
En el presente trabajo se realiza una revisión del conocimiento actual sobre la regulación de las concentraciones intracelulares del ion calcio, los principales mecanismos de entrada y salida de este a través de la membrana plasmática, con especial atención en el intercambiador Na+/Ca2+, y la función de este importante segundo mensajero en la secreción de insulina, así como la muerte celular programada de las células ß pancreáticas.
Palabras clave: Calcio, intercambio Na+/Ca2+, apoptosis, insulina.
El ion calcio es utilizado por las células para regular múltiples actividades. Al inicio de la vida, un incremento rápido de Ca+2 intracelular durante la fertilización desencadena el desarrollo de un nuevo organismo.1
Una vez diferenciadas las células, el Ca2+ es utilizado como una señal intracelular para el control de diferentes procesos, entre los cuales se encuentran la contracción muscular, la secreción, el metabolismo, la excitabilidad y la proliferación celular. El Ca2+ no puede ser degradado como ocurre con otras moléculas que actúan como segundos mensajeros, por lo tanto, las células regulan estrictamente las concentraciones intracelulares de este ion. En el agua de mar, donde se originó la vida, la concentración del otro catión divalente más común, el Mg2+, es mayor que la del Ca2+, sin embargo, este no es excluido del citoplasma en la magnitud que ocurre con el Ca2+. Una explicación para esta singularidad es la capacidad que posee el Ca2+, en mayor grado que el Mg2+, de precipitar al fosfato, el cual es empleado por las células para almacenar y utilizar la energía. Las principales fuentes de Ca2+ son el medio extracelular y algunos organelos intracelulares: el retículo endoplasmático (RE) o el retículo sarcoplasmático (RS) en las células musculares.1
Se plantea que el mecanismo general de acción del Ca2+ es el siguiente: cuando las concentraciones del Ca2+ citoplasmático se mantienen en niveles bajos (10-00 nmol/L), las células permanecen quiescentes; sin embargo, cuando esas concentraciones se elevan (500-1 000 nmol/L) dichas células se activan para realizar sus funciones específicas. Existen sensores intracelulares responsables de la detección de los aumentos de Ca2+, como son la calmodulina (CAM) y la troponina C (TnC). Esta señal es traducida en respuestas específicas de las células.1
La activación de las células está determinada por un balance entre mecanismos Ca2+-ON y Ca2+-OFF (figura 1). Como se detalla a continuación, todos los elementos que promueven la entrada de Ca2+ hacia el citosol forman parte de los mecanismos ON, y los que realizan la función contraria se incluyen en los mecanismos OFF.
Las células presentan dos fuentes principales de Ca2+: la extracelular y la existente en organelos intracelulares. El reservorio más importante de Ca2+ intracelular es el retículo endoplasmático, el cual es denominado retículo sarcoplasmático en las células musculares. Por ser la concentración de Ca2+ en el exterior de la célula ~20,000 veces mayor que en el citoplasma y que el interior de la célula, y está cargado negativamente con respecto al exterior, existe un gradiente electroquímico que favorece la entrada de Ca2+. Los 2 grupos de canales encargados de la liberación de Ca2+ hacia el citosol son: los canales de Ca2+ de la membrana plasmática1 y los canales de Ca2+ intracelulares.1,2 El primer grupo se compone de los canales sensibles a voltaje (VOCs), los canales regulados por ligando (ROCs) y los canales activados por liberación de Ca2+ (CRAC). En el segundo grupo se incluyen los canales intracelulares: los receptores de inositol 1,4,5-trifosfato2 y los receptores de rianodina (RyRs).2 Además, el Ca2+ puede salir del retículo endoplasmático a través de un mecanismo denominado "fuga de Ca2+".3
En las células no excitables, como son las sanguíneas, los hepatocitos y las endoteliales, el incremento de las concentraciones de Ca2+ se realiza predominantemente a través de la vía del inositol 3-fosfato (InsP3). Existen dos clases de receptores que conducen a la liberación de InsP3 por diferentes vías: los receptores que se acoplan a la proteína G (GCR) y los receptores con función de tirosino-quinasas (RTK). Los receptores GCR activan a la fosfolipasa Cß mientras que los RTK realizan la misma función sobre la fosfolipasa Cg. Ambas fosfolipasas catalizan la conversión del fosfatidilinositol (4,5) difosfato en InsP3 y diacilglicerol.4 El InsP3 actúa como un segundo mensajero mediante su unión al receptor tetramérico del InsP3 (~ 310 kD a cada subunidad) situado en la membrana del retículo endoplasmático y este produce la liberación de Ca2+ almacenado en este organelo.1 La liberación de Ca2+ por esta vía puede incrementar la concentración de Ca2+ citosólico desde ~0,1 hasta ~1 µmol/L. El receptor de InsP3 se encuentra expresado, en mayor o en menor medida, en todos los tipos celulares. La unión del InsP3 a cada subunidad del receptor ocurre en la región N-terminal cargada positivamente porque esta es rica en arginina y lisina. Esta unión puede ser bloqueada por la heparina y no se conoce otro antagonista efectivo hasta el momento. El receptor de InsP3 posee una homología parcial con el receptor de rianodina, mientras que esta semejanza no es significativa en relación con los canales de Ca2+ sensibles a voltaje. La región N-terminal de este receptor contiene dos sitios de unión a ATP y un sitio de unión al Ca2+ 5. La regulación de la actividad del receptor de InsP3 es compleja, pues este se desensibiliza por el propio InsP3, es sensible a las concentraciones de Ca2+ citoplasmático y puede ser fosforilado por la protein-quinasa A.6 La sensibilidad de este receptor es mayor a concentraciones de InsP3 entre 0,5 y 1 µmol/L; además, no es sensible al InsP3 a muy altas o muy bajas concentraciones de Ca2+ citoplasmático.6,7
Las células excitables, como son las nerviosas y musculares, además de poseer el sistema descrito anteriormente para las no-excitables, poseen canales de Ca2+ sensibles a voltaje. Estos canales de Ca2+ producen incrementos superiores de la concentración citosólica de Ca2+ en comparación con los receptores del retículo endoplasmático. Esto permite que las proteínas cercanas a la superficie interna de la membrana plasmática, las cuales constituyen sensores de las variaciones de la concentración de Ca2+, inicien funciones especializadas como son la exocitosis de neurotransmisores en las neuronas y la contracción en el músculo. La despolarización a partir del potencial basal de membrana (entre -90 y -70 mV, generalmente) inicia cambios conformacionales en los canales de Ca2+ dependientes de voltaje, específicamente en regiones especiales sensibles a voltaje denominadas S4.1 Estos cambios conformacionales permiten el flujo de Ca2+ a través de la membrana plasmática. Para el Ca2+, el balance entre las fuerzas químicas y eléctricas es equivalente a la fuerza eléctrica resultante de un voltaje aproximado de +150 mV, por lo tanto, existe un gradiente favorable para la entrada de este ion en todos los potenciales de membrana fisiológicos (desde -90 hasta +60 mV). La actividad de los canales de Ca2+ sensibles a voltaje está autolimitada, ya que estas proteínas se inactivan rápidamente después de la despolarización. Por otra parte, una despolarización de mayor magnitud sólo provocaría la disminución de la fuerza electroquímica que permite la entrada de Ca2+ al citoplasma. En las células excitables, la entrada de Ca2+ a través de los canales de Ca2+ sensibles a voltaje produce directamente la activación de los receptores de rianodina, los cuales provocan la liberación del Ca2+ acumulado en el retículo endoplasmático.1 El receptor de rianodina es tetramérico y está formado por subunidades de ~560 kDa de peso molecular relativo. Su función es modulada por Ca2+, Mg2+ y ATP; sin embargo, la inhibición ejercida por los primeros iones ocurre en el rango de los mmol/L de concentración. Relativamente, el receptor de rianodina no es selectivo para los cationes aunque sí excluye a todos los aniones.1
A principio de la década del 90, fue descrito que el vaciamiento de los reservorios intracelulares de Ca2+ inducía la entrada de este ion a través de la membrana plasmática por un mecanismo desconocido hasta ese momento.8 Este fenómeno se denominó entrada capacitiva de Ca2+. Por ejemplo, la estimulación de los receptores muscarínicos tipo 3, los cuales incrementan las concentraciones de InsP3 a través de la activación de la fosfolipasa Cß, producen una elevación rápida y durante un período corto de las concentraciones de Ca2+ en el citosol como consecuencia de la salida de Ca2+ del retículo endoplasmático.9 Sin embargo, las altas concentraciones de Ca2+ citosólico se mantienen por un tiempo prolongado mediante mecanismos de entrada capacitiva de Ca2+. El potencial de membrana puede influir en este proceso, pues la hiperpolarización puede incrementar la concentración de Ca2+ en el citoplasma. Se ha propuesto que el papel fisiológico de la entrada capacitiva de Ca2+ es permitir el retorno del Ca2+ a los reservorios intracelulares de una forma más rápida. La entrada de Ca2+ a través de la membrana plasmática en ausencia de despolarización ha sido descrita por varios autores10-12 y esta pudiera ocurrir a través de los denominados canales activados por liberación de Ca2+ (CRAC); sin embargo, ninguno de estos supuestos canales iónicos ha sido purificado o clonado. Por otra parte, la corriente neta de Ca2+ relacionada con el mecanismo de entrada capacitiva es ~ 5 pA en una célula entera, a diferencia de los cientos de pA de corriente neta que atraviesan la membrana plasmática a través de los canales de Ca2+ sensibles a voltaje. La entrada capacitiva de Ca2+ es, además, inactivada por altas concentraciones de Ca2+ en el citoplasma.
Aunque el incremento de Ca2+ citosólico producido por la salida de Ca2+ del retículo endoplasmático ocurre fundamentalmente a través de los receptores de InsP3 y rianodina, se ha descrito la existencia de un fuga basal de Ca2+ el cual es independiente de estos mecanismos.3 Por ejemplo, la inhibición de la Ca2+-ATPasa del retículo endoplasmático como consecuencia de la exposición a la tapsigargina o la eliminación de ATP, da lugar a la liberación relativamente rápida del 92 % del Ca2+ secuestrado en este organelo en células ß de ratón.13
Aunque la principal función del intercambiador Na+/Ca2+ es el transporte de Ca2+ desde el citoplasma hacia el medio extracelular, esta proteína también contribuye a la entrada de Ca2+ mediante un fenómeno denominado intercambio Na+/Ca2+ en modo reverso. Como se describe posteriormente, el intercambio Na+/Ca2+ en el modo reverso puede ocurrir en determinadas condiciones fisiológicas, por ejemplo, durante la despolarización de la membrana o el incremento de la concentración intracelular del ion sodio.
En la célula ß pancreática, los mecanismos de entrada de Ca2+ hacia el citoplasma son similares a los presentes en las células excitables, aunque los primeros muestran algunas particularidades importantes. Por ejemplo, la exposición a la cafeína (10 mmol/L) o a la rianodina (1 µmol/L) no produce la liberación del Ca2+ almacenado en el retículo endoplasmático de las células ß, y esto ocurre por la ausencia de receptores de rianodina;13 sin embargo, estas células sí poseen canales de Ca2+ sensibles a voltaje en la membrana plasmática, característica típica de las células excitables. Por consiguiente, la entrada de Ca2+ a través de estos canales, inducida indirectamente por las altas concentraciones de glucosa extracelular y el incremento del contenido de ATP intracelular, lejos de provocar un aumento adicional de la concentración de Ca2+ citosólico por el vaciamiento del retículo endoplasmático, facilita la entrada de Ca2+ a este organelo a través de la Ca2+-ATPasa localizada en su membrana. Por otra parte, y a diferencia de las células musculares, la membrana del retículo endoplasmático de las células ß exhibe altos niveles de fuga pasiva de Ca2+, y este fenómeno puede ser observado cuando las células se exponen a inhibidores de la Ca2+-ATPasa del retículo endoplasmático o se les limita la disponibilidad de ATP.13 Estas peculiaridades de la célula ß en cuanto al manejo del Ca2+ se ajustan estrechamente a su función principal, la producción y secreción de insulina. En este sentido, un grupo de investigadores reveló la importancia de la presencia de un alto contenido de Ca2+ en el retículo endoplasmático para el correcto procesamiento de la proinsulina y la transferencia de este precursor a los gránulos secretores.13
Las células poseen una gran variedad de proteínas que unen Ca2+, y estas contribuyen a las señales mediadas por Ca2+, ya sea cuando actúan como tampones del Ca2+ libre, que modulan la respuesta celular, o cuando lo hacen como sensores que median el papel del Ca2+ como mensajero intracelular (figura 1).14 Las proteínas capaces de funcionar como tampones de Ca2+ se encuentran tanto en el citoplasma como en el interior del RE. Sólo una pequeña proporción de moléculas citoplasmáticas capaces de unir Ca2+ actúan como sensores, el resto parece funcionar exclusivamente como tampones. El Ca2+ unido a estos tampones se encuentra en equilibrio con el Ca2+ citoplasmático libre y como los mecanismos de extrusión disminuyen la concentración del Ca2+ citoplasmático, la cantidad de Ca2+ unido a estos tampones también disminuye.5
]]> La calmodulina es el principal sensor de Ca2+ en las células ß.16 Esta proteína de 17 kDa de peso molecular relativo es un receptor intracelular ubicuo de Ca2+ el cual contiene, como el elemento estructural que une Ca2+ de manera específica, a la denominada mano-EF.17 El término mano-EF describe a dos secuencias de estructura helicoidal las cuales se orientan perpendicularmente entre ellas y están conectadas por un lazo que contiene residuos aminoacídicos con alta afinidad por el Ca2+.Las proteínas que contienen este elemento estructural son denominadas proteínas de mano-EF.18 La calmodulina contiene 4 elementos del tipo mano-EF y todos pueden unir Ca2+ con una afinidad entre 10-5 y 10-6 mol/L con alguna cooperatividad y una leve dependencia de las concentraciones de Mg2+. Después de la unión del Ca2+, la calmodulina es capaz de interactuar con algunas proteínas dentro de las que se destacan las protein-quinasas dependientes de calmodulina/Ca2+. Las regiones de interacción con la calmodulina/Ca2+ de estas proteínas no constituyen secuencias conservadas, aunque generalmente son estructuras helicoidales a con un alto contenido de aminoácidos básicos.
La calbindina-D28 es otra proteína de la superfamilia de proteínas de mano-EF. Esta posee 6 elementos estructurales del tipo mano-EF pero el sexto elemento es aberrante y no une Ca2+. Se sugiere que la calbindina-D28 cumpla la función de tampón de Ca2+ citoplasmático en la célula ß, pues no se han encontrado enzimas que interactúen con ella.19
Desde el descubrimiento de la calmodulina como la principal proteína que une Ca2+ en las células ß pancreáticas16 se sugirió que las protein-quinasas dependientes de Ca2+/calmodulina (quinasas CaM) eran las principales mediadoras de la acción del Ca2+ como segundo mensajero. Seguidamente, se observó actividad de la quinasa de la cadena ligera de la miosina (MLCK) en preparaciones de células de islotes pancreáticos,20 lo cual indicaba que la fosforilación de la miosina ejercía una influencia en la secreción de insulina a través de la modulación de la interacción actina-miosina;20,21 sin embargo, recientemente se describió que la acción de la MLCK se limita a promover la movilidad de los gránulos secretores proximales.22 Contemporáneamente, se encontró que las células ß poseían otra enzima con actividad de quinasa dependiente de Ca2+/calmodulina, la cual fosforilaba proteínas diversas.23,24 Se determinó que esta proteína era la denominada quinasa CaM II y su activación provocada por secretagogos de insulina se correlacionaba con la secreción de esta hormona.25 A pesar de que la quinasa CaM II es una enzima que puede fosforilar varias proteínas, su función puede ser limitada por su localización intracelular. En la célula ß, la quinasa CaM II se localiza en los gránulos secretores,26 lo cual indica la participación específica de esta enzima en el proceso de secreción de insulina. Para conocer la relevancia de la activación de la quinasa CaM II en la secreción de insulina, se identificaron las proteínas que son fosforiladas por esta enzima, además de los efectos que las modificaciones de estas pudieran tener en su función. La sinapsina I y la MAP-2 son los únicos substratos de la quinasa CaM II validados hasta la actualidad en la célula ß27,28 y estas realizan funciones similares en la secreción neuronal.29,30 La última etapa de la exocitosis de la insulina comienza con la interacción de proteínas que se encuentran en las vesículas secretoras (v-SNARE) y la membrana plasmática (t-SNARE).31 Este complejo de anclaje, en realidad, es un receptor que reconoce a la NSF (proteína de fusión sensible a la N-etil-maleimida) y a la SNAP (proteína de adherencia a la NSF soluble). La NSF posee la capacidad de hidrolizar ATP y suministra la energía requerida en la formación de vesículas competentes (denominadas vesículas "iniciadas") para la fusión con la membrana plasmática.32 La fusión de los gránulos secretores a la membrana plasmática es independiente de protein-quinasas y se piensa que este proceso ocurra por la acción directa del Ca2+ a través de proteínas que unen complejos Ca2+/fosfolípidos, como son la sinaptotagmina 33 y la sincollina.34 Para la fusión de los gránulos secretores a la membrana plasmática, las concentraciones de Ca2+ deben estar en el orden de los µmol/L y es muy probable que estas se alcancen en esta zona, pues los canales de Ca2+ tipo-L se concentran en las regiones de la membrana plasmática donde la exocitosis es más activa.35 Sólo una mínima fracción (~1 %) de los gránulos secretores se encuentran en el estado de vesículas "iniciadas"36,37, o sea, listos para fusionarse con la membrana plasmática; sin embargo, esta pequeña proporción de vesículas competentes puede contener la insulina suficiente para suplir la denominada primera fase de liberación de insulina.36 Por su parte, la secreción prolongada de la hormona depende del reclutamiento de gránulos de los reservorios adicionales o de reserva donde la quinasa CaM II desempeña un papel importante.
La fosforilación de la sinapsina I por la quinasa CaM II ha sido propuesta como el mecanismo clave en el reclutamiento de vesículas sinápticas, en células neuronales, para la liberación de su contenido hacia el medio extracelular. En la célula ß pancreática, la sinapsina I es fosforilada por la quinasa CaM II cuando las concentraciones de Ca2+ citoplasmático se elevan a niveles que se correlacionan con la secreción de insulina.28,38
El proceso de ensamblaje y el desensamblaje de los microtúbulos es un factor importante en la secreción celular. La estabilidad de estas estructuras es controlada por el estado de fosforilación de proteínas asociadas a los microtúbulos.39 La fosforilación de la proteína MAP-2 por la quinasa CaM II reduce su afinidad por la tubulina y favorece la ruptura del microtúbulo.30 La MAP-2 aparentemente desempeña un papel importante en la secreción de insulina, pues se determinó que los gránulos secretores purificados sólo interactúan con los microtúbulos en presencia de MAP-2 exógena.40 Además, la utilización del agente fosfato de estramustina, el cual interactúa selectivamente con el dominio de unión de la MAP-2 con los microtúbulos, es capaz de inhibir la secreción de insulina.41 La fosforilación de la MAP-2 también puede regular la secreción de insulina por su influencia sobre la interacción de los microtúbulos con los microfilamentos de actina.42,43 En la célula ß, los microfilamentos forman una red que actúa como una barrera que impide la migración de los gránulos secretores hacia la membrana plasmática, aunque también puede proveer la fuerza motriz de dicha migración a través de su contracción cuando la célula es estimulada.44 El reordenamiento de la red de actina cortical es dependiente de Ca2+ en células neuroendocrinas.45
La quinasa CaM II posee la capacidad de autofosforilarse. Esta característica permite que la enzima pueda fosforilar substratos durante un cierto tiempo posterior al cese del estímulo, lo cual es importante e incluso crítico para la función de la célula ß.41 Por ejemplo, la célula ß para responder apropiadamente a estímulos repetidos requiere de adecuadas reservas de gránulos secretores. Por lo tanto, la activación de la quinasa CaM II es importante no sólo para el sustento de la denominada segunda fase de liberación de insulina, sino también para el reaprovisionamiento de las reservas de gránulos después que la estimulación cesa. La quinasa CaM II es desfosforilada por la proteína-fosfatasa I, pero la inactivación total de esta ocurre alrededor de 5 minutos después que el estímulo finaliza.46
La calsecuestrina es el principal tampón de Ca2+ en el retículo sarcoplasmático de los músculos estriados, y su función principal es el almacenamiento de Ca2+ en este organelo.47 Esta proteína de 45 kDa de peso molecular relativo facilita el transporte de Ca2+ desde los sitios de entrada a través de la Ca2+ ATPasa del retículo sarcoplasmático hasta los sitios de liberación a través de los receptores de rianodina. La calsecuestrina, además de unir Ca2+ con una baja constante de disociación (400-600 µmol/L en presencia de KCl 150 mmol/L y 100 µmol/L en presencia de KCl 20 mmol/L) y una alta capacidad (40-50 mol/mol), posiblemente neutraliza los efectos potencialmente negativos de las altas concentraciones de Ca2+ sobre la función del retículo sarcoplasmático (ej. la inhibición de la Ca2+ ATPasa).48 El punto isoeléctrico de este tampón es pH 3,75 y posee más de un 30 % de residuos ácidos. La calsecuestrina se expresa también en el músculo liso y en otros tipos celulares como las neuronas e incluso en células de plantas.15
La calreticulina, cuyo peso molecular relativo es de 60 kDa, se considera la proteína homóloga de la calsecuestrina en células no musculares, y aunque está presente en el retículo endoplasmático a altas concentraciones, también se expresa en el núcleo celular. La calreticulina contiene 37 residuos ácidos en el dominio cercano al extremo carboxilo-terminal y une Ca2+ de dos modos: con alta capacidad (25 mol/mol) y baja afinidad (Kd ~ 250 µmol/L) y con baja capacidad (1 mol/mol) y alta afinidad (Kd ~ 1 µmol/L). El extremo carboxilo terminal culmina con la secuencia señal KDEL (lisina-ácido aspártico-ácido glutámico-leucina). Esta secuencia interactúa con el receptor KDEL, y por lo tanto, la calreticulina es retenida mayoritariamente en el retículo endoplasmático. Además de su función como tampón de Ca2+, la calreticulina realiza otras funciones, dentro de las cuales se destacan la regulación de la proliferación celular y la transcripción génica.49
Una vez que el Ca2+ citoplasmático alcanza el nivel umbral para generar señales de activación de la célula, se desencadenan los procesos que promueven el retorno al estado basal de la concentración de Ca2+. Estos procesos son muy activos, pues deben extrudir no sólo el Ca2+ libre sino también el Ca2+ unido a tampones y sensores cuando el catión se disocia de estos. La eliminación del Ca2+ citoplasmático es realizado por bombas e intercambiadores. ]]>
Las células disponen de 2 familias de Ca2+-ATPasas: la Ca2+-ATPasa de la membrana plasmática (PMCAs)50 y la Ca2+-ATPasa del retículo sarco/endoplasmático (SERCAs),15 cuyas funciones son transportar el Ca2+ introducido en el citoplasma hacia el medio extracelular o hacia el retículo endoplasmático, respectivamente. Ambas Ca2+-ATPasas utilizan el ATP como fuente de energía para transportar Ca2+ contra el gradiente electroquímico existente a través de las membranas donde estas se localizan.50 Ellas son miembros de las ATPasas tipo P, pues forman un intermediario covalentemente fosforilado durante el ciclo de reacción.15,50
La PMCA es una proteína de ~135 kDa de la cual se han descrito hasta la actualidad 4 isoformas. Cada isoforma de la PMCA puede mostrar variabilidad estructural, la cual es generada por splicings alternativos de los transcriptos primarios.51 Esta proteína parece estar expresada en todas las células eucarióticas y se considera que constituye el principal transportador de Ca2+ de alta afinidad localizado en la membrana plasmática.52 La PMCA posee 10 segmentos que atraviesan la membrana plasmática, mientras que los extremos carboxilo y amino se localizan en la parte citoplasmática de esta. La mayor parte de la PMCA se encuentra localizada hacia el citosol y consiste en 3 grandes fragmentos: un lazo intracelular limitado por los segmentos transmembrana 2 y 3, otro entre los segmentos 4 y 5 y finalmente una larga "cola" a continuación del último segmento transmembrana. El segundo lazo (constituido por ~400 residuos) contiene el dominio catalítico el cual incluye el sitio de unión del ATP y el residuo de aspartato que forma el intermediario acil fosfato durante la hidrólisis del ATP. La extensa "cola" carboxiterminal constituye el principal dominio regulatorio de la PMCA.53 El complejo Ca2+-calmodulina se une a una secuencia situada a una distancia de ~40 aminoácidos posterior al último segmento transmembrana. En ausencia de Ca2+-calmodulina esta secuencia actúa como un dominio autoinhibitorio, pues interactúa con dos sitios situados en el primer y segundo lazos citosólicos, mientras mantienen a la proteína en un estado inactivo. Cuando se incrementa la concentración de Ca2+ en el citosol, aumenta también la concentración del complejo Ca2+-calmodulina, el cual se une con alta afinidad al sitio autoinhibitorio. Consecuentemente, la PMCA se libera de la autoinhibición y su actividad se recupera. La Km para el Ca2+ de la PMCA es 0,4-0,7 µmol/L en presencia de Ca2+-calmodulina.54
La estructura de la SERCA se asemeja considerablemente a la estructura de la PMCA. La diferencia estructural principal entre ambas bombas está confinada a la "cola" carboxi-terminal. Este segmento es mucho más pequeño en la SERCA (~20-50 residuos) que en la PMCA (~70-200 residuos). El peso molecular relativo de la SERCA es de ~110 kDa y transporta 2 iones Ca por cada ATP hidrolizado, a diferencia de la PMCA, la cual transporta solo 155. Mediante estudios de mutagénesis se han identificado 2 sitios (I y II) de alta afinidad por el Ca2+ en la SERCA, formados en su mayor parte por residuos ácidos conservados en todas las bombas tipo P. Estos sitios se encuentran localizados dentro de los segmentos transmembrana 4, 5 y 6.56 La PMCA, sin embargo, conserva sólo el sitio II. La SERCA, además, contiene secuencias que interactúan con la tapsigargina, su inhibidor específico, el cual fue aislado originalmente de la planta Thapsia garganica.57
El intercambiador Na+/Ca2+ (NCX) se encuentra en la membrana plasmática y utiliza la energía almacenada en el gradiente electroquímico de Na+ para extrudir Ca2+ de la célula.58 Este mecanismo de extrusión está más desarrollado en células excitables como son las neuronas, el músculo cardíaco y liso, y en células neuroendocrinas, como las ß pancreáticas. El intercambiador Na+/Ca2+ puede, en ocasiones, operar en el modo reverso y causar influjo de Ca2+, como ocurre en el potencial de acción cardíaco, donde un incremento temporal de la concentración de Na+ en el citoplasma revierte la dirección de operación del intercambiador, que resulta en un rápido influjo de Ca2+. Una vez que la concentración de Na+ disminuye en el interior de la célula, el intercambiador revierte su acción a la extrusión de Ca2+ 59.
En la década de los años 60, se obtuvieron las primeras evidencias acerca de la existencia del intercambiador Na+/Ca2+ en el músculo cardíaco60,61 y desde entonces esta proteína ha sido detectada virtualmente en todos los tejidos62 y en especies, como el hombre,63 el perro,64 el calamar65 y la mosca de la fruta.66 La función principal del intercambiador Na+/Ca2+ es la de extrudir Ca2+ desde el citosol, intercambiándolo por Na+, sin consumo directo de ATP, pues utiliza el gradiente electroquímico favorable del último ion. Este intercambiador puede actuar en el modo reverso bajo algunas condiciones, permitiendo la entrada de Ca2+ al citosol. Además, el intercambiador Na+/Ca2+ puede realizar intercambios Ca2+/Ca2+ y Na+/Na+.67 Todas las formas del movimiento de iones del intercambiador se resumen en la figura 2.

FIG. 2. Diferentes modos de transporte de iones vía el intercambiador Na+/Ca2+.
]]>El intercambio Na+/Ca2+ es electrogénico, pues su estequiometría es 3Na+:1Ca2+. 68,69 Por lo tanto, se crea una corriente eléctrica hacia el citoplasma por el movimiento de una carga positiva neta. Además, el intercambiador Na+/Ca2+ es sensible a cambios en el potencial de membrana y al gradiente de Na+. Recientemente, determinaciones realizadas del potencial reverso de la corriente del intercambio Na+/Ca2+, han sugerido que la estequiometría de la proteína intercambiadora pudiera ser de 4Na+:1Ca2+ y que esta varía en dependencia de las concentraciones iónicas de Na+ y Ca2+ en el lado citoplasmático.70
El intercambio Na+/Ca2+ es una reacción que incluye 2 sustratos y su mecanismo puede ser clasificado como secuencial o consecutivo (ping-pong). En el mecanismo consecutivo, el primer sustrato es unido por la proteína y el primer producto es liberado antes de la unión del segundo sustrato, mientras que en el secuencial, ambos sustratos se unen a la proteína antes de la liberación de los productos. Mediante la determinación de las velocidades iniciales de incorporación de 45Ca2+ dependiente de Na+ a proteoliposomas reconstituidos con el intercambiador Na+/Ca2+, donde se incorporó el agente quelante EDTA y el estudio de los parámetros cinéticos, se propuso que el intercambio Na+/Ca2+ se realiza a través de un mecanismo consecutivo.71
De igual forma, la utilización de técnicas electrofisiológicas de alta resolución ha permitido resolver ciclos de reacciones parciales en el intercambio Na+/Ca2+ 72. Aparentemente, el paso electrogénico de este mecanismo de transporte iónico incluye la translocación del ion sodio, mientras que el movimiento del Ca2+ es electroneutral. Esto último implica que 2 cargas negativas pertenecientes a la proteína del intercambiador Na+/Ca2+ atraviesan el campo eléctrico creado a través de la membrana plasmática mediante cambios conformacionales que acompañan el transporte iónico.69,73,74 El ciclo de reacción hipotético para el intercambio Na+/Ca2+ se muestra en la figura 3.75,76
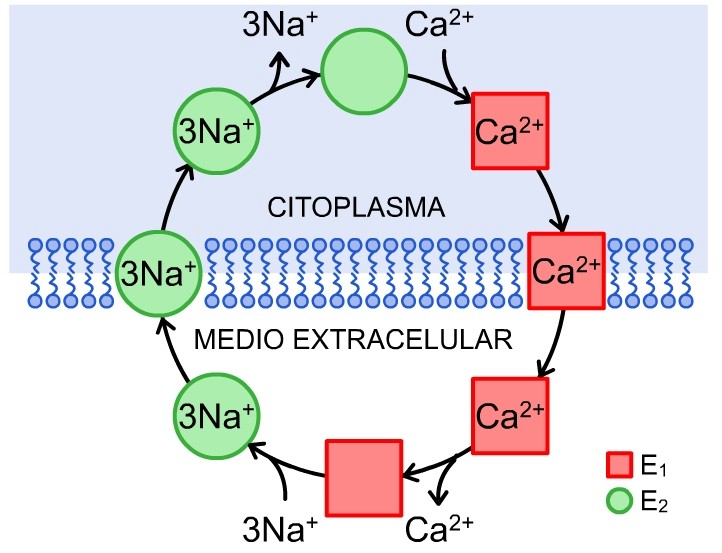
FIG. 3. Mecanismo de intercambio Na+/Ca2+ en el modo directo (extrusión de Ca2+). El intercambiador expone su sitio de unión a iones hacia el lado citoplasmático (conformación E2), une un Ca2+ y provoca así un cambio de conformación hacia E1. Posteriormente el Ca2+ es liberado en el lado extracelular, se unen 3Na+ y ocurre un cambio de conformación de regreso hacia E2. El sitio que une Na+ se expone hacia la cara citoplasmática, se liberan los iones de Na+, y el intercambiador vuelve a unir Ca2+ para completar el ciclo.
Clonaje del intercambiador Na+/Ca2+. Hasta el momento, 3 isoformas del intercambiador han sido clonadas en mamíferos: la NCX1,64 la NCX277 y la NCX3.78 La NCX1 se encuentra ampliamente distribuida en el corazón63 y en muchos otros tejidos y tipos celulares.62 En cambio, la expresión de la NCX2 y la NCX3 se limita al cerebro y al músculo esquelético.77,78 Otras isoformas del gen que codifica para el intercambiador Na+/Ca2+ se han clonado en la Drosophila melanogaster (Dmel/Ncx),66 el nemátodo Caenorhabditis elegans79 y el calamar (NCX-SQ1).65 La isoforma NCX1 posee sitios de splicing alternativos en 6 exones: A, B, C, D, E y F.80 Sin embargo, todas las variantes incluyen a los exones A o B, que se excluyen mutuamente. Las isoformas con combinaciones de exones diferentes presentan variabilidades especie-específicas y expresiones tejido-específicas (tabla 1).80-82 Los tejidos excitables poseen usualmente intercambiadores que contienen el exón A, mientras que en otros tejidos predomina el exón B. Por ejemplo: en el corazón se encuentra una isoforma única (NCX1.1, combinación de exones ACDEF) del intercambiador, mientras otros órganos como el riñón, el cerebro, los ojos, etc, presentan 2 o más isoformas.81,83 Hasta el momento, no se conoce el significado fisiológico y funcional de la existencia de diferentes variantes de splicing descritas.
Tabla 1. Expresión tejido-específica de las diferentes isoformas del intercambiador Na+/Ca2+ en ratas
| Isoformas NCX1 | ]]> 1 | 12 | 5 | 4 | 2 | 9 | 7 | 3 | 10 | 11 |
| Exones | ]]> ACDF | ADEF | ADF | AD | BCD | BDE | BDF | BD | BDEF | BCDEF |
| Corazón | ]]> Ö | |||||||||
| Músculo esquelético | ]]> Ö | Ö | Ö | Ö | ||||||
| Cerebro | ]]> | Ö | Ö | Ö | Ö | Ö | ||||
| Ojos | ]]> | Ö | Ö | Ö | ||||||
| Aorta | ]]> | Ö | Ö | Ö | ||||||
| Riñon | ]]> | Ö | Ö | |||||||
| Intestino | ]]> | Ö | Ö | |||||||
| Timo | ]]> | Ö | Ö | |||||||
| Bazo | ]]> | Ö | Ö | |||||||
| Células endoteliales | ]]> | Ö | Ö | |||||||
| Células b | ]]> | Ö | Ö |
Estructura y topología en la membrana del intercambiador Na+/Ca2+. Los modelos iniciales basados en análisis hidropáticos de la isoforma NCX1 del intercambiador Na+/Ca2+ sugerían que esta proteína poseía 12 segmentos transmembrana. Posteriormente, se encontró que el segmento hidrofóbico inicial de 32 aminoácidos es un péptido señal, el cual se separa de la proteína durante su procesamiento inicial en el retículo endoplasmático.84,85 En la actualidad, se plantea que el intercambiador posee solamente 9 segmentos transmembrana.86 La región N-terminal de esta proteína está glicosilada en la posición 9, y por lo tanto, es extracelular. Se ha descrito que la glicosilación85 y la hidrólisis del péptido señal no influyen significativamente en la función de este transportador.87,88
El intercambiador Na+/Ca2+ posee dos dominios hidrofóbicos que contienen los fragmentos transmembrana. Estos dominios están separados por un lazo intracelular largo constituido por 550 aminoácidos (figura 4). El dominio hidrofóbico N-terminal posee 5 segmentos transmembrana y el C-terminal está formado por 4 segmentos de este tipo. Mediante la eliminación del lazo intracelular, se demostró que esta región no es esencial para el transporte iónico y que los segmentos transmembranas son los responsables de la función de la proteína. Sin embargo, el lazo intracelular contiene secuencias relacionadas con funciones reguladoras, tales como una zona de splicing alternativo y un dominio de unión a Ca2+.
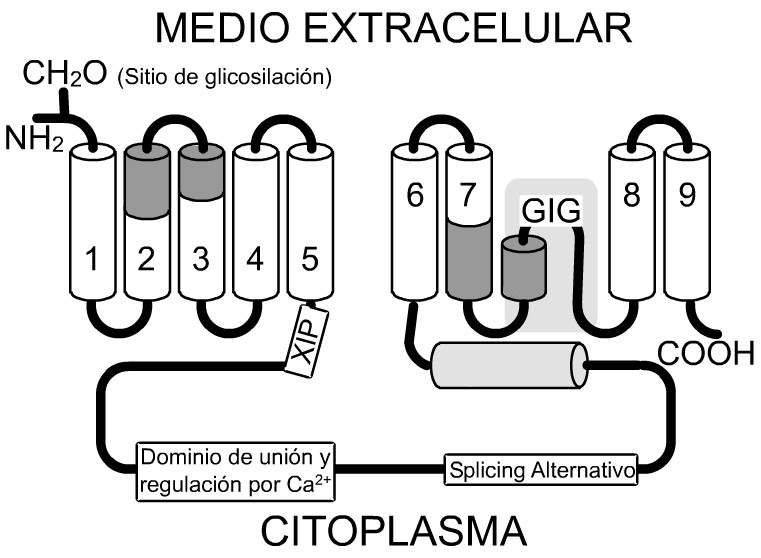
FIG. 4. Modelo actual de la topología del intercambiador Na+/Ca2+. La proteína posee 9 segmentos, los cuales atraviesan la membrana plasmática. El extremo amino-terminal se glicosila y es extracelular. Entre los segmentos 7 y 8 hay un fragmento hidrofóbico con estructura similar al lazo P presente en los canales de K+ (área sombreada con gris claro, GIG: Gli-Ile-Gli). Las regiones homólogas a, las cuales intervienen directamente en el transporte iónico y se sitúan en los segmentos transmembrana 2,3 y 7, se indican con sombreado gris oscuro. Se muestran además en el lazo intracelular la región reconocida por el péptido inhibitorio XIP, el sitio de unión al Ca2+ regulatorio y la región correspondiente a la secuencia de ARN donde ocurre el splicing alternativo. El lazo citoplasmático no está dibujado a escala, pues constituye más de la mitad de la longitud de la proteína (550 aminoácidos). Tomado de: (Philipson y Nicoll, 2000).
Existen regiones del intercambiador Na+/Ca2+ que muestran una homología intramolecular. La secuencia que abarca una parte de los segmentos transmembrana 2 y 3 es muy similar a la secuencia correspondiente al segmento 7 (figura 4). Las regiones homólogas anteriores se denominaron a-1 y a-2, respectivamente, y aparentemente desempeñan un papel importante en el transporte iónico. Esta homología indica que durante la evolución del intercambiador ocurrió un evento de duplicación génica. La actividad de intercambio Na+/Ca2+ es muy sensible a mutaciones en las regiones a. Por ejemplo, un cambio del aminoácido serina por alanina, cisteína o treonina en la posición 110 inactiva al intercambiador Na+/Ca2+. Lo mismo ocurre cuando se sustituye el ácido glutámico por el ácido aspártico o la glutamina en la posición 113. Algunas mutaciones en la región cercana al citoplasma del segmento 2 alteran la selectividad a iones.89 Normalmente, el intercambiador utiliza solamente el ion sodio para realizar su función y este requerimiento es muy riguroso. La sustitución de la treonina por la valina en la posición 103 no solo permite la realización de intercambio Na+/Ca2+, sino también Li+/Ca2+.
Existen también secuencias homólogas en el lazo intracelular. Estas poseen un tamaño aproximado de 60 aminoácidos y se denominaron ß-1 y ß-2. Sin embargo, hasta el presente no se conoce la importancia de estas regiones en la función de la proteína. Generalmente, el intercambio Na+/Ca2+ no se afecta por mutaciones en el lazo intracelular, aunque algunas de estas pueden influir sobre las propiedades reguladoras del propio Ca2+ 72. Por ejemplo, el intercambiador Na+/Ca2+ se inactiva si no hay Ca2+ unido a un sitio localizado en el lazo intracelular. Algunos autores han propuesto que las concentraciones de Ca2+ inferiores a 100-300 nmol/L 72 pueden inactivar el intercambiador Na+/Ca2+ y otros sugirieron que estas pueden ser menores (20-50 nmol/L).90,91 Si estos últimos valores fueran las concentraciones de Ca2+ requeridas para la activación del intercambio Na+/Ca2+, entonces el sitio del lazo intracelular regulado por el Ca2+ quizás estuviera siempre saturado, y su relevancia en la regulación de la función de la proteína fuera insignificante. Si lo fuesen los primeros, el intercambiador sólo se activaría cuando se elevan las concentraciones de Ca2+ citosólico, como ocurre durante la contracción del músculo cardíaco. Hasta el presente, no existe un consenso en relación con estos valores.
El intercambiador Na+/Ca2+ en el control del acoplamiento de la excitación-contracción del músculo cardíaco. En el músculo cardíaco, el acoplamiento de la excitación-contracción comienza cuando se activan los canales de Ca2+ tipo L por la despolarización del potencial de membrana.95 La entrada de Ca2+ a través de estos canales incrementa localmente la concentración intracelular de Ca2+, hasta valores suficientes para activar además la liberación de Ca2+ del RS por los canales RyR, mediante un fenómeno conocido como "liberación de Ca2+ inducida por Ca2+" 96,97. El intercambiador Na+/Ca2+ extrude el Ca2+ que entra a la célula desde el espacio extracelular, y este es distribuido por toda la superficie extracelular, incluyendo los túbulos transversales.98 La eficiencia del intercambiador Na+/Ca2+ como principal ejecutor dentro del mecanismo de extrusión de Ca2+ resulta posiblemente de su densa distribución (aproximadamente de 250 a 400 unidades de intercambiador Na+/Ca2+ por micrómetro cuadrado).69,99,100
El intercambiador Na+/Ca2+ puede además operar en el modo reverso y suministrar el Ca2+ necesario para activar los canales RyR, del mismo modo que lo hacen los canales de Ca2+ tipo-L. Esto posiblemente ocurriría después de la despolarización, cuando los niveles locales de Na+ intracelular pudieran favorecer la entrada de Ca2+ a través del intercambiador Na+/Ca2+.59,101,102
El retículo sarcoplasmático de los cardiomiocitos está poco desarrollado en muchos mamíferos neonatos. El acoplamiento excitación-contracción de las células cardíacas depende, en gran medida, de los flujos de Ca2+ a través de los canales de Ca2+ y el intercambiador Na+/Ca2+ durante el inicio de la vida. De hecho, la expresión del intercambiador Na+/Ca2+ cardíaco es mayor en mamíferos neonatos y disminuye rápidamente con la edad. La disminución de la actividad de intercambio Na+/Ca2+ es simultánea al desarrollo del retículo sarcoplasmático y los túbulos transversos.
El intercambiador Na+/Ca2+ neuronal. Imágenes obtenidas mediante técnicas de inmunofluorescencia han sugerido que existe una alta concentración del intercambiador Na+/Ca2+ en las zonas de sinapsis neuronal y en las uniones neuromusculares .103 Debido a que las señales locales de Ca2+ parecen ser importantes en la sinapsis neuronal, probablemente el intercambiador Na+/Ca2+ participe en la extrusión de Ca2+ que entra a través de los canales de Ca2+ de la membrana plasmática.104
El intercambiador Na+/Ca2+ en el riñón. El riñón, junto al corazón y al cerebro, son 3 fuentes ricas en proteína del intercambiador Na+/Ca2+. En el riñón, la más alta concentración del intercambiador Na+/Ca2+ se localiza en el túbulo colector de la neurona;81,105 sin embargo, los detalles referentes a su función no se conocen totalmente. Se han descrito diferentes isoformas del intercambiador Na+/Ca2+ en el riñón106 y estas pudieran explicar funciones diferentes para este tejido.
El intercambiador Na+/Ca2+ en las células ß del páncreas. En las células ß pancreáticas el intercambiador Na+/Ca2+ participa de forma importante en el control de la concentración citoplasmática de Ca2+.107-109 Recientemente, se identificaron dos isoformas del intercambiador (NCX1.3 y NCX1.7) en la célula ß pancreática, y su papel en la fisiología de esta fue caracterizado.108,110 Mediante la utilización de oligonucleótidos que contenían una secuencia antisentido a la del ARNm que codifica para el intercambiador Na+/Ca2+, con el objetivo de bloquear la biosíntesis de la proteína, los autores de estos estudios demostraron que el intercambio Na+/Ca2+ puede contribuir hasta en un 70 % al retorno de las concentraciones citosólicas de Ca2+ ([Ca2+]i) a sus valores basales cuando cesa un estímulo despolarizante.
Se describió además que esta contribución a la extrusión de Ca2+ a través del intercambio Na+/Ca2+ se hace significativa a una concentración de Ca2+ citosólico alrededor de 200-500 nmol/L, lo cual implica la participación limitada del intercambiador Na+/Ca2+ en el mantenimiento de la [Ca2+]i basal (~150 nmol/L) (Van Eylen et al., 1998). Este hallazgo concuerda con la baja afinidad por el Ca2+ referida para esta proteína. Sin embargo, está claro que el intercambiador Na+/Ca2+ participa activamente en la restauración de las [Ca2+]i y este es complementado por otros efectores, como son los tampones intracelulares de Ca2+ y las Ca2+-ATPasa de la membrana plasmática y del retículo endoplasmático. Por otra parte, el intercambiador Na+/Ca2+ puede funcionar en el modo reverso, lo que permite la incorporación de Ca2+ desde el medio extracelular bajo determinadas condiciones, como son la despolarización de la membrana plasmática y la disminución o la inversión del gradiente de Na+.
Apoptosis, definición y características. La apoptosis es un término empleado para describir un modo de muerte celular que se caracteriza por la constricción de la célula, la condensación del núcleo (sin afectar la permeabilidad de la membrana citoplasmática) y la fragmentación de la célula, lo cual conduce a la formación de los cuerpos apoptóticos.111 El destino de los cuerpos apoptóticos es un fenómeno de gran importancia biológica; ellos son englobados por fagocitos "profesionales", como pueden ser las células epiteliales adyacentes, sin provocar una respuesta inflamatoria porque conservan su contenido hasta que ocurre la fagocitosis.112 De esta forma, el daño accidental de los tejidos causado por la liberación de enzimas de los gránulos está minimizado. Consecuentemente, esta forma de muerte celular participa en la remodelación de los tejidos, prerequisito importante para un desarrollo embriológico normal. Por esta razón, el término apoptosis se intercambia de forma habitual con el de "muerte celular programada". La apoptosis ocurre donde exista proliferación celular y es controlada por genes compartidos que participan en el ciclo celular. La relación estrecha entre el control de la apoptosis y la proliferación significa que un defecto en alguna de ellas probablemente de lugar a una alteración del número de células. En realidad, algunos genes involucrados en la inducción de la apoptosis (p53)113 se reconocen como clásicos genes supresores de tumores, mientras otros que previenen la apoptosis e incrementan la proliferación celular (blc2)114 son oncogenes clásicos. La apoptosis se diferencia de forma significativa de la necrosis, ya que esta última es un evento accidental donde la célula pierde la capacidad de mantener su vida, pues el resto de los procesos vitales se desacoplan de la respiración, se pierde energía y finalmente se degrada de forma incontrolada. Esto provoca la pérdida del contenido celular, que contiene además moléculas reactivas y enzimas activas.114 La liberación de estas últimas hacia el intersticio representa un potente estímulo para que ocurra una inflamación aguda que conduce a un daño tisular adicional.
La regulación de la apoptosis y sus principales vías. El concepto filosófico que proclama a "la muerte como una condición esencial de la vida" se evidencia en el hecho de que la apoptosis es necesaria para el desarrollo normal de los organismos pluricelulares.115 Pero también la apoptosis es un "arma de doble filo", y el descontrol de este tipo de muerte celular está implicado en numerosas patologías;116 por esta razón, este proceso debe estar estrechamente regulado. Uno de los elementos implicados en dicho control es la compartimentación de los componentes de la maquinaria apoptótica en diferentes organelos.
]]> Sólo cuando la señal de muerte es enviada, los instrumentos de su ejecución son llevados al citosol y el programa de suicidio es activado. Los organelos conocidos que participan en esta compartimentación son la membrana plasmática, donde se sitúan los receptores de muerte y supervivencia, la mitocondria, que alberga a algunas proteínas que regulan la apoptosis, y muy recientemente fue incluido el retículo endoplasmático, pues en él se localiza la enzima pro-apoptótica caspasa-12.117,118Las principales moléculas que participan en la ejecución de la muerte celular programada son las caspasas. Estas proteínas pertenecen a la familia de proteasas dependientes de cisteína y aspartato específicas.119 Según su función, se dividen en 2 grupos conocidos como caspasas iniciadoras (caspasas-8 y -9), cuya función es la de activar otras caspasas, y las caspasas ejecutoras (caspasas-3, -6 y -7), que son responsables de la digestión de proteínas celulares, tanto funcionales como estructurales.
Las caspasas de mamíferos son análogas al producto del gen de muerte celular Ced-3 del nemátodo Caenorhabditis elegans. Estas proteasas normalmente se encuentran en la célula en forma de proenzimas inactivas (procaspasas), y pueden ser activadas mediante un corte proteolítico realizado por un complejo de activación o por otra caspasa.120 Se ha descrito que la activación de las caspasas contribuye a la muerte celular en la isquemia cerebral,121 en la isquemia cardíaca122 y en enfermedades crónicas neurodegenerativas, como la enfermad de Alzheimer123 y de Huntington.124
La activación de las caspasas se realiza por 2 vías, denominadas extrínseca e intrínseca. La vía extrínseca consiste en la transducción de señales a través de receptores de muerte, la cual provoca la activación de la caspasa-8. Esta activación ocurre si los receptores de muerte, como el Fas y el receptor del factor de necrosis tumoral, se oligomerizan después de la unión a sus ligandos específicos. A continuación, la caspasa-8 produce la activación de caspasas efectoras como son las caspasa-3 y -7125. La otra vía más estudiada, la intrínseca, se inicia con la liberación de una proteína mitocondrial: el citocromo-c, el cual forma un complejo con dATP y la proteína Apaf-1.126,127 Este complejo une a la caspasa-9 y le produce un corte proteolítico. La caspasa-9 activa es liberada del complejo y cataliza la activación de las caspasas efectoras
-3, -6 y -7.127,128 Las caspasas efectoras activadas, ya sea por la vía extrínseca como por la intrínseca, son capaces de hidrolizar proteínas estructurales del citoesqueleto y del núcleo celular, y además, proteínas funcionales como la poli-ADP-ribosa polimerasa.129 Estas cascadas de reacciones conducen finalmente a la muerte celular.
Recientemente, la activación de la caspasa-12 se asoció específicamente al estrés del retículo endoplasmático.118 El tratamiento de las células con un inhibidor del transporte desde el retículo endoplasmático hacia el complejo de Golgi: la brefeldina A, y con un inhibidor de la N-glucosilación en el retículo endoplasmático: la tunicamicina, provoca la activación de la caspasa-12. Sin embargo, la activación más significativa de la caspasa-12 se obtiene al exponer las células a un inhibidor de la Ca2+-ATPasa del retículo endoplasmático: la tapsigargina. La apoptosis inducida por estímulos que no afectan al retículo endoplasmático, como es la supresión de suero o activación del Fas, no resulta en una activación de la caspasa-12.
La caspasa-12 forma parte de una subfamilia de caspasas denominada ICE (interleukin-1ß converting enzime, en inglés).118 La secuencia aminoacídica de la caspasa-12 murina contiene secuencias homólogas a las caspasas murinas -1 y -11 (39 % y 38 % de identidad, respectivamente) y a las caspasas humanas -4 y -5 (48 % y 45 % de identidad, respectivamente). La caspasa-12 está presente en todos los tejidos de ratones, aunque la expresión es superior en el músculo, el hígado y el riñón. Esta proteasa se localizó en las fracciones microsomales y solubles de células nerviosas mediante la utilización de una técnica de fraccionamiento celular.118 Sin embargo, la forma inactiva de esta proteína, las procaspasa
-12 (60 kDa de peso molecular relativo) se detectó predominantemente en la fracción microsomal, mientras que su forma activa, la caspasa-12 (36 kDa de peso molecular relativo), se localizó en la fracción soluble. Estas determinaciones sugieren que la procaspasa-12 está asociada al retículo endoplasmático y que la caspasa-12 se liberó durante el proceso de fraccionamiento celular. Además, la asociación específica de esta proenzima con el retículo endoplasmático se comprobó por inmunohistoquímica mediante la utilización de anticuerpos monoclonales que reconocen a esta proteína y a proteínas asociadas a este organelo.118
Por otra parte, se determinó que la activación de la caspasa-12 no ocurre a través de la vía extrínseca descrita anteriormente, pues la exposición de timocitos aislados de ratones transgénicos que expresan o no la procaspasa-12 (procaspasa-12 +/+ y procaspasa-12 -/-, respectivamente) a un anticuerpo que reconoce al receptor Fas, produjo decrementos similares de la vitalidad en ambos grupos de células. La participación de la caspasa-12 en la vía intrínseca o mitocondrial también fue descartada. Para demostrar esta hipótesis se realizaron estudios en fibroblastos Sack2 (Apaf-1 -/-) los cuales carecen de la proteína Apaf-1. Las células Sack 2 son resistentes al tamoxifeno, un activador de la vía apoptótica mitocondrial; sin embargo, la exposición a la brefeldina A y a la tapsigargina redujeron la vitalidad de estas células.118
La sobrecarga de Ca2+ ha sido relacionada con el daño celular en muchos tipos de tejidos. En las neuronas, por ejemplo, la neurotoxicidad inducida por glutamato está asociada con un incremento prolongado de las [Ca2+]i y se plantea que la acumulación excesiva de este ion en el citosol produce pérdidas neuronales severas como la isquemia y el trauma cerebral,130,131 además de enfermedades degenerativas como la de Alzheimer.132,133 Otros estudios han mostrado que la prevención de la sobrecarga de Ca2+, mediante el pretratamiento con agentes quelantes134 y bloqueadores de canales de Ca2+ 135, puede poseer efectos protectores sobre las células. De forma similar, se ha comprobado que las células que expresan altos niveles de una proteína que une Ca2+ (calbindina-D28K), son más resistentes a la muerte celular.136 ]]>
Se plantea que la sobrecarga de Ca2+ puede mediar la muerte celular del tipo necrótico, aunque existen numerosas pruebas que involucran al Ca2+ en la apoptosis. Por ejemplo, el corte de la cromatina nuclear en fragmentos oligonucleosómicos, característico en el proceso apoptótico, es el resultado de la activación de una endonucleasa dependiente de Ca2+ 138. Además, se ha descrito que el incremento del ion calcio puede comenzar en el núcleo de algunos tipos celulares, lo cual sugiere que un aumento selectivo de la concentración del ion divalente en esa estructura pudiera ser suficiente para el inicio de la fragmentación del ADN; Sin embargo, no resulta aun claro si la fragmentación del ADN mediada por Ca2+ comienza con la activación de algunas endonucleasas o con la alteración de la superestructura de la cromatina, lo cual pudiera hacer accesible sitios adicionales de las cadenas de ADN a la digestión por endonucleasas. En realidad, algunas determinaciones indican que el corte del ADN en fragmentos internucleosómicos es secundario a la condensación de la cromatina y al debilitamiento de la interacción histona-ADN.137
Recientemente, existe una creciente acumulación de pruebas que demuestran que la disminución de la concentración de Ca2+ en el lumen del retículo endoplasmático produce un estado de estrés en este organelo con la consiguiente afectación de sus funciones, dentro de las cuales se encuentran la síntesis, el plegamiento y el transporte de proteínas.139 Este evento se ha relacionado con la activación de una vía apoptótica mediada por la proteasa caspasa-12117,118 y la inducción de un factor de transcripción codificado por el gen CHOP, también conocido como GADD153 (growth arrest and DNA-damage-inducible gene, en inglés) el cual es capaz de detener el crecimiento celular y provocar daños en el ADN nuclear.139,140
The present paper made a review of the present knowledge on regulation of intracellular concentrations of calcium ion, the main mechanisms of inlet/outlet through the plasma membrane, with special attention to Na+/Ca2+, and the function of this important second messenger in insulin secretion and in programmed cell death of pancreatic cells.
Key words: Calcium, Na+/Ca2+ exchange; apoptosis; insulin
1. Clapham DE. Calcium signaling. Cell 1995;80:259-268.
2. Berridge MJ. Cell signalling. A tale of two messengers. Nature 1993;365:388-389.
3. Lomax RB, Camello C, Van Coppenolle F, Petersen OH, Tepikin AV. Basal and physiological Ca(2+) leak from the endoplasmic reticulum of pancreatic acinar cells. Second messenger-activated channels and translocons. J Biol Chem 2002;277:26479-26485.
4. Berridge MJ, Irvine RF. Inositol phosphates and cell signalling. Nature 1989;341:197-205.
5. Mignery GA, Sudhof TC. The ligand binding site and transduction mechanism in the inositol-1,4,5-triphosphate receptor. EMBO J 1990;9:3893-3898.
6. Combettes L, Champeil P. Calcium and inositol 1,4,5-trisphosphate-induced Ca2+ release. Science 1994;265:813-815.
7. Stehno-Bittel L, Luckhoff A, Clapham DE. Calcium release from the nucleus by InsP3 receptor channels. Neuron 1995;14:163-167.
8. Putney JW Jr. Capacitative calcium entry revisited. Cell Calcium 1990;11:611-624.
9. Lechleiter JD, Clapham DE. Molecular mechanisms of intracellular calcium excitability in X. laevis oocytes. Cell 1992;69:283-294.
10. Hoth M, Penner R. Depletion of intracellular calcium stores activates a calcium current in mast cells. Nature 1992;355:353-356.
11. Zweifach A, Lewis RS. Mitogen-regulated Ca2+ current of T lymphocytes is activated by depletion of intracellular Ca2+ stores. Proc Natl Acad Sci U S A 1993;90:6295-6299.
12. Luckhoff A, Clapham DE. Calcium channels activated by depletion of internal calcium stores in A431 cells. Biophys J 1994;67:177-182.
13. Tengholm A, Hagman C, Gylfe E, Hellman B. In situ characterization of nonmitochondrial Ca2+ stores in individual pancreatic beta-cells. Diabetes 1998;47:1224-1230.
14. Baimbridge KG. Calcium-binding proteins in the dentate gyrus. Epilepsy Res 1992;7:211-220.
15. Pozzan T, Rizzuto R, Volpe P, Meldolesi J. Molecular and cellular physiology of intracellular calcium stores. Physiol Rev 1994;74:595-636.
16. Sugden MC, Christie MR, Ashcroft SJ. Presence and possible role of calcium-dependent regulator (calmodulin) in rat islets of Langerhans. FEBS Lett 1979;105:95-100.
17. Lu KP, Means AR. Regulation of the cell cycle by calcium and calmodulin. Endocr Rev 1993;14:40-58.
18. Kretsinger RH. Structure and evolution of calcium-modulated proteins. CRC Crit Rev Biochem 1980;8:119-174.
19. Malaisse WJ, Blachier F, Pochet R, Keenoy B, Sener A. Calmodulin and calbindin in pancreatic islet cells. Adv Exp Med Biol 1990;269:127-133.
20. Penn EJ, Brocklehurst KW, Sopwith AM, Hales CN, Hutton JC. Ca2+--Calmodulin dependent myosin light-chain phosphorylating activity in insulin-secreting tissues. FEBS Lett 1982;139:4-8.
21. MacDonald MJ, Kowluru A. Calcium-calmodulin-dependent myosin phosphorylation by pancreatic islets. Diabetes 1982;31:566-570.
22. Iida Y, Senda T, Matsukawa Y, Onoda K, Miyazaki JI, Sakaguchi H et al. Myosin light-chain phosphorylation controls insulin secretion at a proximal step in the secretory cascade. Am J Physiol 1997;273:E782-E789.
23. Landt M, McDaniel ML, Bry CG, Kotagal N, Colca JR, Lacy PE et al. Calmodulin-activated protein kinase activity in rat pancreatic islet cell membranes. Arch Biochem Biophys 1982;213:148-154.
24. Norling LL, Colca JR, Brooks CL, Kloepper RF, McDaniel ML, Landt M. Specificity of inhibition of calcium- and calmodulin-dependent protein kinase by alloxan. Biochim Biophys Acta 1984;801:197-205.
25. Wenham RM, Landt M, Easom RA. Glucose activates the multifunctional Ca2+/calmodulin-dependent protein kinase II in isolated rat pancreatic islets. J Biol Chem 1994;269:4947-4952.
26. Mohlig M, Wolter S, Mayer P, Lang J, Osterhoff M, Horn PA et al. Insulinoma cells contain an isoform of Ca2+/calmodulin-dependent protein kinase II delta associated with insulin secretion vesicles. Endocrinology 1997;138:2577-2584.
27. Krueger KA, Bhatt H, Landt M, Easom RA. Calcium-stimulated phosphorylation of MAP-2 in pancreatic betaTC3-cells is mediated by Ca2+/calmodulin-dependent kinase II. J Biol Chem 1997;272:27464-27469.
28. Krueger KA, Ings EI, Brun AM, Landt M, Easom RA. Site-specific phosphorylation of synapsin I by Ca2+/calmodulin-dependent protein kinase II in pancreatic betaTC3 cells: synapsin I is not associated with insulin secretory granules. Diabetes 1999;48:499-506.
29. Greengard P, Valtorta F, Czernik AJ, Benfenati F. Synaptic vesicle phosphoproteins and regulation of synaptic function. Science 1993;259:780-785.
30. Yamamoto H, Fukunaga K, Goto S, Tanaka E, Miyamoto E. Ca2+, calmodulin-dependent regulation of microtubule formation via phosphorylation of microtubule-associated protein 2, tau factor, and tubulin, and comparison with the cyclic AMP-dependent phosphorylation. J Neurochem 1985;44:759-768.
31. Wollheim CB, Lang J, Regazzi R. The exocytotic process of insulin secretion and its regulation by Ca2+ and G-proteins. Diabetes Rev 1996;4:277-97.
32. Hay JC, Martin TF. Resolution of regulated secretion into sequential MgATP-dependent and calcium-dependent stages mediated by distinct cytosolic proteins. J Cell Biol 1992;119:139-151.
33. Lang J, Fukuda M, Zhang H, Mikoshiba K, Wollheim CB. The first C2 domain of synaptotagmin is required for exocytosis of insulin from pancreatic beta-cells: action of synaptotagmin at low micromolar calcium. EMBO J 1997;16:5837-5846.
34. Edwardson JM, An S, Jahn R. The secretory granule protein syncollin binds to syntaxin in a Ca2(+)-sensitive manner. Cell 1997;90:325-333.
35. Bokvist K, Eliasson L, Ammala C, Renstrom E, Rorsman P. Co-localization of L-type Ca2+ channels and insulin-containing secretory granules and its significance for the initiation of exocytosis in mouse pancreatic B-cells. EMBO J 1995;14:50-57.
36. Eliasson L, Renstrom E, Ding WG, Proks P, Rorsman P. Rapid ATP-dependent priming of secretory granules precedes Ca(2+)-induced exocytosis in mouse pancreatic B-cells. J Physiol 1997;503 ( Pt 2):399-412.
37. Renstrom E, Eliasson L, Bokvist K, Rorsman P. Cooling inhibits exocytosis in single mouse pancreatic B-cells by suppression of granule mobilization. J Physiol 1996;494 ( Pt 1):41-52.
38. Easom RA, Tarpley JL, Filler NR, Bhatt H. Dephosphorylation and deactivation of Ca2+/calmodulin-dependent protein kinase II in betaTC3-cells is mediated by Mg2+- and okadaic-acid-sensitive protein phosphatases. Biochem J 1998;329 ( Pt 2):283-288.
39. Maccioni RB, Cambiazo V. Role of microtubule-associated proteins in the control of microtubule assembly. Physiol Rev 1995;75:835-864.
40. Suprenant KA, Dentler WL. Association between endocrine pancreatic secretory granules and in-vitro-assembled microtubules is dependent upon microtubule-associated proteins. J Cell Biol 1982;93:164-174.
41. Easom RA. CaM kinase II: a protein kinase with extraordinary talents germane to insulin exocytosis. Diabetes 1999;48:675-684.
42. Sattilaro RF. Interaction of microtubule-associated protein 2 with actin filaments. Biochemistry 1986;25:2003-2009.
43. Selden SC, Pollard TD. Interaction of actin filaments with microtubules is mediated by microtubule-associated proteins and regulated by phosphorylation. Ann N Y Acad Sci 1986;466:803-812.
44. Hisatomi M, Hidaka H, Niki I. Ca2+/calmodulin and cyclic 3,5' adenosine monophosphate control movement of secretory granules through protein phosphorylation/dephosphorylation in the pancreatic beta-cell. Endocrinology 1996;137:4644-4649.
45. Howell SL, Tyhurst M. The cytoskeleton and insulin secretion. Diabetes Metab Rev 1986;2:107-123.
46. Easom RA, Filler NR, Ings EM, Tarpley J, Landt M. Correlation of the activation of Ca2+/calmodulin-dependent protein kinase II with the initiation of insulin secretion from perifused pancreatic islets. Endocrinology 1997;138:2359-2364.
47. Yano K, Zarain-Herzberg A. Sarcoplasmic reticulum calsequestrins: structural and functional properties. Mol Cell Biochem 1994;135:61-70.
48. Wu KD, Lee WS, Wey J, Bungard D, Lytton J. Localization and quantification of endoplasmic reticulum Ca(2+)-ATPase isoform transcripts. Am J Physiol 1995;269:C775-C784.
49. Nash PD, Opas M, Michalak M. Calreticulin: not just another calcium-binding protein. Mol Cell Biochem 1994;135:71-78.
50. Carafoli E. The signaling function of calcium and its regulation. J Hypertens 1994;12:S47-S56.
51. Kamagate A, Herchuelz A, Bollen A, Van Eylen F. Expression of multiple plasma membrane Ca(2+)-ATPases in rat pancreatic islet cells. Cell Calcium 2000;27:231-246.
52. Strehler EE, Zacharias DA. Role of alternative splicing in generating isoform diversity among plasma membrane calcium pumps. Physiol Rev 2001;81:21-50.
53. Penniston JT, Enyedi A. Modulation of the plasma membrane Ca2+ pump. J Membr Biol 1998;165:101-109.
54. Missiaen L, Raeymaekers L, Wuytack F, Vrolix M, De Smedt H, Casteels R. Phospholipid-protein interactions of the plasma-membrane Ca2+-transporting ATPase. Evidence for a tissue-dependent functional difference. Biochem J 1989;263:687-694.
55. Hao L, Rigaud JL, Inesi G. Ca2+/H+ countertransport and electrogenicity in proteoliposomes containing erythrocyte plasma membrane Ca-ATPase and exogenous lipids. J Biol Chem 1994;269:14268-14275.
56. Clarke DM, Loo TW, Inesi G, MacLennan DH. Location of high affinity Ca2+-binding sites within the predicted transmembrane domain of the sarcoplasmic reticulum Ca2+-ATPase. Nature 1989;339:476-478.
57. Mikkelsen EO, Thastrup O, Christensen SB. Effects of thapsigargin in isolated rat thoracic aorta. Pharmacol Toxicol 1988;62:7-11.
58. Philipson KD, Nicoll DA. Sodium-calcium exchange. Curr Opin Cell Biol 1992;4:678-683.
59. Lipp P, Niggli E. Sodium current-induced calcium signals in isolated guinea-pig ventricular myocytes. J Physiol 1994;474:439-446.
60. Baker PF, Blaustein MP, Hodgkin AL, Steinhardt RA. The influence of calcium on sodium efflux in squid axons. J Physiol 1969;200:431-458.
61. Reuter H, Seitz N. The dependence of calcium efflux from cardiac muscle on temperature and external ion composition. J Physiol 1968;195:451-470.
62. Kofuji P, Hadley RW, Kieval RS, Lederer WJ, Schulze DH. Expression of the Na-Ca exchanger in diverse tissues: a study using the cloned human cardiac Na-Ca exchanger. Am J Physiol 1992;263:C1241-C1249.
63. Komuro I, Wenninger KE, Philipson KD, Izumo S. Molecular cloning and characterization of the human cardiac Na+/Ca2+ exchanger cDNA. Proc Natl Acad Sci U S A 1992;89:4769-4773.
64. Nicoll DA, Longoni S, Philipson KD. Molecular cloning and functional expression of the cardiac sarcolemmal Na+-Ca2+ exchanger. Science 1990;250:562-565.
65. He Z, Tong Q, Quednau BD, Philipson KD, Hilgemann DW. Cloning, expression, and characterization of the squid Na+-Ca2+ exchanger (NCX-SQ1). J Gen Physiol 1998;111:857-873.
66. Ruknudin A, Valdivia C, Kojufi P, Lederer WJ, Schulze DH. Na+/Ca2+ exchanger in Drosophila: cloning, expression, and transport differences. Am J Physiol 1997;273:C257-C265.
67. Philipson KD. Sodium-calcium exchange in plasma membrane vesicles. Annu Rev Physiol 1985;47:561-571.
68. Eisner DA, Lederer WJ. Na-Ca exchange: stoichiometry and electrogenicity. Am J Physiol 1985;248:C189-C202.
69. Hilgemann DW, Nicoll DA, Philipson KD. Charge movement during Na+ translocation by native and cloned cardiac Na+/Ca2+ exchanger. Nature 1991;352:715-718.
70. Fujioka Y, Komeda M, Matsuoka S. Stoichiometry of Na+-Ca2+ exchange in inside-out patches excised from guinea-pig ventricular myocytes. J Physiol 2000;523 (Pt 2):339-351.
71. Khananshvili D. Distinction between the two basic mechanisms of cation transport in the cardiac Na(+)-Ca2+ exchange system. Biochemistry 1990;29:2437-2442.
72. Philipson KD, Nicoll DA. Sodium-calcium exchange: a molecular perspective. Annu Rev Physiol 2000;62:111-133.
73. Hilgemann DW. Unitary cardiac Na+, Ca2+ exchange current magnitudes determined from channel-like noise and charge movements of ion transport. Biophys J 1996;71:759-768.
74. Kappl M, Hartung K. Rapid charge translocation by the cardiac Na(+)-Ca2+ exchanger after a Ca2+ concentration jump. Biophys J 1996;71:2473-2485.
75. Khananshvili D. Mechanism of partial reactions in the cardiac Na(+)-Ca2+ exchange system. Ann N Y Acad Sci 1991;639:85-95.
76. Li JM, Kimura J. Translocation mechanism of cardiac Na-Ca exchange. Ann N Y Acad Sci 1991;639:48-60.
77. Li Z, Matsuoka S, Hryshko LV, Nicoll DA, Bersohn MM, Burke EP et al. Cloning of the NCX2 isoform of the plasma membrane Na(+)-Ca2+ exchanger. J Biol Chem 1994;269:17434-17439.
78. Nicoll DA, Quednau BD, Qui Z, Xia YR, Lusis AJ, Philipson KD. Cloning of a third mammalian Na+-Ca2+ exchanger, NCX3. J Biol Chem 1996;271:24914-24921.
79. Philipson KD, Nicoll DA, Matsuoka S, Hryshko LV, Levitsky DO, Weiss JN. Molecular regulation of the Na+-Ca2+ exchanger. Ann N Y Acad Sci 1996;779:20-28.
80. Kojufi P, Lederer WJ, Schulze DH. Mutually exclusive and cassette exons underlie alternatively spliced isoforms of the Na/Ca exchanger. J Biol Chem 1994;269:5145-5149.
81. Lee SL, Yu AS, Lytton J. Tissue-specific expression of Na+-Ca2+ exchanger isoforms. J Biol Chem 1994;269:14849-14852.
82. Oshima T, Ikeda K, Furukawa M, Takasaka T. Alternatively spliced isoforms of the Na+/Ca2+ exchanger in the guinea pig cochlea. Biochem Biophys Res Commun 1997;233:737-741.
83. Quednau BD, Nicoll DA, Philipson KD. Tissue specificity and alternative splicing of the Na+/Ca2+ exchanger isoforms NCX1, NCX2, and NCX3 in rat. Am J Physiol 1997;272:C1250-C1261.
84. Durkin JT, Ahrens DC, Pan YC, Reeves JP. Purification and amino-terminal sequence of the bovine cardiac sodium-calcium exchanger: evidence for the presence of a signal sequence. Arch Biochem Biophys 1991;290:369-375.
85. Hryshko LV, Nicoll DA, Weiss JN, Philipson KD. Biosynthesis and initial processing of the cardiac sarcolemmal Na(+)-Ca2+ exchanger. Biochim Biophys Acta 1993;1151:35-42.
86. Nicoll DA, Ottolia M, Lu L, Lu Y, Philipson KD. A new topological model of the cardiac sarcolemmal Na+-Ca2+ exchanger. J Biol Chem 1999;274:910-917.
87. Furman I, Cook O, Kasir J, Low W, Rahamimoff H. The putative amino-terminal signal peptide of the cloned rat brain Na(+)-Ca2+ exchanger gene (RBE-1) is not mandatory for functional expression. J Biol Chem 1995;270:19120-19127.
88. Loo TW, Ho C, Clarke DM. Expression of a functionally active human renal sodium-calcium exchanger lacking a signal sequence. J Biol Chem 1995;270:19345-19350.
89. Doering AE, Nicoll DA, Lu Y, Lu L, Weiss JN, Philipson KD. Topology of a functionally important region of the cardiac Na+/Ca2+ exchanger. J Biol Chem 1998;273:778-783.
90. Miura Y, Kimura J. Sodium-calcium exchange current. Dependence on internal Ca and Na and competitive binding of external Na and Ca. J Gen Physiol 1989;93:1129-1145.
91. Fang Y, Condrescu M, Reeves JP. Regulation of Na+/Ca2+ exchange activity by cytosolic Ca2+ in transfected Chinese hamster ovary cells. Am J Physiol 1998;275:C50-C55.
92. Cook O, Low W, Rahamimoff H. Membrane topology of the rat brain Na+-Ca2+ exchanger. Biochim Biophys Acta 1998;1371:40-52.
93. Santacruz-Toloza L, Ottolia M, Nicoll DA, Philipson KD. Functional analysis of a disulfide bond in the cardiac Na(+)-Ca(2+) exchanger. J Biol Chem 2000;275:182-188.
94. Walz T, Hirai T, Murata K, Heymann JB, Mitsuoka K, Fujiyoshi Y et al. The three-dimensional structure of aquaporin-1. Nature 1997;387:624-627.
95. Niggli E, Lederer WJ. Molecular operations of the sodium-calcium exchanger revealed by conformation currents. Nature 1991;349:621-624.
96. Bers DM. Species differences and the role of sodium-calcium exchange in cardiac muscle relaxation. Ann N Y Acad Sci 1991;639:375-385.
97. Fabiato A. Stimulated calcium current can both cause calcium loading in and trigger calcium release from the sarcoplasmic reticulum of a skinned canine cardiac Purkinje cell. J Gen Physiol 1985;85:291-320.
98. Kieval RS, Bloch RJ, Lindenmayer GE, Ambesi A, Lederer WJ. Immunofluorescence localization of the Na-Ca exchanger in heart cells. Am J Physiol 1992;263:C545-C550.
99. Nicoll DA, Philipson KD. Molecular studies of the cardiac sarcolemmal sodium-calcium exchanger. Ann N Y Acad Sci 1991;639:181-188.
100. Niggli E, Lederer WJ. Voltage-independent calcium release in heart muscle. Science 1990;250:565-568.
101. Lederer WJ, Cannell MB, Cohen NM, Berlin JR. Excitation-contraction coupling in heart muscle. Mol Cell Biochem 1989;89:115-119.
102. Lederer WJ, Niggli E, Hadley RW. Sodium-calcium exchange in excitable cells: fuzzy space. Science 1990;248:283-
103. Luther PW, Yip RK, Bloch RJ, Ambesi A, Lindenmayer GE, Blaustein MP. Presynaptic localization of sodium/calcium exchanger in neuromuscular preparations. J Neurosci 1992;12:4898-4904.
104. Bootman MD, Berridge MJ. The elemental principles of calcium signaling. Cell 1995;83:675-678.
105. Reilly RF, Shugrue CA, Lattanzi D, Biemesderfer D. Immunolocalization of the Na+/Ca2+ exchanger in rabbit kidney. Am J Physiol 1993;265:F327-F332.
106. Reilly RF, Shugrue CA. cDNA cloning of a renal Na+-Ca2+ exchanger. Am J Physiol 1992;262:F1105-F1109.
107. Van Eylen F, Gourlet P, Vandermeers A, Lebrun P, Herchuelz A. Inhibition of Na/Ca exchange by Phe-Met-Arg-Phe-NH2 (FMRFa)-related peptides in intact rat pancreatic B-cells. Mol Cell Endocrinol 1994;106:R1-R5.
108. Van Eylen F, Lebeau C, Albuquerque-Silva J, Herchuelz A. Contribution of Na/Ca exchange to Ca2+ outflow and entry in the rat pancreatic beta-cell: studies with antisense oligonucleotides. Diabetes 1998;47:1873-1880.
109. Plasman PO, Lebrun P, Cragoe EJ Jr. Herchuelz A. Inhibition of Na/Ca exchange in pancreatic islet cells by 3',4'-dichlorobenzamil. Biochem Pharmacol 1991;41:1759-1768.
110. Gall D, Gromada J, Susa I, Rorsman P, Herchuelz A, Bokvist K. Significance of Na/Ca exchange for Ca2+ buffering and electrical activity in mouse pancreatic beta-cells. Biophys J 1999;76:2018-2028.
111. Arends MJ, Wyllie AH. Apoptosis: mechanisms and roles in pathology. Int Rev Exp Pathol 1991;32:223-254.
112. Savill JS, Wyllie AH, Henson JE, Walport MJ, Henson PM, Haslett C. Macrophage phagocytosis of aging neutrophils in inflammation. Programmed cell death in the neutrophil leads to its recognition by macrophages. J Clin Invest 1989;83:865-875.
113. Harrison DJ, Howie SE, Wyllie AH. Lymphocyte death, p53, and the problem of the "undead" cell. Curr Top Microbiol Immunol 1995;200:123-135.
114. Malcomson RD, Oram SH, Harrison DJ. The importance of apoptosis: is it real or imaginary? Biologicals 1996;24:295-299.
115. Jacobson MD, Weil M, Raff MC. Programmed cell death in animal development. Cell 1997;88:347-354.
116. Thompson CB. Apoptosis in the pathogenesis and treatment of disease. Science 1995;267:1456-1462.
117. Mehmet H. Caspases find a new place to hide. Nature 2000;403:29-30.
118. Nakagawa T, Zhu H, Morishima N, Li E, Xu J, Yankner BA, et al. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-beta. Nature 2000;403:98-103.
119. Wolf BB, Green DR. Suicidal tendencies: apoptotic cell death by caspase family proteinases. J Biol Chem 1999;274:20049-20052.
120. Cryns V, Yuan J. Proteases to die for. Genes Dev 1998;12:1551-1570.
121. Namura S, Zhu J, Fink K, Endres M, Srinivasan A, Tomaselli KJ et al. Activation and cleavage of caspase-3 in apoptosis induced by experimental cerebral ischemia. J Neurosci 1998;18:3659-3668.
122. Bialik S, Cryns VL, Drincic A, Miyata S, Wollowick AL, Srinivasan A et al. The mitochondrial apoptotic pathway is activated by serum and glucose deprivation in cardiac myocytes. Circ Res 1999;85:403-414.
123. Gervais FG, Xu D, Robertson GS, Vaillancourt JP, Zhu Y, Huang J et al. Involvement of caspases in proteolytic cleavage of Alzheimer's amyloid-beta precursor protein and amyloidogenic A beta peptide formation. Cell 1999;97:395-406.
124. Sanchez I, Xu CJ, Juo P, Kakizaka A, Blenis J, Yuan J. Caspase-8 is required for cell death induced by expanded polyglutamine repeats. Neuron 1999;22:623-633.
125. Rao RV, Castro-Obregon S, Frankowski H, Schuler M, Stoka V, Rio G del, et al. Coupling endoplasmic reticulum stress to the cell death program. An Apaf-1-independent intrinsic pathway. J Biol Chem 2002;277:21836-21842.
126. Li P, Nijhawan D, Budihardjo I, Srinivasula SM, Ahmad M, Alnemri ES et al. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997;91:479-489.
127. Zou H, Li Y, Liu X, Wang X. An APAF-1.cytochrome c multimeric complex is a functional apoptosome that activates procaspase-9. J Biol Chem 1999;274:11549-11556.
128. Saleh A, Srinivasula SM, Acharya S, Fishel R, Alnemri ES. Cytochrome c and dATP-mediated oligomerization of Apaf-1 is a prerequisite for procaspase-9 activation. J Biol Chem 1999;274:17941-17945.
129. Lazebnik YA, Kaufmann SH, Desnoyers S, Poirier GG, Earnshaw WC. Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 1994;371:346-347.
130. Szatkowski M, Attwell D. Triggering and execution of neuronal death in brain ischaemia: two phases of glutamate release by different mechanisms. Trends Neurosci 1994;17:359-365.
131. Choi DW. Ischemia-induced neuronal apoptosis. Curr Opin Neurobiol 1996;6:667-672.
132. Mattson MP, Guo Q. Cell and molecular neurobiology of presenilins: a role for the endoplasmic reticulum in the pathogenesis of Alzheimer's disease? J Neurosci Res 1997;50:505-513.
133. Pascale A, Etcheberrigaray R. Calcium alterations in Alzheimer's disease: pathophysiology, models and therapeutic opportunities. Pharmacol Res 1999;39:81-88.
134. Furuya Y, Lundmo P, Short AD, Gill DL, Isaacs JT. The role of calcium, pH, and cell proliferation in the programmed (apoptotic) death of androgen-independent prostatic cancer cells induced by thapsigargin. Cancer Res 1994;54:6167-6175.
135. Iversen LL, Mortishire-Smith RJ, Pollack SJ, Shearman MS. The toxicity in vitro of beta-amyloid protein. Biochem J 1995;311 ( Pt 1):1-16.
136. Heizmann CW, Braun K. Changes in Ca(2+)-binding proteins in human neurodegenerative disorders. Trends Neurosci 1992;15:259-264.
137. Orrenius S, Nicotera P. The calcium ion and cell death. J Neural Transm Suppl 1994;43:1-11.
138. Nicotera P, Rossi AD. Nuclear Ca2+: physiological regulation and role in apoptosis. Mol Cell Biochem 1994;135:89-98.
139. Oyadomari S, Takeda K, Takiguchi M, Gotoh T, Matsumoto M, Wada I et al. Nitric oxide-induced apoptosis in pancreatic beta cells is mediated by the endoplasmic reticulum stress pathway. Proc Natl Acad Sci U S A 2001;98:10845-10850.
140. Wang XZ, Lawson B, Brewer JW, Zinszner H, Sanjay A, Mi LJ et al. Signals from the stressed endoplasmic reticulum induce C/EBP-homologous protein (CHOP/GADD153). Mol Cell Biol 1996;16:4273-4280.
Recibido: 13 de octubre de 2003. Aprobado: 23 de septiembre de 2003.
Dr. Oscar Díaz Horta. Instituto Nacional de Endocrinología. Zapata y D, Vedado, Ciudad de La Habana, Cuba.
1 Doctor en Ciencias Biológicas. Investigador Agregado.
]]>