]]> Datos de fósiles y moleculares han demostrado que los HMA son muy antiguos, datan de alrededor de 350 a 400 millones de años (13, 14).
Existen varias hipótesis que defienden distintas teorías acerca del origen del hongo. Pirozynski y Malloch (15) sugirieron que fuese un alga semiacuática. Por otra parte, Morton (16) no aceptó este ancestro y sugirió un hongo zygomicetes saprobiótico con hábitat más terrestre. Recientemente la hipótesis de Morton ha sido corroborada, hoy se conoce que es un hongo zygomicetes llamado Geosiphon pyriformis (Kuts & Wettstein) (16), que no se asocia con plantas superiores pero tiene gran similitud morfológica y molecular con la línea Glomus en Glomales.
Otros trabajos demostraron que en Geosiphon pyriformis existen secuencias de nucleótidos (18S ARNr), que tienen gran similitud con hongos pertenecientes al orden Glomales, que con hongos fuera de este orden (17) y sus esporas guardan cierta semejanza con especies típicas del género Glomus (18).
Debido a la actividad fisiológica del hongo, se conoce que es capaz de vivir en autotrofía de carbono y nitrógeno (17); quizás la asimilación de carbono por el micobionte en la fotosíntesis esté estrechamente ligada con indicadores de parentesco con hongos del orden Glomales, lo cual sugiere que ha existido una estrecha y obligada relación con procariontes autotróficos.
De gran relevancia para la presente investigación fue conocer que en Geosiphon pyriformis se hubiesen detectado microorganismos asociados; se plantea la observación de especies filamentosas de Nostoc (usualmente N. puntiforme) (19). Sin embargo, Geosiphon junto al endosimbionte han sido considerados como un raro sistema endocitobiótico primitivo, porque los procariontes encontrados en su interior pueden ser experimentalmente separados y unificados (20), poniéndose en evidencia no solo el origen de estos HMA, sino también que en el hongo reconocido como más antiguo se ha detectado la presencia de microorganismos endógenos.
Relación entre los HMA y otros microorganismos
En adición a la bien conocida interacción entre plantas y hongos, las micorrizas ofrecen un excelente nicho ecológico para otros microorganismos (21). Los hongos frecuentemente hospedan microorganismos del suelo, los cuales pueden influir de manera beneficiosa en la fisiología y el desarrollo de ellos (22, 23, 24). Los microorganismos pueden ser derivados de la tierra circundante o ser transportados por esporas de hongos. Desde el punto de vista ecológico, muchos estudios muestran que numerosos microorganismos del suelo interactúan con los hongos, promoviendo actividad antagónica, sinérgica o de competencia (25).
]]> 1. Interacción de los HMA y las bacterias rizosféricas
Artursson y Jansson (26) confirmaron la importancia de las bacterias asociadas a las micorrizas, las que años anteriores Garbaye (27) había identificado como bacterias que ayudan a la micorrización y las denominó mycorrhization helper bacteria (MHB) y propuso cinco mecanismos que explican el comportamiento de las MHB:
cambios beneficiosos en el suelo rizosférico
mejoramiento del reconocimiento entre la raíz y el hongo
incremento de la susceptibilidad de las plantas a la colonización micorrízica
mejoramiento del estado nutricional del hongo
estimulación de la germinación de los propágulos del hongo.
Estas poblaciones microbianas tienen un efecto beneficioso en el crecimiento del HMA, no solo porque mejoran la colonización radical y estimulan el crecimiento de la hifa extrarradical, sino porque facilitan la germinación de la esporas del HMA (28). Este efecto ha sido visualizado con Actinomycetes, Pseudomonas Corynebacterium y Bacillus (29) spp.
Después de varias investigaciones, Garbaye y Bowen (30) señalaron que existían verdaderas bacterias que ayudaban al hongo a adaptarse a vivir en simbiosis, localizándose en la vecindad del hongo en el suelo; con mucha frecuencia observaron Pseudomonas fluorescens, que en un 80 % traían consigo un efecto positivo sobre la formación de las micorrizas y en un 20 % exhibían un halo inhibitorio, revelando entonces que en la relación bacterias y su efecto sobre la formación de las micorrizas, no se debe ser totalmente absoluto. ]]> También fueron encontradas bacterias formando parte de la simbiosis entre el hongo y la planta, en diversas condiciones del sitio a muestrear y en diferentes combinaciones hongo-planta.
Estas bacterias rizosféricas en gran medida son las responsables de los cambios beneficiosos en la rizosfera, como por ejemplo la transformación de las propiedades físico-químicas del suelo, con el objetivo de facilitar la colonización micorrízica.
2. Antecedentes de bacterias asociadas a HMA
Como es ya conocido, existe una estrecha relación entre los hongos HMA y las plantas; ahora se abre una nueva posibilidad con el estudio de bacterias y otros microorganismos que viven asociados con el hongo, y constituye una de las nuevas promesas para entender la compleja simbiosis.
Además de la conocida pregunta de si los hongos emiten señales que sean reconocidas por receptores potenciales en la planta, hoy se abre una nueva incógnita:
l ¿La extensa cantidad de bacterias en el interior o específicamente asociadas con el hongo sugiere que muchas de las simbiosis entre este y la planta sean asociaciones tripartitas?
Existen numerosas evidencias que demuestran la relación existente entre los HMA y otros microorganismos; en algunas ocasiones la literatura refiere términos como: microorganismos endógenos, bacterias que ayudan a la micorrización, endófitos y en otras ocasiones se hace referencia a microorganismos asociados.
l ¿Qué evidencias existen de microorganismos asociados a las esporas de HMA? ]]> En 1970, Mosse (31) planteó algunas hipótesis sobre la posible presencia de organismos parecidos a bacterias en el citoplasma de Endogonaceae, actual Orden Glomales. Este autor observó que las bacterias eran capaces de suplir con productos metabólicos a las esporas de los glomales en períodos de rápida multiplicación. No obstante, citó que otros habían demostrado que los microorganismos podían estimular, inhibir o matar las esporas en procesos de germinación.
Mac Donald y Chandler (32) informaron la presencia de organismos parecidos a bacterias embebidos en las paredes de las esporas de HMA Glomus caledonium (Nicol. & Gerd., Trappe & Gerd); estas bacterias localizadas en las paredes se protegen físicamente de soluciones esterilizantes y afirmaron que el crecimiento eventual de bacterias embebidas puede ser la causa de contaminaciones aparentes y espontáneas de las esporas de HMA en cultivos axénicos.
A partir de muestras de suelos rizosféricos de caña de azúcar fueron obtenidas esporas vivas y muertas de la familia Endogonaceae, principalmente de Glomus sp. y Acaulospora sp. y observaron bacterias que crecían en medio LGI. Al realizar pruebas bioquímicas (catalasa, oxidasa, producción de H2S, utilización de azúcares, hidrólisis de almidón y gelatina, tinción de Gram) a los aislados, encontraron que los siete crecían en medio LGI, que por sus características concluyeron que eran cepas de A. diazotrophicus y que las esporas podían constituir un reservorio natural de esta bacteria fijadora de nitrógeno.
En el mencionado trabajo la desinfección solo se realizó con estreptomicina (no se especifica la concentración) y agua destilada, sin que se haya comprobado que hubo una verdadera desinfección externa de las esporas. Este trabajo tiene el mérito de haber sido pionero en la idea de bacterias asociadas a las esporas de estos hongos en Cuba, pero las evidencias presentadas no constituyen una verdadera prueba de que las cepas aisladas fueran Acetobacter diazotrophicus y mucho menos que los resultados probaran la presencia de bacterias provenientes del interior de las esporas de hongos micorrizógenos (33).
No se explica cómo analizaron que los microorganismos podían haber estado albergados en el interior de las esporas. No se considera que se hizo una buena desinfección de ellas, porque no se dan datos de la concentración del antibiótico usado y no se menciona el uso de agentes clorados, desinfectantes biológicos, reactivos estos empleados para un trabajo de desinfección total de esporas de HMA.
Para una identificación de los aislados y concluir que son cepas de A. diazotrophicus, es necesario en primer lugar realizar pruebas moleculares, por ejemplo 16 SARN y secuenciar el fragmento para compararlo con el Blast (una lista de microorganismos y su fragmento conservado secuenciado, que sirve de comparación), prueba de identificación API e incluso el perfil de los ácidos grasos. Las pruebas bioquímicas realizadas dan la probabilidad de que las cepas obtenidas sean Acetobacter diazotrophicus, pero no constituye un resultado concluyente.
Años más tarde, otros observaron al microscopio electrónico de transmisión en esporas de Glomus clarum NT4 (Nicol. & Schenck) estructuras semejantes a bacterias; aun cuando no confirmaron que estas fueran verdaderas bacterias, atribuyeron la presencia de tales estructuras a posibles fallos en la descontaminación de las esporas (34).
De forma semejante, Bianciotto y colaboradores con el uso de técnicas morfológicas y moleculares demostraron que el citoplasma de HMA Gigaspora margarita (Becker & Hall) sirve de albergue a verdaderas bacterias, las cuales fueron detectadas ultraestructuralmente en esporas y micelios simbióticos. Y con el uso de microscopía confocal fluorescente, estimaron aproximadamente 250 000 organismos parecidos a bacterias en el interior de esporas de Gigaspora margarita. Utilizando microscopía electrónica, otros han detectado organismos semejantes a bacterias en HMA Glomus caledonium y Acaulospora laevis (Gerdemann & Trappe) (13).
En el HMA Gigaspora margarita se realizó una comparación entre los genes representativos de eubacterias y la secuencia de los genes de la subunidad menor de ARNr de bacterias endosimbióticas, concluyendo que los genes del ARNr de las endosimbióticas pertenecían al género Burkholderia. Los análisis filogenéticos hechos al ADNr 16S demostraron la presencia de esta bacteria en todas las fases del ciclo de vida del HMA Gigaspora margarita (14).
Numerosos estudios aportaron ideas e hipótesis suficientes, a través de las cuales varios científicos (13, 14, 21, 35) llegaron a la conclusión que los sistemas micorrízicos debían incluir otros microorganismos, además de las raíces y el hongo que era lo estudiado. ]]> Según Bianciotto y colaboradores (13, 14), la identificación taxonómica ha estado muy limitada, por lo difícil e infructuoso que ha constituido el aislamiento y crecimiento de bacterias endosimbiontes de esporas de HMA en medios de cultivo axénicos. Solamente se menciona el género Burkholderia (13) y un nuevo taxón bacteriano ‘Candidatus Glomeribacter gigasporanum’ (36), el que solo fue detectado en el citoplasma de esporas dormantes y micelios de HMA Gigaspora margarita, Scutellospora persica (Koske & Walker Walker & Sanders) y Scutellospora castanea (Walker).
Análisis electroforéticos revelan que la comunidad bacteriana asociada con las esporas depende más del hongo micorrízico arbuscular que de la identidad de la planta hospedera (37).
Bacterias asociadas con esporas HMA colonizan principalmente la pared celular externa de las esporas y raras veces penetran en el interior de la pared más interna (34). Aunque se han observado algunas bacterias en el citoplasma de esporas de HMA (14, 32), el papel de estas bacterias aún no se ha esclarecido, ellas pueden estimular la germinación de las esporas por erosión en las paredes (38), por la producción de compuestos estimuladores, por ejemplo el CO2 y otros compuestos volátiles, o por influencia en la adquisición de fósforo por los HMA (39).
En estudios previos de microorganismos asociados a las esporas, las bacterias fueron aisladas de los cultivos in vitro del HMA. No obstante, estas bacterias no cultivables en medios de cultivo no selectivos podían representar una parte significativa de la comunidad bacteriana asociada a las esporas. Solo una pequeña fracción (de 1 al 10 %) del total de la comunidad bacteriana es cultivable (40).
3. Hongos filamentosos en HMA
Los Actinomicetes se han encontrado en esporas y ascocarpos de HMA (38), y representan un componente normal de su microflora. Estos microorganismos incrementan la germinación de las esporas, debido a la producción de compuestos volátiles como por ejemplo 2-methylisoborneol (41).
Por su parte, otros han detectado hongos filamentosos en esporas de HMA. Ellos plantean que los HMA podían ser habitados por numerosos microorganismos y aislaron dos hongos Ascomicetos con fenotipos diferentes, de esporas vivas y muertas del HMA Scutellospora castanea con secuencias muy parecidas o semejantes a ascomycetes (ascomycete-like) (42). Estas secuencias pueden haber sido aisladas de hongos contaminantes que podían estar habitando la superficie de las esporas de HMA; otra posibilidad es que las secuencias fueran de hongos ascomicetos que viven en el interior de las esporas de HMA y la tercera situación es que podía ser que las secuencias sean originadas desde el genoma de la espora (42).
Investigaciones futuras podrían revelar si la presencia de tales hongos juega un papel importante en el funcionamiento de los HMA o su interacción con las plantas. ]]>
4. Levaduras en HMA
En los HMA también se han detectado levaduras, que fueron observadas asociadas a esporas de Glomus mosseae (cepa INCAM-2, cepario del INCA) después de un proceso de desinfección total denominado CCfT. Dos levaduras fueron caracterizadas (EndoGm1 y EndoGm11), ambos aislados fueron capaces de solubilizar fuentes de fósforo poco solubles (fosfato de hierro y calcio) y acumular polifosfatos. Los resultados del análisis del 18S ADNr revelaron que dichas levaduras pertenecían a los géneros Rhodotorula o Rhodosporidium (EndoGm1) y Cryptococcus (EndoGm11). Los resultados de los experimentos de inoculación revelaron que existe un efecto de las levaduras asociadas a las esporas sobre el crecimiento radical de plántulas de arroz, lo cual pudiera indicar una posible interacción tripartita entre las levaduras asociadas a las esporas, los HMA y las plantas (43).
Hipótesis del establecimiento de la simbiosis entre HMA y bacterias endosimbióticas
La simbiosis intracelular trajo consigo cuestiones fascinantes acerca de la adquisición, transmisión de los endosimbiontes y la evolución de adaptaciones recíprocas.
Margulis y Chapman (44) propusieron que en los miembros de la familia Gigasporaceae (Morton & Benny), se observan claramente dos posibles y opuestos tipos de simbiosis. En un estudio realizado en distintas áreas geográficas, se obtuvieron once aislamientos fúngicos, pertenecientes a seis especies diferentes de la familia Gigasporaceae: se apreció que Gigaspora margarita es capaz de albergar gran cantidad de endobacterias, Gigaspora roseae (Nicol. & Schenck) no alberga ninguna y en Gigaspora gigantea (Nicol. & Gerd.) se observan bacterias endosimbióticas, pero con características diferentes a las encontradas en otras especies de Gigaspora, debido a la presencia de bacterias con formas redondeadas o cocoides.
La compleja situación observada en la familia Gigasporaceae, puede deberse a la temporal pero interesante asociación de los HMA seleccionados por los simbiontes bacterianos (bacterias del suelo de vida libre) desde el ambiente (simbiosis cíclica). Aunque la existencia de endocitosis y adquisición de la bacteria es un evento muy raro en hongos, debido a la pared que rodea las hifas, algunos Zygomicetes pueden representar un caso especial entre los hongos, solo se conoce que estos tienen la posibilidad de establecer relaciones cíclicas con cianobacterias (14). ]]> La simbiosis permanente es el resultado de la adquisición de bacterias durante la evolución, mostrada solo por transmisión vertical, durante las generaciones de los endosimbiontes. La reproducción asexual típica de los HMA y la naturaleza cenocítica del micelio en los Zygomicetes, son factores que pueden facilitar este tipo de transmisión.
La simbiosis permanente ha sido muy discutida y es la que cuenta con un mayor número de votos a su favor. En el 2000 se demostró, con el uso de análisis celulares y moleculares, que las bacterias se mueven de una generación fúngica a la siguiente, por lo que bacterias presentes en el interior de HMA pueden llevar millones de años en sus estructuras, teniendo en cuenta que estos hongos datan de alrededor de millones de años (14).
Aislamiento de microorganismos asociados y metodologías de desinfección de esporas utilizadas
La desinfección total de las esporas ha sido intentada con diferentes objetivos, entre estos para lograr cultivos in vitro de los HMA; pero si se desean aislar bacterias del interior de las esporas, esto es imprescindible. La descontaminación de la superficie es requerida para evitar la proliferación de microorganismos contaminantes y asegurar que estos no influyan en los resultados experimentales, siendo las esporas frescas más sensibles al proceso de descontaminación que las esporas maduras (34).
Las esporas antes del proceso de desinfección presentan una coloración oscura a su alrededor debido a los contaminantes. Cuando las esporas son desinfectadas e incubadas en medio de cultivo Agar agua, la germinación de las esporas es limitada; niveles de germinación del 28 % fueron obtenidos después de una exposición en Cloramina T al 5 % por 120 minutos, porque la Cloramina T inhibe la germinación (34).
Existe mucha contradicción en cuanto a la eficiencia o no del método de Mosse. El planteó que con el uso de una solución de Cloramina T al 2 %, 200 µg.mL-1 de estreptomicina y trazas de detergente biológico se logra un 90 % de esporas descontaminadas (43); sin embargo, informaron que con dicho método solo se conseguía del 32-64 % de esporas axénicas (45).
En la literatura se citan otros métodos para la desinfección de las paredes de las esporas en dependencia del objetivo que se persigue, por ejemplo:
Inmersión en hipoclorito de sodio al 0.5 % por 20 minutos, seguido de tres lavados con agua destilada estéril e inmersión en una solución de sulfato de estreptomicina 100 mg.L-1 por 30 minutos y nuevamente tres lavados con agua destilada estéril (46). ]]> Inmersión por diez minutos en hipoclorito de sodio al 0.5 %, seguido de dos lavados con agua destilada estéril e inmersión en una solución de sulfato de estreptomicina 100 mg.L-1 por 10 minutos y nuevamente tres lavados con agua estéril (46, 47).
Lavado con alcohol 70 %, seguido de dos lavados con agua estéril e inmersión en solución de Cloramina T al 1 % por 10 minutos, seguido de tres lavados con agua estéril (47, 48).
Inmersión en Cloramina T 2 % durante un tiempo entre 10 y 120 minutos (34).
Inmersión en solución 1:1 de Cloramina T 2 %, sulfato de estreptomicina 200 mg.L-1 por 30 minutos, seguido de tres lavados con agua estéril durante 30 minutos (47, 49).
Inmersión en solución de Cloramina T 2 % durante cinco minutos, seguido de tres lavados en agua destilada estéril por el mismo tiempo (47).
Inmersión en solución de Cloramina T 4 %, 300 ppm de estreptomicina por 30 minutos y lavar cinco veces por una hora con agua filtrada y destilada estéril (13, 50).
Otros utilizaron la metodología anterior con una variante; en lugar de estreptomicina 300 ppm, usaron estreptomicina al 0.04 % (14).
Los métodos de desinfección superficial no son capaces de eliminar el 100 % de los microorganismos asociados, debido a la cantidad de microorganismos que pudiera estar localizada en el lumen de las esporas, así como incrustadas en las paredes de las hifas (47).
En la literatura se encuentran publicados un gran número de métodos rutinarios, con el objetivo final de proveer la superficie desinfectada de las esporas. El éxito en el uso de estas metodologías de desinfección es altamente dependiente de cómo son manipuladas las técnicas y sobre qué material se aplican.
Sin embargo, se ha propuesto una metodología de desinfección eficiente por un colectivo (51) del Instituto Nacional de Ciencias Agrícolas (INCA) y la Universidad de La Habana (UH), en un trabajo conjunto que se realizó con esporas frescas del HMA Glomus mosseae, las que fueron completamente desinfectadas y la limpieza de sus paredes puede ser apreciada en la Figura 1. Esta desinfección puede contribuir al estudio de la ultraestructura del esporangio, sus núcleos, sus sistemas de membranas, sus diferentes paredes celulares y, por supuesto, el objetivo principal para la que fue concebida, el estudio de microorganismos estrechamente asociados a las esporas de HMA Glomus mosseae (49). ]]>
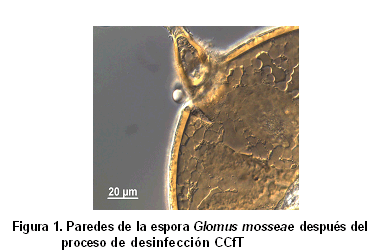
Dicha metodología se denominó CCfT y en ella se emplean diferentes combinaciones de Cloramina T, Tween y Cefalexina. El no crecimiento microbiano fue verificado durante siete días en tres medios de cultivos diferentes LGI-P (libre de nitrógeno, 10 % de sacarosa y pH 5,5) (52), AN (Agar nutriente) (53) y SYP (extracto de levadura, sacarosa y sales, pH 6,2) (54), en los cuales se habían inoculado esporas desinfectadas e intactas; lo inverso ocurrió cuando se inocularon en placas de cultivo esporas desinfectadas y maceradas (51), lo cual evidenció que las paredes estaban completamente desinfectadas y que en el macerado existen microorganismos que pudieran haber estado adheridos muy estrechamente en las paredes e introducidos en poros de estas o que simplemente vivían en el interior de las esporas.
Conclusiones
El concepto endosimbiosis da la posibilidad de estudiar e investigar la relación entre células y genomas. Este informe resalta que las micorrizas tienen un interés particular en un contexto evolutivo, porque ofrecen un selecto ejemplo de interacciones entre las plantas, los hongos y las bacterias u otros microorganismos.