Diversidad genética del gato (Felis catus) en San Bernardo del Viento, Colombia, mediante marcadores morfológicos
Genetic diversity of the cat (Felis catus) through morphological markers in San Bernardo del Viento, Colombia
Enrique Pardo-Pérez1*, Teodora Inés Cavadía-Martínez1, José Luis Barrera-Violeth2
1 Departamento de Biología, Facultad de Ciencias Básicas, Universidad de Córdoba, Montería, Colombia.
2 Departamento de Ingeniería Agronómica y Desarrollo Rural. Facultad de Ciencias Agrarias, Universidad de Córdoba, Montería, Colombia.
]]>
RESUMEN
Los marcadores fenotípicos (genes de pelaje) son una herramienta para determinar la diversidad de las poblaciones de gatos. El objetivo de este estudio fue evaluar la diversidad y la estructura genética en poblaciones de gato (Felis catus) en San Bernardo del Viento, Córdoba, Colombia. Se realizaron muestreos aleatorios y, mediante observaciones directas y se caracterizaron fenotípicamente 213 gatos, entre los meses de septiembre y noviembre de 2015, en recorridos por las zonas urbanas de San Bernardo del Viento, en concordancia con la nomenclatura recomendada por Committe Standardized Genetic Nomenclature For Cats, atendiendo a los marcadores autosómicos de codificación morfológica: el locus ligado al sexo Orange (O) y los loci autosómicos Non-agouti (a), Tabby blotched (Tb), No diluido (d), Pelo largo (l) Manchado de blanco (s) y Dominante blanco (W). Se calcularon los siguientes parámetros genéticos: frecuencia alélica, diversidad genética, flujo génico, estructura genética, equilibrio de Hardy-Weinberg y distancia genética y se infirieron las relaciones filogenéticas entre las poblaciones de gatos. El marcador Non-agouti fue el de mayor frecuencia, mientras los genes Tabby blotched y Pelo largo presentaron los menores valores. Se encontró escasa diversidad genética entre las poblaciones (DST) y diversidad dentro de las poblaciones (HS), al igual que un elevado flujo génico; se observó un exceso de heterocigotos a nivel poblacional y la ausencia de equilibrio Hardy-Weinberg. Las comunidades de gatos estudiadas se hallan conectadas genéticamente.
Palabras clave: diversidad, heterocigosidad, Felis catus, marcadores fenotípicos, equilibrio Hardy-Weinberg.
ABSTRACT
The phenotypic markers (coat genes) are a tool to determine the diversity of cat populations. The objective of this study was to evaluate the diversity and genetic structure of the cat (Felis catus) populations, in San Bernardo del Viento, Córdoba, Colombia. Samples were randomized and, through direct observations, 213 cats were phenotypically characterized taking into account the following aspects: September-November period, on routes through the urban areas of San Bernardo del Viento, and in accordance with the nomenclature recommended by the Committee Standardized Genetic Nomenclature For Cats. Autosomal markers of morphological coding like the sex-linked locus Orange (O) and the autosomal loci Non-agouti (a), Tabby blotched (Tb), Dilution (d), Long hair (l), Spotted white (S), and Dominant white (W), were used. The following genetic parameters were calculated: allele frequency, genetic diversity, gene flow, Hardy-Weinberg equilibrium, and genetic distance. The phylogenetic relationships among cat populations were inferred. The Non-agouti marker was the most frequent while Tabby blotched and Long hair genes had the lowest values. There was a low genetic diversity among populations (DST) and a great diversity within populations (HS), as well as a high gene flow. An excess of heterozygotes at the population level and the absence of Hardy-Weinberg equilibrium were observed. The communities of cats studied are genetically connected.
Keywords: diversity, heterozygosity, Felis catus, phenotypic markers, Hardy-Weinberg equilibrium.
]]> INTRODUCCIÓN
El gato doméstico (Felis catus) es un mamífero pequeño, carnívoro, perteneciente a la familia de los félidos, con un peso corporal de casi 5 kg, pero con gran variación entre razas, pues los machos suelen ser de mayor tamaño que las hembras (1).
La gran variedad de colores de la capa en los gatos domésticos (Felis catus) es la consecuencia de un complejo proceso de desarrollo y funcionamiento de estructuras implicadas en la formación del pigmento (2). Este proceso, conocido como pigmento génesis, está regulado por una serie de genes, los cuales normalmente conducen al desarrollo del llamado fenotipo silvestre (atigrado). Todas las otras variaciones del color de la capa del gato son causadas por mutaciones de los genes correspondientes, características posibles de identificar a simple vista, motivo por el cual la recopilación de datos es un proceso sencillo. Asimismo, los gatos son una especie ideal para este tipo de análisis poblacionales por ser animales cosmopolitas y ser una población con apareamientos aleatorios (3).
Las frecuencias de los alelos mutantes y silvestres de los genes de color de pelaje son específicas para cada población y deben reflejar su estructura genética (4).
En la actualidad, las frecuencias alélicas de los genes del pelaje se han descrito para los gatos domésticos de casi todos los territorios, incluidos poblaciones americanas (5,6,7), europeas (8,9) y asiáticas (10).
El objetivo de este estudio fue determinar la diversidad genética y el nivel de diferenciación de Felis catus en las poblaciones de las áreas urbanas de San Bernardo del Viento.
MATERIALES Y MÉTODOS
Área de estudio
La recolección de datos se realizó en barrios de zonas urbanas del municipio San Bernardo del Viento, Córdoba, Colombia, ubicados a 09º 19' 48″ latitud Norte, 75º 58' 10″ longitud Oeste y una altura de 18 m sobre el nivel del mar.
]]> Recolección de datosSe realizaron muestreos mediante excursiones urbanas y observación directa en los barrios de San Bernardo del Viento: Guayabal, San Felipe, Paraíso, El Centro, San José y Villa Galilea, donde se efectuó una categorización fenotípica de cada uno de los individuos adultos encontrados (n=213). Cada ruta se utilizó solo una vez, a fin de evitar el remuestreo, atendiendo a la presencia o no del locus ligado al sexo Orange (O) y los loci autosómicos Non-agouti (a), Blotched tabby (Tb), Dilution (d)Pelo largo (l) Manchado de blanco (S) y Dominante blanco (W) (Tabla 1). Además, se compararon los resultados con estudios realizados en otras ciudades del Caribe colombiano.
Variables de estudio
Para el estudio del perfil genético de las poblaciones de gatos domésticos (Felis catus) se tuvieron en cuenta marcadores fenotípicos (Tabla 1) propuestos por el Committee on Standardized Genetic Nomenclature for Cats (12): O (Orange: Carácter ligado al sexo) y los loci autosómicos, A (A, a; agouti vs. non-agouti), T (Ta, t+, tb; Abyssinian tabby vs. mackerel vs. Blotched tabby), D (D, d; full color vs. Dilution, L (L, l; pelo corto vs. pelo largo), S (s, S; no manchado de blanco vs. manchado de blanco) y W (w, W; color normal vs. Dominante blanco).
Análisis poblacional
El cálculo de las frecuencias alélicas, la diversidad genética, el flujo génico, el equilibrio Hardy-Weinberg y la distancia genética se estimaron a través del programa Pop Gene 1.31 (13); la estructura genética mediante el programa FSTAT y los dendogramas con el programa MEGA 5.2 (14).
RESULTADOS
Frecuencias alélicas
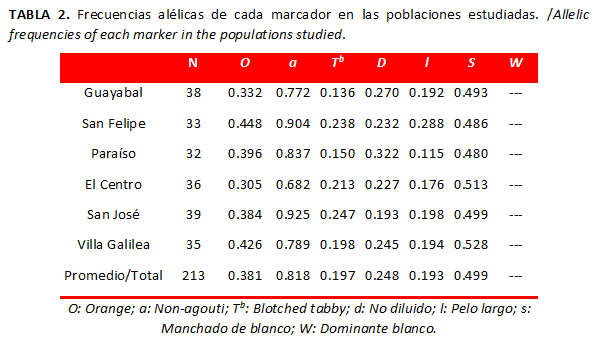
Se observaron 213 especímenes. Al determinar las frecuencias alélicas para cada población (Tabla 2) se encontró que el fenotipo Non-agouti fue el más frecuente, principalmente en las poblaciones de San José (q=0. 0.925) y San Felipe (q=0. 0.904). El gen Dominante blanco, responsable de la capa blanca, no se registró en las poblaciones y este fue el marcador menos frecuente en la población de San Bernardo del Viento. Asimismo, los marcadores pelo largo y Blotched tabby mostraron frecuencias bajas, tanto a nivel poblacional como a nivel global (Tabla 2). Es importante señalar que la frecuencia del gen Manchado de blanco fue alta (p=0.499), si se compara con las reportadas en otras poblaciones.
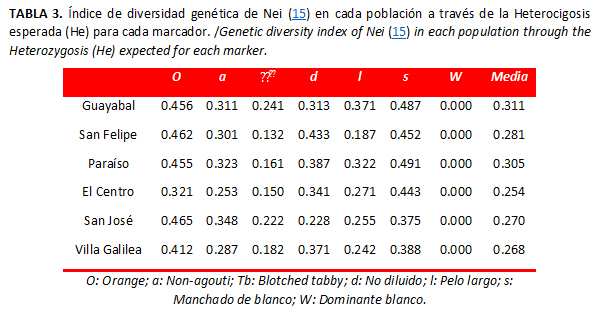
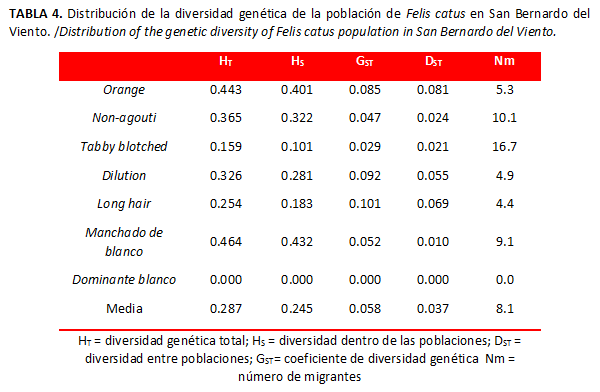
]]> Diferenciación y flujo genéticoPara cada uno de los genes estudiados en las poblaciones observadas, el nivel medio de heterocigosidad esperado fue 0.281 (Tabla 3), la heterocigosidad total (Tabla 4) fue baja (HT=0.287) al igual que la diferenciación genética entre las poblaciones (DST=0,037). El coeficiente de diferenciación genética en San Bernardo del Viento fue bajo, pues mostró que alrededor del 5,8 % de la variación detectada se debe a divergencias entre las poblaciones analizadas, lo cual revela que las poblaciones no fueron significativamente diferentes para los genes estudiados e indica que todas se comportan como una sola población. De la misma forma, el alto valor de flujo génico confirma que las poblaciones mantienen un elevado intercambio genético.
Estructura poblacional
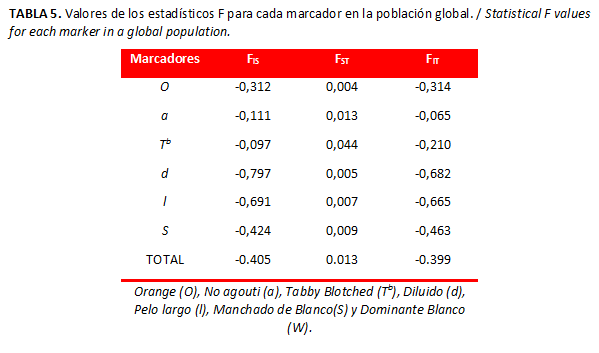
Los valores negativos obtenidos para los estadístico FIS y FIT en todos los genes (Tabla 5) señalan un exceso de heterocigotos de los individuos en cada población y de los individuos respecto a la población total. Por otro lado, el valor promedio de FST resultó ser bajo (0,013), lo cual indica exigua diferenciación genética entre las poblaciones.
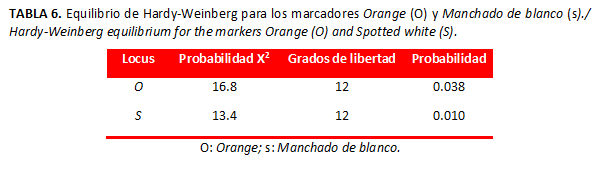
Equilibrio Hardy-Weinberg
En cuanto al test de equilibrio Hardy-Weinberg (Tabla 6), a nivel global la población de San Bernardo del Viento reveló falta de equilibrio para los marcadores Manchado de Blanco y Orange.
Distancia Genética
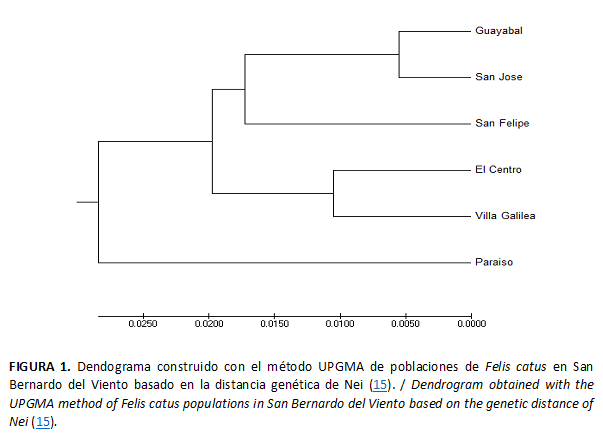
La distancia genética entre las poblaciones de San Bernardo del Viento reveló a Guayabal y San José como las poblaciones más cercanas (Tabla 7), mientras que El Centro y Paraíso mostraron ser las de mayor divergencia genética, pero con valores poco significativos, pues estos no superan el 9 %.
El dendograma UPGMA realizado a partir de la distancia genética de Nei (15) para las seis poblaciones de San Bernardo del Viento (Fig. 1) enfatiza la semejanza genética entre las poblaciones de Guayabal, San José y San Felipe; igualmente la población Paraíso es la más alejada, con un valor de distancia poco significativo, lo que permite deducir que las poblaciones en conjunto están estrechamente relacionadas.
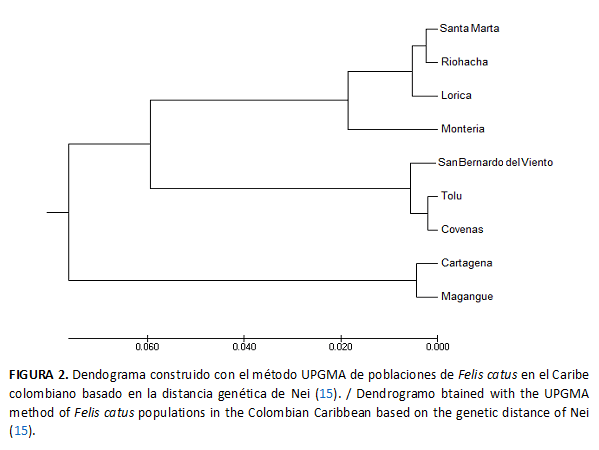
Asimismo, el dendograma UPGMA obtenido a partir de la distancia genética de Nei (15) para las poblaciones del Caribe colombiano (Fig. 2) muestra a Tolú, Coveñas y San Bernardo del Viento como las poblaciones más cercanas, mientras Cartagena y Magangué se comportaron como las más alejadas.
]]>DISCUSIÓN
La ausencia del fenotipo Dominante blanco es un resultado semejante a lo alcanzado en investigaciones realizadas en otras poblaciones colombianas (16), lo cual puede mostrar el elevado grado de intervención humana en las poblaciones de gatos. Sin embargo, la ausencia del fenotipo Dominante blanco puede responder a efectos pleiotrópicos sobre la audición (17), que los coloca en desventaja selectiva frente a los no portadores del gen; esto puede ocasionarle la muerte a una edad temprana y disminuir así la cantidad de animales portadores.
De igual manera, la mayor frecuencia del fenotipo Non-agouti podría relacionarse con los beneficios que dicho fenotipo brinda a los individuos que lo portan en ambientes urbanos, donde las densidades poblacionales son altas (18), pues tienden a ser más sociables con otros congéneres y así poder adaptarse a una mayor intrusión humana en sus vidas; lo anterior permite suponer que los gatos portadores de este gen están mejor acondicionados a las situaciones preponderantes de estos lugares que los que no lo llevan. Igualmente, es posible que las elevadas temperaturas acaecidas durante la mayor parte del año en San Bernardo del Viento podrían estar favoreciendo la presencia y el aumento de individuos portadores de dicho fenotipo Non-agouti, que los hace más resistentes a las altas temperaturas (19); asimismo, las predilecciones antrópicas evidencian una posible selección artificial, de tal manera que sus frecuencias se elevaron progresivamente (20).
La ausencia del equilibrio de Hardy-Weinberg para los alelos O (Orange) y S (Manchado de blanco) pudo deberse a diferentes causas; como los resultados revelan un déficit de homocigotos, podría atribuirse, en este caso, a algunos factores evolutivos como la selección artificial, pues los humanos tienen predilección por cierto tipo de carácter en los gatos, lo que ha favorecido algunas características fenotípicas más que otras. Otra razón para explicar esta desviación es la proximidad geográfica entre las poblaciones analizadas, lo que produce un incremento importante del flujo génico existente entre estas y, por tanto, provoca en la población un descenso de genotipos homocigotos (21).
La diversidad genética total (HT) encontrada en San Bernardo del Viento fue moderada; la mayor parte de la diversidad génica se encuentra dentro de las poblaciones (Hs) y poca entre ellas (DST), lo que indica que las poblaciones locales comparten una gran proporción de la diversidad total. El flujo de genes (Nm) resultó alto; en términos generales se considera que valores de Nm cercanos o mayores de cuatro se consideran suficientes para conservar una relativa homogeneidad del acervo génico (22). Estos resultados demuestran que supuestamente no hay diferencia entre las poblaciones observadas. La no diferenciación genética entre poblaciones es regularmente considerada como la consecuencia de un suficiente flujo de genes, tradicionalmente dado por migraciones, acaecido en todas las poblaciones para contrarrestar los efectos de la deriva genética o la selección, ya que cuando Nm es mayor a 1 se espera que las poblaciones conserven conectividad genética, dado que el flujo genético sobrepasa los resultados de la deriva e imposibilita la distinción local (23).
El alto nivel de flujo génico permite deducir que las poblaciones se encuentran estrechamente relacionadas genéticamente y se comportan como una metapoblación (24).
En el dendograma elaborado a partir de los valores de distancia genética de Nei (22) para las seis poblaciones de San Bernardo del Viento se aprecia que la cercanía génica responde a la vecindad geográfica y a la antigüedad de los barrios, donde se presume que arribaron los primeros gatos a la población (25).
En el dendograma obtenido a partir de los valores de la distancia genética de Nei (22) para las ciudades del Caribe colombiano se encontró que las ciudades como Santa Marta, Riohacha, Lorica y Montería se unieron en un grupo y, muy cercanas a ellas, se concentraron Coveñas, Tolú y San Bernardo del Viento, mientras que Cartagena y Magangué fueron las más alejadas. Este gran parecido genético entre las poblaciones puede relacionarse con un efecto fundador, ya que se originaron a partir de las poblaciones españolas (24) por ser esta una especie inexistente en América hasta la llegada de los europeos, y su ingreso debió estar coligado a las rutas colonizadoras de diferentes naciones europeas en el Nuevo Mundo (16). Adicional a esto se debe resaltar el parecido en las condiciones medio ambientales del Caribe, tales como la temperatura, la humedad relativa, entre otras.
Podemos concluir que el marcador con la mayor frecuencia fue Non-agouti, lo cual podría estar asociado con factores como las elevadas temperaturas y las preferencias antrópicas. Se encontró, además, ausencia del equilibrio de Hardy-Weinberg para los alelos O (Orange) y S (Manchado de blanco). La diversidad genética total fue moderada: la mayor parte de la diversidad génica se encontró dentro de las poblaciones y poca entre ellas, lo cual evidencia que las poblaciones locales comparten una gran proporción de la diversidad total. El dendrograma mostró que San Bernardo del Viento se comportó como una metapoblación y, al contrastarla con otras ciudades del Caribe colombiano, se observó un gran parecido entre ellas que puede corresponder a un evento fundador común.
]]>
Recibido: 6/9/2016
Aceptado: 22/4/2017
*Autor para correspondencia: Enrique Pardo-Pérez. E-mail: epardop@correo.unicordoba.edu.co
]]>{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}