touch due to excessive turgor (Figure 4). The changes in leaf texture and color intensified during the flowering process. The germination percent of seeds was 97, 0, and 84% for clones 2, 3, and 6, respectively. Levan production remained stable in the second generation of clones 2 and 6.
RELEVANCE AND PRACTICAL APPLICATION OF THE RESEARCH
Due to its high degree of polymerization, good solubility in water, demonstrated prebiotic effect, non-immunogenic, antitumor and antiviral capacity, bacterial levan has wide potential applications. However, the lack of an industrial production system technically and economically feasible limits the commercial exploitation of this polyfructan at a large-scale level. In this paper, P. pastoris and N. tabacum were transformed into novel sources for massive levansucrase production and direct levan synthesis from plant sucrose, respectively.
This research constitutes the first report of constitutive expression and secretion of a bacterial fructosyltransferase in yeast, as well as the production of transgenic levan in a plant expressing the G. diazotrophicus levansucrase gene (lsdA). We demonstrated that N-glycosylation produced by eukaryotic hosts does compromise LsdA folding and activity. Levan accumulation in tobacco leaves reached up to 70% (w/w) of dry weight, which is the maximum reported value reported in the literature for a transgenic plant grown in soil. The lack of aberrant phenotypes in the progenies (T1 and T2) of plants with levan content in mature leaves ranging 10-30% (w/w, dry weight) is of physiological relevance and it could be exploited for biotechnological purposes. Our results also demonstrated the feasibility of using a plant fructosyltransferase pre-propeptide sequence to direct a bacterial protein to vacuoles in transgenic plants. In conclusion, our work provides knowledge, methodologies, tools, and experimental results pointing in favor to the use of transgenic yeast and plants as alternative hosts for the cost-effective production of bacterial fructosyltransferases and fructans with commercial application.
REFERENCES
]]>1. Kang SA, Jang KH, Seo JW, Kim KH, Kim YH, Rairakhwada D, et al. Levan: applications and perspectives. In: Rehm BHA, editor. Microbial Production of Biopolymers and Polymer Precursors. Norfolk: Caister Academic Press; 2009. p. 145-63.
2. Esawy MA, Ahmed EF, Helmy WA, Mansour NM, El-Senousy WM, El-Safty MM. Production of levansucrase from novel honey Bacillus subtilis isolates capable of producing antiviral levans. Carbohyd Polym. 2011;86(2):823-30.
3. Támbara Y, Hormaza JV, Pérez C, León A, Arrieta J, Hernández L. Structural analysis and optimised production of fructo-oligosaccharides by levansucrase from Acetobacter diazotrophicus SRT4. Biotechnol Lett. 1999;21(2):117-21.
4. Trujillo LE, Arrieta JG, Dafhnis F, Garcia J, Valdes J, Tambara Y, et al. Fructo-oligosaccharides production by the Gluconacetobacter diazotrophicus levansucrase expressed in the methylotrophic yeast Pichia pastoris. Enzyme Microb Technol. 2001;28(2-3):139-44.
5. Trujillo LE, Banguela A, País J, Tambara Y, Arrieta JG, Sotolongo M, et al. Constitutive expression of enzymatically active Gluconacetobacter diazotrophicus levansucrase in the methylothrophic yeast Pichia pastoris. Afinidad. 2002;59(500):365-70.
6. Trujillo Toledo LE, Gómez Riera R, Banguela Castillo A, Soto Romero M, Arrieta Sosa JG, Hernández García L. Catalytical properties of N-glycosylated Gluconacetobacter diazotrophicus levansucrase produced in yeast. Electron J Biotechnol. 2004;7(2):115-23.
7. Hernandez L, Arrieta J, Menendez C, Vazquez R, Coego A, Suarez V, et al. Isolation and enzymic properties of levansucrase secreted by Acetobacter diazotrophicus SRT4, a bacterium associated with sugar cane. Biochem J. 1995;309(Pt 1):113-8.
8. Scotti PA, Praestegaard M, Chambert R, Petit-Glatron MF. The targeting of Bacillus subtilis levansucrase in yeast is correlated to both the hydrophobicity of the signal peptide and the net charge of the N-terminus mature part. Yeast. 1996; 12(10):953-63.
9. Banguela A, Hernández L. Fructans: from natural sources to transgenic plants. Biotecnol Apl. 2006;23(3):202-10.
10. Banguela A, Arrieta JG, Rodriguez R, Trujillo LE, Menendez C, Hernandez L. High levan accumulation in transgenic tobacco plants expressing the Gluconaceto-bacter diazotrophicus levansucrase gene. J Biotechnol. 2011;154(1):93-8.
11. Sevenier R, Hall RD, van der Meer IM, Hakkert HJ, van Tunen AJ, Koops AJ. High level fructan accumulation in a transgenic sugar beet. Nat Biotechnol. 1998;16(9):843-6.
Luis E Trujillo. División de Plantas, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, PO Box 6162, Playa, CP 10600, La Habana, Cuba. E-mail: luis.trujillo@cigb.edu.cu.
]]>{kind=link}
{kind=link}
{kind=link}