0258-59360258-5936S0258-59362008000400011Cuba00122008001220082947784Establecimiento y validación de una colección núcleo en el germoplasma cubano de papa
Ms.C. J. G. CastilloI; Dra.C. Ana EstévezII; Ms.C. J. L. SalomónIII; Ms.C. Aymara PérezIV y Ursula OrtizIV; Dra.C. Zoila FundoraV
III Investigador Auxiliar ]]>
IV Especialista del departamento de Genética y Mejoramiento Vegetal, Instituto Nacional de Ciencias Agrícolas, gaveta postal 1, San José de las Lajas, La Habana, CP 32 700
V Investigadora Titular del departamento de Genética y Mejoramiento Vegetal, Instituto de Investigaciones Fundamentales en Agricultura Tropical (INIFAT), Santiago de las Vegas, La Habana, Cuba.
ABSTRACT
For four potato campaigns, the available morphoagronomic characterization data of 351 accessions of wild species and cultivated varieties from Cuban germplasm bank were used to establish and validate a core collection. Qualitative and quantitative characters related to tuber, crop behaviour vs. Alternaria solani and PLRV were evaluated. The main component and cluster analyses were based on the square Euclidean distance matrix for group selection. Means, standard deviations, variation ranges and coefficients of the cluster were calculated for quantitative characters, as well as the state appearance frequencies of qualitative attributes and Alternaria solani and PLRV occurrence. Core collection was selected by a stratified sampling, also having in mind the geographic origin of accessions. Results enabled to establish and validate a core collection in Cuban potato germplasm, adequately representing 21.9 % of base collection; therefore, maximizing both qualitative and quantitative characters. An appropriate geographical distribution was observed when core collection accessions are derived from eight out of the nine countries of origin.
Key words: potato, germplasm, core collections
RESUMEN
]]>
El trabajo se desarrolló a partir de los datos de la caracterización morfoagronómica disponible de 351 accesiones de especies silvestres y variedades cultivadas del banco de germoplasma cubano de papa, durante cuatro campañas, con el objetivo de establecer y validar una colección núcleo. Se evaluaron los caracteres cualitativos y cuantitativos relacionados con el tubérculo, el comportamiento ante el hongo Alternaria solani y la incidencia del virus del enrollamiento de la hoja (PLRV). Se realizaron análisis de componentes principales y conglomerados, sobre la base de la matriz de distancias Euclidianas al cuadrado para la selección de los grupos. Se calcularon las medias, desviaciones estándar, los rangos de variación y coeficientes de variación de los grupos para los atributos cuantitativos, así como las frecuencias de aparición de estados de los atributos cualitativos e incidencia del hongo Alternaria solani y el PLRV. La colección núcleo se seleccionó mediante un muestreo estratificado, teniendo en cuenta, además, el origen geográfico de las accesiones. Los resultados permitieron establecer y validar una colección núcleo en el germoplasma cubano de papa, que representa adecuadamente la colección base de partida. La selección de la colección núcleo representa el 21.9 % de la colección base y en ella se maximizan tanto los caracteres cuantitativos evaluados como los cualitativos. Existió una representación geográfica apropiada en tanto las accesiones de la colección núcleo proceden de ocho de los nueve países de origen.
Los bancos de germoplasma del mundo contienen colecciones de recursos genéticos, presentes en las especies cultivadas para la conservación a largo plazo y facilitar que los fitomejoradores, investigadores y otros usuarios accedan a ellos con facilidad. En los últimos años, han habido enormes progresos en la colecta y conservación de estos recursos, tanto es así que muchos bancos de germoplasma vegetal enfrentan hoy grandes problemas de tamaño y organización. Algunas colecciones han crecido tanto, que resulta difícil conservar y usar la diversidad genética que contienen, yendo así en contra de los objetivos para los cuales se establecieron (1). ]]>
Cuando se comprendió que el gran tamaño de algunas colecciones podría desalentar su uso, se propuso establecer una colección limitada o “núcleo” a partir de una colección existente. La colección núcleo, dada la semejanza mínima que hay entre una y otra de sus entradas, es de tamaño reducido y representa la diversidad genética de una colección de mayor tamaño de una especie cultivada, de una especie silvestre o de un grupo de especies (2). La colección núcleo no reemplaza la colección base o el material del cual se obtiene. Desde que se expuso esta propuesta, ha aparecido una bibliografía considerable sobre la teoría y práctica de las colecciones núcleo y en ella se encuentran muchos ejemplos de este enfoque. Las colecciones núcleo han sido aceptadas como herramientas eficaces, para mejorar la conservación y el uso de las colecciones de recursos genéticos (1). En el caso de la papa se han realizado diferentes estudios, que han estado encaminados fundamentalmente al establecimiento de una colección núcleo del cultivo de la subespecie Andígena (3). Por otra parte, se han realizado algunos estudios en el US Potato Genebank utilizando parámetros geográficos (4) y otros estudios para determinar el tamaño óptimo de una colección núcleo de papas tetraploides (5). El Instituto Nacional de Ciencias Agrícolas (INCA) cuenta con una colección de germoplasma superior a las 650 accesiones, que se incrementa cada año a partir de clones y variedades obtenidas en el programa de mejoramiento. El manejo de esta colección resulta difícil y costoso, pues cada año se necesita plantarla en campo, conservarla in vitro y en cámaras frías durante nueve meses. Es por ello de vital importancia contar con una colección reducida, bien caracterizada, que contenga el máximo posible de variabilidad genética y que, en casos de urgencia extrema, se le dediquen todos los esfuerzos y recursos a su mantenimiento y conservación. Es por todo ello que el presente trabajo estuvo dirigido a establecer y validar una colección núcleo de papa a partir de una representación importante de la colección base, caracterizada para diferentes descriptores morfoagronómicos así como su comportamiento ante el hongo Alternaria solani y la incidencia del virus del enrollamiento de la hoja (PLRV).
MATERIALES Y MÉTODOS
El trabajo se desarrolló a partir de los datos de caracterización disponibles de 351 accesiones de especies silvestres y variedades cultivadas provenientes de la colección del banco de germoplasma del INCA, las cuales fueron plantadas durante cuatro campañas (2001-2002, 2002-2003, 2003-2004, 2004-2005), en la finca “Las Papas”, sobre un suelo Ferralítico rojo (6). Se plantaron 10 plantas por cada accesión y las labores culturales y fitosanitarias fueron realizadas según el instructivo técnico del cultivo (7). La muestra caracterizada (colección base) representa el 54 % de la colección total. ]]>
Se evaluaron los siguientes caracteres: ð cualitativos: color del tubérculo, forma del tubérculo y profundidad de los ojos, ð cuantitativos: número de tubérculos por planta, masa promedio (kg), rendimiento por planta (kg), rendimiento total (t.ha-1) y altura de la planta (cm). Se evaluó el comportamiento de las accesiones ante Alternaria solani y el PLRV; en el caso de la primera, según una escala de nueve grados propuesta por Horsfall y Barrat (8), a los 65 días de plantada y en el segundo caso, se evaluó en porcentaje de afectación según la metodología de Cordero (9). Los datos de ambos orígenes se tomaron de la realizada en la evaluación morfoagronómica anteriormente descrita. Los datos promedio de los cuatro años fueron sometidos a un análisis de componentes principales sobre la base de la matriz de datos estandarizados (10). Se seleccionaron los componentes que mayor variabilidad tuvieron en la población de datos, con valores propios mayores que uno, y se tomó como criterio de selección los autovectores que tuvieron los valores mayores y cercanos entre sí. Con las coordenadas de los cuatro primeros componentes, se realizó un análisis de conglomerados (cluster), sobre la base de la matriz de distancias Euclideanas al cuadrado mediante el programa SPSS ver. 11.5 utilizando un análisis de K-medias y un modelo de iteración estimado en 10 iteraciones como máximo, formándose 10 grupos como el mejor criterio de agregación y clasificación. Durante el establecimiento de la colección núcleo, el peso relativo de cada grupo en la variabilidad total fue determinado por su relativa diversidad, o sea, por la mayor o menor presencia de accesiones dentro de cada tipo, incluyendo además el criterio de los especialistas del cultivo. Se realizó la selección de las accesiones representantes de cada grupo, mediante un muestreo estratificado (11), considerando además la representatividad del origen geográfico, tanto como fuera posible. Se calcularon las medias, desviaciones estándar, los rangos de variación y coeficientes de variación (para eliminar el efecto de la escala) de los grupos del cluster para los atributos cuantitativos, así como las frecuencias de aparición de los estados de los atributos cualitativos (%), con el objetivo de comparar el núcleo seleccionado con la colección base, así como la variabilidad de cada grupo del núcleo con su grupo correspondiente de la colección base. Se calcularon, además, las coincidencias entre las frecuencias de los estados de los descriptores mínimos cualitativos, así como las frecuencias de incidencia del hongo Alternaria solani y el PLRV, a partir de la estimación de la correlación de rangos de Spermann, entre las frecuencias de aparición en las colecciones base y núcleo, utilizando para ello el paquete estadístico SPSS ver. 11.5.
RESULTADOS Y DISCUSIÓN
]]>
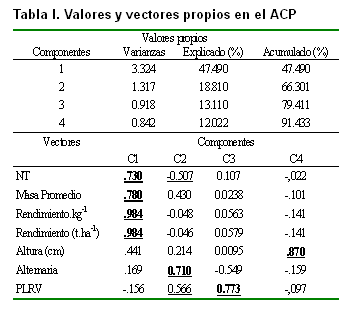
En la Tabla I se presentan los valores y vectores propios del análisis de componentes principales a las seis variables independientes analizadas. Como se observa en las primeras cuatro componentes, se extrajo el 91.43 % del total de la variabilidad presente en los caracteres estudiados. Las variables que más contribuyeron a la formación de las componentes fueron el número de tubérculos, la masa promedio y el rendimiento por planta y total en la C1; en la C2 la mayor contribución la realizó la incidencia del hongo Alternaria solani, mientras que en la C3 fue la afectación por el PLRV. La altura fue la variable que más contribuyó en la formación de la componente 4.
Resultados similares a estos fueron informados anteriormente, los que siempre han encontrado que las variables analizadas tienen una fuerte relación entre sí, por ser componentes fundamentales en el rendimiento de la papa (12, 13, 14). Se ha observado también alta correlación entre los caracteres cualitativos usando métodos multivariados (15). En la Tabla II se presentan los grupos formados mediante el análisis de conglomerados, tomando como base las coordenadas de las cuatro primeras componentes, donde el mejor criterio de clasificación fue la formación de 10 grupos, el cual presentó una distribución adecuada y la presencia de al menos una accesión en cada uno de ellos. Los porcentajes de representatividad estuvieron entre 0.3 y 20 %. El número de accesiones en los diversos grupos de una colección es un buen indicador general de la utilidad de los grupos o la diversidad que hay en ellos (1).
]]>
En la Tabla III se presentan las accesiones seleccionadas para la conformación de la colección núcleo, a partir de la aplicación de un muestreo estratificado combinado con criterios de representatividad dentro de cada grupo y tomando como base el nivel de utilización de las accesiones. En este mismo sentido se consideró, en cierta medida, la procedencia geográfica de las muestras. Para el caso específico de la papa se ha trabajado en el tamaño óptimo para conformar una colección núcleo en la subespecie Andígena (5). Estos resultados están en concordancia con otros, donde se plantea que la mejor clasificación es la que minimiza la variación dentro de los grupos y maximiza la variación entre los grupos (16).
El proceso de estratificación se puede desarrollar con relativa facilidad, dividiendo los grupos taxonómicos en tipos de cultivo o en grupos ecogeográficos bien establecidos (17). Se han desarrollado otros procesos de estratificación de forma más compleja e involucrando una mayor cantidad de pasos (18, 19). Otros han propuesto métodos similares al usado en este estudio en el establecimiento de colecciones núcleos, pero utilizando otros métodos de agrupamiento y trabajando con un número diferente de accesiones (20). En este mismo sentido, se han utilizado también análisis isoenzimáticos y moleculares para darle mayor fortaleza a este procedimiento (21, 22). Comparación de la propuesta de colección núcleo con la colección base. Los estadísticos descriptivos fundamentales de los caracteres cuantitativos evaluados de ambas colecciones aparecen en la Tabla IV. Los valores promedio de todos los caracteres de la colección núcleo fueron similares a la colección base; sin embargo, en la colección núcleo se obtuvieron coeficientes de variación mayores que en la colección base en todos los casos, excepto en el número de tubérculos, lo cual nos indica que la selección efectuada maximizó el rango de variación de los caracteres. Estos resultados están en concordancia con otros planteamientos que expresan que los rangos de variación deben permanecer similares a los grupos de donde se derivaron (1), o pueden incluso ser superiores (20, 23).
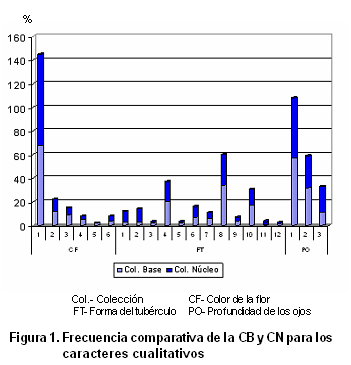
En la Tabla V y en la Figura 1 se presenta la comparación gráfica de cada uno de los descriptores cualitativos. Se observa, en todos los casos, un comportamiento similar en el estado de las frecuencias para los caracteres cualitativos en ambas colecciones. De forma general, se maximizan algunos estados en la colección núcleo, mientras que otros disminuyen, existiendo solamente la ausencia de un estado en el color de la piel del tubérculo.
]]>
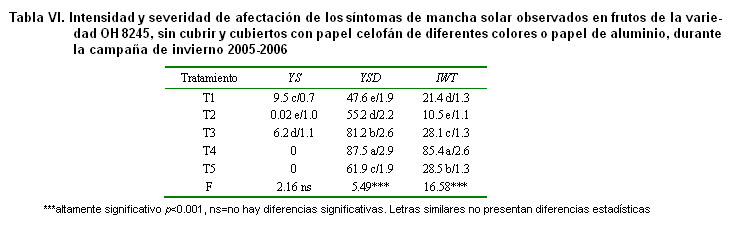
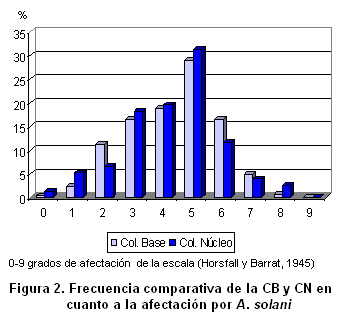
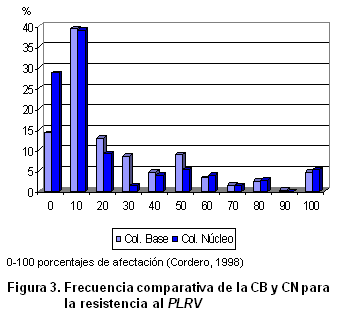
En este mismo sentido, existe una correlación Spearman alta y significativa para los caracteres cualitativos y para la afectación por Alternaria solani y la del virus de enrollamiento de la hoja, entre las frecuencias de la colección base y la colección núcleo (Tabla VI). En las Figuras 2 y 3 se muestra de forma gráfica el comportamiento similar entre ambas colecciones para estas dos afectaciones. En el primer caso se manifiestan dentro de la colección accesiones resistentes y susceptibles, aunque es mayoritaria la presencia en la colección núcleo de las primeras. El hongo Alternaria solani es la enfermedad más importante que afecta al cultivo de la papa en Cuba y ha sido una de las líneas priorizadas en el programa de mejoramiento cubano desde sus inicios (13, 24). Otros trabajos avalan la importancia de contar con variedades resistentes a este patógeno en los bancos de germoplasma del género Solanum (25).
]]>
La colección núcleo está integrada por 77 accesiones, lo que representa el 21.9 % de la colección base (Tabla VII). La selección fluctuó entre un 11.6 y 100 % en cada uno de los grupos. El tamaño excede al propuesto por Brown (79), pero se considera adecuado, si se tiene en cuenta que se trabajó con un 54 % de la colección completa y el tamaño es manejable para las condiciones experimentales de trabajo. Resultados similares ya han sido obtenidos, algunos de ellos trabajando con un número menor de accesiones (20) y otros con un número mayor (11).
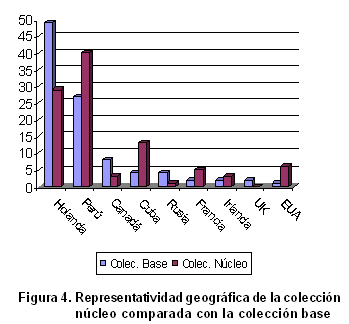
Las accesiones seleccionadas para conformar la colección núcleo tienen una representatividad geográfica adecuada (Figura 4), pues proceden de ocho de los nueve países a que pertenecen las muestras introducidas al país. El origen geográfico de las accesiones es uno de los aspectos esenciales a tomar en cuenta en el establecimiento de las colecciones núcleo (30) y, en este caso, es muy importante por cuanto Cuba no es centro de origen de este cultivo.
]]>
Descripción de la colección núcleo Grupo 1: Compuesto por una especie silvestre y cinco variedades cultivadas caracterizadas por su susceptibilidad elevada al PLRV y una resistencia media a A. solani. Los rendimientos fluctúan de medios a bajos. Grupo 2: Compuesto por tres especies silvestres y cinco cultivadas con resistencia elevada al PLRV y comportamiento medio ante A. solani. Presenta rendimientos de bajo a medios. Grupo 3: Compuesto por ocho variedades cultivadas con alto valor del número de tubérculos y de masa promedio, resistencia moderada a A. solani y buen comportamiento ante el PLRV. Se encuentran las variedades comerciales foráneas como Desirée y la variedad cubana Yara (C-63). Grupo 4: Compuesto por cuatro especies silvestres, con rendimientos muy bajos y alta resistencia al PLRV y cierta resistencia a A. solani y la mayor altura de la planta todos los grupos. Grupo 5: Compuesto por tres variedades con comportamiento medio para el rendimiento y sus componentes, susceptibilidad a A. solani y buen comportamiento ante el PLRV. Grupo 6: Compuesto por 15 especies silvestres caracterizadas por presentar un comportamiento extremadamente bajo para el rendimiento y sus componentes, pero una gran resistencia al PLRV, presentan susceptibilidad moderada a A. solani. Grupo 7: Compuesto por 16 variedades cultivadas caracterizadas por presentar altos valores para el rendimiento y sus componentes, susceptibilidad ante la incidencia de A. solani y afectaciones moderadas al PLRV. Compuesto por la mayoría de las variedades comerciales foráneas que se plantan en la actualidad en nuestro país y tres variedades cubanas. Grupo 8: Compuesto por nueve variedades comerciales con rendimientos altos pero altamente susceptibles a A. solani y susceptibilidad media al PLRV. ]]>
Grupo 9: Compuesto por seis variedades, cinco de ellas silvestres, que se caracterizan por ser altamente susceptibles tanto para A. solani como al PLRV. Grupo 10. Compuesto por una sola variedad, la cual se aisló del resto de los grupos por presentar un alto rendimiento, la menor altura y un comportamiento medio ante A. solani y el PLRV. De forma general, se puede plantear que la metodología empleada permitió establecer una colección núcleo en el germoplasma cubano de papa, que representará adecuadamente la colección base de partida. La selección de la colección núcleo representa el 21.9 % de la colección base y en ella se maximizan tanto los caracteres cuantitativos evaluados como los cualitativos. Se demostró que existe una composición adecuada tanto para la resistencia a A. solani como ante la incidencia del PLRV con la presencia de especies silvestres y variedades cultivadas resistentes, susceptibles y de comportamiento intermedio. Existió una adecuada representación geográfica en tanto las accesiones de la colección núcleo proceden de ocho de los nueve países de origen.
Referencias
1. Van Hintum, J. L.; Brown, A. H. D.; Spillane, C. y Hodgkin, T. Colecciones núcleos de recursos fitogenéticos. IPGRI Boletín Técnico. 2003, no. 3, p. 44. 2. Frankel, O. H.; Brown, A. H. D. Plant genetic resources today: A critical appraisal. En: Crop genetic resources: Conservation and evaluation (J. H. W. Holden y J. T. Williams, Eds.) Massachusetts: Unin, Winchester, 1984, p. 161-170. 3. Huamán, Z.; Ortiz, R.; Zhang, D. y Rodríguez, F. Isozyme analysis of entire and core collections of Solanum tuberosum subsp. andigena potato cultivars. Crop Science, 2000, vol. 40, no. 1, p. 273-276. 4. Río, A. H. del; Bamberg, J. B. y Huaman, Z. Genetic equivalence of putative duplicate germplasm collections held at CIP and US Potato Genebanks. Amer. Journal of Potato Res., 2006, vol. 83, no. 2, p. 279-285. 5. Chandra, S.; Huaman, Z.; Hari Krishna, S. y Ortiz, R. Optimal sampling strategy and core collection size of Andean tetraploid potato based on isozyme data. A simulation study. Theoretical and Applied Genetics, 2002, vol. 104, no. 8, p. 1325-1334. 6. Cuba. Minagri. Instituto de Suelos. Nueva versión de la clasificación genética de los suelos de Cuba. La Habana: Agrinfor, 1999, 64 p. 7. Cuba-MINAGRI. Normas técnicas para el cultivo de la papa. La Habana, Ministerio de la Agricultura. 1990, p. 46. 8. Estévez, A.; Salomón, J. L.; Castillo, J. G.; Ortiz, U. y Ortiz, E. Regionalización de clones cubanos de papa (Solanum tuberosum L.). Cultivos Tropicales, 2005, vol. 26, no. 3, p. 59-63. 9. Cordero, M. Bases metodológicas para la evaluación de la resistencia al PLRV en genotipos de papa (Solanum tuberosum) en Cuba. (Tesis de Maestría), UH, 1998. 10. Franco, T. L. e Hidalgo, R. Análisis Estadístico de Datos de Caracterización Morfológica de Recursos Fitogenéticos. IPGRI Boletín Técnico, 2003, no. 8, p. 89. 11. Quero-Garcia, J.; Noyer, J. L.; Perrier, X.; Marchand, J. L. y Lebot, V. A germplasm stratification of taro (Colocasia esculenta) based on agro-morphological descriptors, validation by AFLP markers. Euphytica, 2004, vol. 137, no. 3, p. 387-395. 12. González, M. E. Mejoramiento por hibridación de la papa (Solanum tuberosum L.) en Cuba. (Tesis de Doctorado), INCA, 1998. 13. Castillo J.; Estévez, A.; González, M. E. y Salomón, J. L. Caracterización morfoagronómica del germoplasma cubano de papa (Solanum ssp). Evaluación de las especies silvestres. Parte I. Cultivos Tropicales, 2007, vol. 28, no. 2, p. 63-64. 14. Estévez, A.; González, M. E y Cordero, M. Primeras variedades cubanas de papa. Cultivos Tropicales, 1994, vol. 15, no. 2, p. 74-78. 15. Cardi, T.; Mazzei, M. y Frusciante, L. Field variation in a tetraploid progeny derived by selfing a Solanum commersonii (+) S. tuberosum somatic hybrid: A multivariate analysis. Euphytica, 2002, vol. 124, no. 1, p. 111-119. 16. Crossa, J. y Franco, J. Statistical methods for classifying genotypes. Euphytica, 2004, vol. 137, no. 1, p. 19-37. 17. Zhang, X.; Zhao, Y.; Cheng, Y.; Feng, X.; Guo, Q.; Zhou, M. y Hodgkin, T. Establishment of sesame germplasm core collection in China. Genet. Resour. Crop Evol., 2000, vol. 47, p. 273-279. 18. Bisht, I.S.; Mahajan, R.K; Lokknathan, T.R.; Agrawal, R.C.. Diversity in Indian sesame collection and stratification of germplasm accessions in different diversity groups. Genetic Resour. and Crop Evol., 1998, vol. 45, p. 325-335. 19. Wang, J. C.; Hu, J.; Xu, H. M. y Zhang, S. A strategy on constructing core collections by least distance stepwise sampling. Theoretical and Applied Genetics, 2007, p. 1432-2242. 20. Fundora, Z. M. /et al./. Establecimiento de colecciones “núcleo” en especies de hortalizas, granos básicos y oleaginosas. Propuesta premio MINAGRI. 2008. 21. Ghislain, M.; Zhang, D.; Fajardo, D.; Huamán, Z. y Hijmans, R. J. Marker-assisted sampling of the cultivated Andean potato Solanum phureja collection using RAPD markers. Genetic Resources and Crop Evolution, 1999, vol. 46, no. 6, p. 1573-5109. 22. Chavarriaga-Aguirre, P.; Maya, M. M.; Tohme, J.; Duque, M. C.; Iglesias, C.; Bonierbale, M. W.; Kresovich, S. y Kochert, G. Using microsatellites, isozymes and AFLPs to evaluate genetic diversity and redundancy in the cassava core collection and to assess the usefulness of DNA-based markers to maintain germplasm collections. Molecular Breeding, 1999, vol. 5, no. 3, p. 263-273. 23. Malosetti, M.; Abadié, T. y Germán, S. Comparing strategies for selecting a core subset for the Uruguayan barley collection. Plant Genetic Resources Newsletter, vol. 121, p. 20-26. 24. Estévez, A.; González, M. E.; Castillo, J. y Salomón, J. L. Anita una nueva variedad cubana de papa. Cultivos Tropicales, 2006, vol. 27, no. 1, p. 69-70. 25. Chaerani, R.; Remmelt, G.; Piet, S. y Voorrips, R. E. Assessment of early blight (Alternaria solani) resistance in tomato using a droplet inoculation method. Journal of General Plant Pathology, 2007, vol. 73, no. 2, p. 96-103. 26. Novy, R. G.; Gillenand, A. M. y Whitworth, J. L. Characterization of the expression and inheritance of potato leafroll virus (PLRV) and potato virus Y (PVY) resistance in three generations of germplasm derived from Solanum etuberosum. Theoretical and Applied Genetics, 2007, vol. 114, no. 7, p. 1161-1172. 27. Vaikonen, J. P. Natural genes and mechanisms for resistance to viruses in cultivated and wild potato species (Solanum spp.). Plant Breeding, 1994, vol. 11, no. 2, p. 1-16. 28. Gillen, A. M. y Novy, R. G. Molecular characterization of the progeny of Solanum etuberosum identifies a genomic region associated with resistance to potato leafroll virus. Euphytica, 2007, vol. 155, no. 3, p. 403-415. 29. Brown, A. H. D. The case for core collections. En: The use of plant genetic resources. Cambridge University Press, Cambridge, Reino Unido. 1989, p. 136-156. 30. Del Río, A. y Bamberg, J.B. Geographical parameters and proximity to related species predict genetic variation in the inbred potato species Solanum verrucosum Schlechtd. Amer. Jour. of Potato Research, 2004, vol. 81, no. 1, p. 55-62.
Recibido: 7 de julio de 2008 Aceptado: 3 de noviembre de 2008

 ]]>
]]>
 ]]>
]]>