 ]]>
]]>
ARTÍCULO ORIGINAL
Producción a nivel de laboratorio de Clostridium septicum IRP15 para la formulación de una vacuna veterinaria
Laboratory production of Clostridium septicum IRP15 for the formulation of a veterinary vaccine
María Lorena González Ramírez,I Ángela María Parra Pedrosa,I Janeth Arias Palacios,II Milton Forero LozanoII
I Universidad Javeriana. Bogotá. Colombia. ]]> II Universidad Nacional de Colombia. Bogotá, Colombia.
RESUMEN
Objetivo: establecer las condiciones de producción a nivel de laboratorio de la alfa toxina de Clostridium septicum IRP15 para la formulación de una vacuna veterinaria y la optimización del proceso de producción.
Métodos: se caracterizó y estandarizó la edad apropiada del inóculo para los cultivos en un fermentador New Brunswick Scientific 7 L. Las condiciones de cultivo fueron: cepa C. septicum IRP15, medio de cultivo VBH, 5 L/vaso de 7 L, inóculo de 250 mL (5 %), 37 ºC, 24 h, bajo agitaciones prueba de 0, 25 y 50 r.p.m. Se estableció el perfil cinético morfológico, de biomasa, consumo de sustrato y producción de toxina.
Resultados: para las fermentaciones de 0 y 25 r.p.m. no se presentó fase de adaptación; el microorganismo creció de manera exponencial hasta las 4 y 6 h de fermentación, consumiendo simultáneamente la mayor cantidad de glucosa presente en el medio. A partir de estas horas y hasta las 24, se realizó la prueba de DL50 en ratones y se destaca que a 25 r.p.m. se obtuvo el mayor título de toxina (1/23). En las fermentaciones a 50 r.p.m. se observó que el microorganismo experimenta una fase de adaptación de 4 h aproximadamente; con un retardo en producción de biomasa, consumo de glucosa y producción de toxina, condición que no resulta óptima para la producción del antígeno.
Conclusiones: la producción de toxina se presenta en la fase logarítmica y durante la fase estacionaria, asociándose así al crecimiento y al fenómeno de esporulación.
Palabras clave: fermentación, agitación, biorreactor, inóculo, cinética.
ABSTRACT
]]> Objective: to set the laboratory production conditions of Clostridium septicum IRP15 alpha toxin for the formulation of a veterinary vaccine and the optimization of the productive process.Key words: fermentation, shaking, bioreactor, inoculum, kinetics.
INTRODUCCIÓN
La presencia de casos de mortalidad en el ganado es un problema económico importante para los sistemas de producción ganadera en Colombia; generalmente indicio de diversos agentes bacterianos, virales, parasitarios o tóxicos, se han asociado como causa de estas muertes.1 Clostridium septicum es el agente causal de enfermedades como la gangrena gaseosa en vacunos y abomastitis en ovinos.1 Esta especie se caracteriza por producir exotoxinas que funcionan como entidades separadas, de acuerdo con estudios serológicos y de actividad enzimática.2 Estas incluyen la toxina a, una toxina caracterizada por ser necrotizante y letal, la DNasa o toxina b, la hialuronidasa o toxina g, la septicolisina o toxina d. La severidad de las enfermedades depende directamente de la producción de estas.
La utilización de vacunas específicas, ha conducido a adecuados niveles de control, por tanto, el objetivo del estudio fue establecer las condiciones de producción a nivel de laboratorio de la alfa toxina de C. septicum IRP15 para la formulación de una vacuna veterinaria optimización del proceso de producción.
]]>La primera etapa consistió en estandarizar las técnicas de peso seco para la determinación de biomasa y antrona para la determinación de azúcares totales, expresados como glucosa. Luego se determinó el perfil cinético de biomasa, consumo de sustrato, producción de toxina y variación del pH de C. septicum cultivado en un biorreactor New Brunswick de 7 L. Finalmente se compararon los resultados del proceso de producción de alfa toxina a diferentes condiciones de agitación (0, 25 y 50 r.p.m.).
CEPA
C. septicum IRP15 del cepario de microorganismos del laboratorio de investigación de la Empresa Farmacéutica, previamente aislada e identificada. Se conservó en criopreservación, 50 % v/v glicerol, a -70 °C y a temperatura ambiente mediante la técnica de semillero en tubo plántula con tapón de vaselina.
INÓCULO
Por fermentaciones de 12 y 24 h en 500 mL medio VBH en fermentador adaptado en frascos SCHOTT de 500 mL bajo condiciones de anaerobiosis, a 37 ºC, sin agitación y un 5 % de inóculo proveniente del semillero en tubo plántula, se realizó seguimiento del crecimiento, logrando establecer las 12 h de fermentación como la edad apropiada del inóculo para los cultivos de C. septicum en el fermentador New Brunswick Scientific. Co. Celligen Plus de 7 L.
]]> CONDICIONES DE CULTIVO
Se realizaron fermentaciones discontinuas con diferentes velocidades de agitación: 0, 25 y 50 r.p.m. y se mantuvieron constantes las siguientes condiciones: 4 750 mL medio de cultivo VBH/vaso del fermentador con capacidad para 7 L, el porcentaje de inóculo fue de 5 % (250 mL) del volumen efectivo de trabajo (5 L), temperatura de incubación 37 ºC, tiempo de incubación 24 h.
TOMA DE MUESTRAS Y ANÁLISIS
Se tomaron muestras asépticamente, a intervalos de 2 h. Se realizó tinción de Gram para observar la morfología de C. septicum y la pureza del cultivo. Se midió pH, se determinó biomasa, concentración de glucosa y título de toxina. Las muestras se dispensaron en tubos tapa rosca, tomando una alícuota para la lectura de biomasa por densidad óptica a 540 nm, se centrifugó la fracción restante a 4 000 r.p.m. durante 30 min a 4 ºC , se congeló el sobrenadante a -70 ºC hasta el momento de uso.
CURVA DE PESO SECO
La concentración de biomasa en gramo por litro (g/L) de peso seco se calculó mediante la ecuación de calibración obtenida correlacionando valores experimentales de suspensiones celulares a diferentes concentraciones:
Biomasa (g/L)= 0,2377(Abs) - 0,049
]]> PRODUCCIÓN DE TOXINA
Se cuantificó la toxina realizando prueba de dosis letal 50 (DL50) en ratones,3 en el sobrenadante (fracción en la que se encuentra la toxina), a partir de las horas en las que se observó un incremento representativo de células durante la fase exponencial, a fin de determinar si la toxina se comporta o no, como un metabolito asociado al crecimiento. La cantidad de toxina se expresa en títulos que representan la mayor dilución en la cual muere el 50 % de la población de estudio, calculada por el método de Reed-Muench
CONSUMO DE SUSTRATO
La glucosa residual se determinó siguiendo la técnica colorimétrica para azúcares reductores antrona. La densidad óptica de las muestras procesadas se leyó a 622 nm y la glucosa en gramo por litro (g/L) se determina según la ecuación:
Glucosa g/L= Am x Cp / Ap x Fd
donde:
Am: absorbancia de la muestra
Cp: concentración del patrón
Ap: absorbancia del patrón ]]>
Fd: factor de dilución expresado como (1/n).
En el procesamiento estadístico se aplicó un análisis de varianza (ANOVA) para probar si existen diferencias en la producción de biomasa, de toxina y en el consumo de glucosa cuando se tienen diferentes condiciones de agitación.
RESULTADOS
CINÉTICAS DE CRECIMIENTO
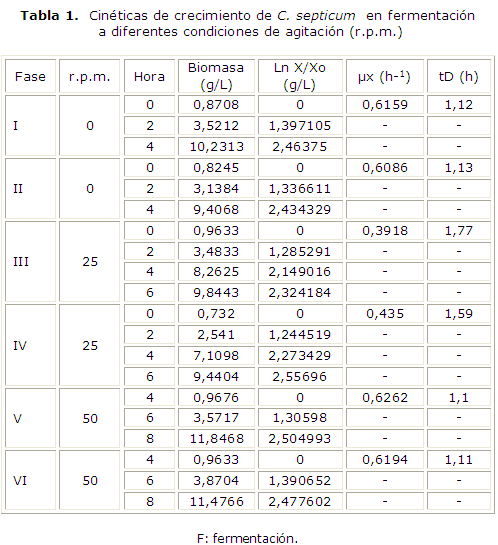
Fue posible verificar el curso del crecimiento de C. septicum en fermentación por 24 h, e identificar las etapas de adaptación, exponencial y estacionaria. La fase de adaptación se presentó durante las cuatro primeras horas de fermentación con valores de biomasa (g/L) comprendidos entre 0,5048 y 0,6058 (tabla 1).
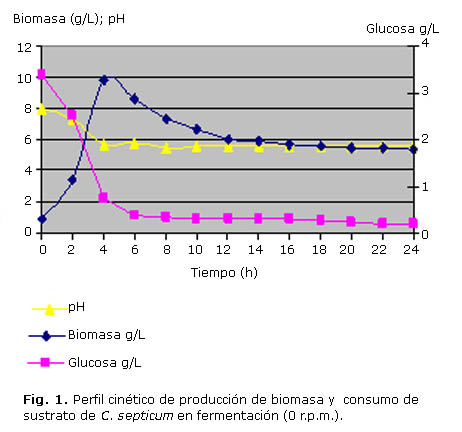
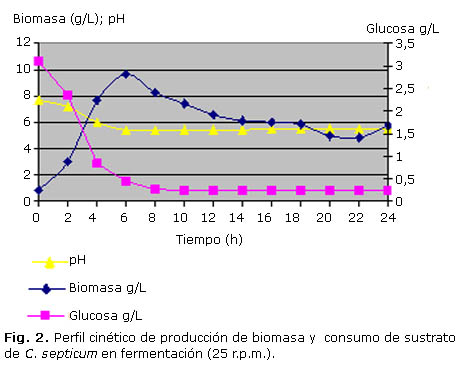
En las figuras 1, 2 y 3 se puede apreciar la evolución de la biomasa, de la glucosa y el curso del pH durante las fermentaciones de 0 r.p.m., 25 r.p.m. y 50 r.p.m. realizadas en el fermentador New Brunswick de 7 L.
Durante el cultivo de la cepa se realizó seguimiento de la glucosa, la producción de biomasa, el pH, desde la hora 0 hasta las 24 h de cultivo a tres diferentes revoluciones de agitación, la tabla 2 muestra los resultados.
]]>En la figura 4 se muestran los valores de las DL50 obtenidas para las diferentes horas de fermentación evaluadas.
DISCUSIÓN
Al revisar las figuras 1 y 2, se evidencia que no hubo una fase de adaptación del inoculo, sino que el microorganismo continuó su crecimiento de manera exponencial hasta la hora 4 y 6 de fermentación respectivamente. Posteriormente se presentó una disminución del crecimiento hasta alcanzar una fase estacionaria que se prolongó hasta la hora 24 en ambos casos, aunque para la curva de 25 r.p.m se observó un mínimo ascenso en el valor de biomasa a la hora 24, el cual puede obedecer probablemente al crecimiento de algunas células a expensas de los restos celulares dejados por la lisis de otras células.
Por otra parte, la glucosa, fuente energética contenida en el medio VBH, fue degradada rápidamente por el microorganismo de manera similar en las fermentaciones de 0 r.p.m. y 25 r.p.m., soportando principalmente el periodo de crecimiento (Fig. 2 y 3), ya que posteriormente se hallaron valores mínimos de glucosa presente en el medio (tabla 2).
De igual manera, en esta etapa del crecimiento posiblemente se consume la mayor parte de las fuentes proteicas presentes en el sustrato (extracto de cerebro, corazón, peptonas y aminoácidos) y como producto del metabolismo de los azúcares y las fuentes proteicas se producen gases como CO2 y ácidos débiles como el ácido acético y el ácido butírico.4
El comportamiento del pH presenta una disminución gradual que está posiblemente relacionada con el aumento de la biomasa y la producción de ácidos en el cultivo durante la fase de crecimiento. Luego se presenta un aumento que aunque es mínimo, se hace progresivo ya que hay liberación de material proteico y metabolitos nitrogenados, producto de la lisis de algunas células lo cual alcaliniza el medio.
A diferencia de lo ocurrido en las fermentaciones de 0 y 25 r.p.m., en las fermentaciones de 50 r.p.m. se evidenció que el microorganismo si experimentó la fase de adaptación por aproximadamente 4 h, como se puede observar en la figura 4. Dado este hecho, se presume que la agitación podría llegar a retardar el crecimiento del microorganismo. Hacia la 6 y 8 h, el microorganismo inicia su fase exponencial experimentando un crecimiento acelerado, para luego sufrir un descenso y entrar en la fase estacionaria.
]]> En cuanto al comportamiento de la glucosa, se evidenció la influencia de la fase de adaptación en el consumo lento del azúcar durante las cuatro primeras horas de fermentación y posteriormente en la fase de crecimiento se presentó el consumo de la mayor parte de la glucosa del medio (tabla 1). El comportamiento del pH sufre un cambio significativo hacia la hora 6 y 8 (tabla 2) que, sumado al comportamiento de la glucosa refleja los cambios metabólicos descritos en las fermentaciones.Basándose en las correlaciones del análisis de varianza con un valor p> 0,05 (0,899) se encontró que no existe diferencia significativa en la producción de biomasa ni en el consumo de glucosa, por parte del microorganismo cuando se tienen condiciones de agitación de 0, 25 y 50 r.p.m. La evolución de la concentración celular en las horas correspondientes a la fase exponencial4-8 de los cultivos a 50 r.p.m. resulta bastante rápida, presentando valores máximos de biomasa de 11,8468 g/L y 11,4766 g/L, dados bajo una velocidad específica de crecimiento (µx) de 0,6262 h-1 y 0,6194 h-1 y de tiempos de duplicación (tD) de 1,10 h y 1,11 h respectivamente, haciendo de esta la fermentación con mayor µx y menor tD a pesar de haber presentado un retardo en el inicio de la fase exponencial (tabla 2).
En los cultivos a 0 r.p.m. se obtuvieron valores máximos de biomasa de 10,2313 g/L y 9,8443 g/L a la altura de las 4 h, los cuales fueron alcanzados con tiempos de duplicación de 1,12 h y 1,13 h y velocidades específicas de crecimiento de 0,6159 h-1 y 0,6086 h-1 que, al ser comparados con los de los cultivos a 50 r.p.m. expresan una menor velocidad de crecimiento y por tanto un mayor tiempo para la duplicación de la población.
Finalmente siendo los valores calculados de µx de 0,3918 y 0,435 y de tD de 1,77 y 1,59 respectivamente, para los cultivos a 25 r.p.m. los que representan una velocidad de crecimiento menor y, por tanto tiempos de duplicación mayores, se puede decir que en estas fermentaciones el crecimiento del microorganismo fue más lento con respecto a las realizadas a 0 y 50 r.p.m.
Con relación a las DL50 obtenidas permiten determinar la presencia de la alfa toxina en todas las horas evaluadas entre las que se incluyen puntos dentro de la fase exponencial, se pudo establecer que la toxina es un metabolito ligado al crecimiento. Se encontró que existe similitud en el comportamiento de la producción de la toxina en las fermentaciones de 0 y 25 r.p.m., a pesar de que en algunas horas de la fase estacionaria se presentan unos valores más altos que otros permitiendo señalar que dicha producción no se va incrementando conforme avanza el tiempo de cultivo. El análisis estadístico permitió establecer que no existe diferencia estadísticamente significativa en la producción de la toxina bajo las condiciones de 0 y 25 r.p.m., contrario a lo que ocurrió con la producción de toxina en fermentación a 50 r.p.m., en que los títulos obtenidos son inferiores.
Las deducciones a partir de los resultados de la producción de la toxina de C. septicum resultan coherentes con lo sugerido por diversos autores entre los que se encuentran,5 quien estableció que la producción de toxina en cultivos de C. septicum se incrementa a medida que aumenta la población bacteriana, alcanzando su máximo valor cuando la concentración celular llega a su punto máximo, o inmediatamente después en el inicio de la fase estacionaria; la producción de la alfa toxina por parte de este microorganismo corresponde con la fase logarítmica, en el proceso de transición entre dicha fase y la fase estacionaria.6
Es importante señalar que debido a que las fermentaciones realizadas a 0 y 25 r.p.m. mostraron resultados representativos, se podría optar por realizar la producción del antígeno en un sistema sin agitación. Sin embargo, es importante destacar que a 25 r.p.m. se obtuvo el mayor título (1/23) de todas las fermentaciones y además, al plantear la producción en cultivos a gran escala no se debe dejar de lado que los volúmenes y escalas en laboratorio son diferentes a los de producción. Tampoco descartar el efecto positivo que puede generar la agitación en cuanto a que se puede obtener un fluido más uniforme eliminando gradientes de concentración y temperatura, así como favorecer la salida de los gases del cultivo de manera eficiente, la combinación de los componentes solubles del medio como los azúcares y el mantenimiento de la suspensión de las células evitando la formación de zonas sin nutrientes en aquellos puntos de mayor consumo por parte de las células.7
Se puede concluir que parámetros cinéticos como la velocidad específica de crecimiento y el tiempo de duplicación, permitieron tener un mayor conocimiento del comportamiento de microorganismo durante el desarrollo de los cultivos a través de la fermentación y correlacionar este comportamiento con el consumo de glucosa.
El perfil cinético observado a 50 r.p.m. evidenció que bajo estas condiciones se retarda la producción de biomasa, razón por la cual no se considera una condición óptima para este proceso.
Las técnicas de peso seco y antrona, resultan apropiadas para conocer el desarrollo de la biomasa y el consumo de azucares, además de determinar el efecto que tienen las condiciones de operación o cambios que se puedan generar durante el proceso de cultivo del C. septicum.
]]>1. Hateway C. Toxigenic Clostridia. Clin Microbiol Rev. 1990;3(1):66-98.
2. Ballard J, Bryant A, Stevens D, Tweten RK. Purification and characterization of the lethal toxin (Alpha Toxin) of Clostridium septicum. Infection and Immunity; 1992;60:784-90.
3. Kaplan MM, Koprowsky H, Meslin FX. Laboratory Techniques in Rabies. Geneva: World Health Organization; 1996.
4. Bergey's manual of systematic bacteriology. Clostridum septicum. Baltimore: Williams and Wilkins; 1984. p. 1197-8.
]]>5. Bernheimer AW. Paralelism in the lethal and hemolytic activity of the toxin of Clostridium septicum. J Exp Med. 1944;80:309-30.
6. Ballard J, Crabtree J, Roe B, Tweten R. The primary structure of Clostridium septicum Alpha toxin Exhibits similarity with that of Aeromonas hydroprhila. Aerol Infect Immun. 1995;63(1):340-4.
7. Doran P. Principios de Ingeniería de los Bioprocesos. Zaragoza: Editorial Acribia SA; 1998. p. 278-95.
Recibido: 4 de noviembre de 2013.
Aprobado: 20 de diciembre de 2013.
María Lorena González Ramírez. Universidad Javeriana. Carretera 7a No. 43-82. Bogotá, Colombia. Correo electrónico: jdcarias@javeriana.edu.co
]]>
{kind=link}
{kind=link}