0258-59360258-5936S0258-5936201400010000400032014000320143512534Efecto de los brasinoesteroides y un inhibidor de su biosíntesis en plántulas de dos variedades de tomate sometidas a estrés salino
Effect of brassinosteroids and its biosynthesis inhibitor in two varieties of tomato´s seedlings under salt stress
Lic.Yanelis Reyes Guerrero,I Lic. Lissy Rosabal Ayan,II Ing. Lisbel Martínez González,III Dr.C Luis M. Mazorra Morales,IV Dra.C. Miriam Núñez VázquezV
IAspirante a Investigador, departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas(INCA), gaveta postal 1, San José de las Lajas, Mayabeque, CP 32700. IIReserva Científica, departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, CP 32700. IIIEspecialista, departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, CP 32700. IVInvestigador Agregado, departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, CP 32700. ]]>
VInvestigadora Titular, departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, CP 32700.
RESUMEN
Los brasinoesteroides (BRs) están considerados actualmente la sexta clase de hormonas vegetales. En los últimos años, varios autores han estudiado la protección que los BR le confieren a las plantas ante determinados tipos de estrés tanto bióticos como abióticos. En este trabajo se evaluó la germinación, el crecimiento inicial, los niveles de prolina y malondialdehído (MDA) y la actividad peroxidasa de dos variedades de tomate (INCA 9(1) y Amalia) sometidas a estrés salino y tratadas con un brasinoesteroide natural (24-epibrasinólida, EBL), un análogo espirostánico de brasinoesteroide (Biobras-16) y un inhibidor de la biosíntesis de los brasinoesteroides (Brz2001). Para ello semillas de estas dos variedades se trataron durante cuatro horas con EBL (0,1µmol.L-1), Biobras-16 (0,1µmol.L-1), Brz2001 (10 µmol.L-1), y la combinación de los brasinoesteroides con el inhibidor. Posteriormente se colocaron, en placas Petri durante siete días, en presencia o no NaCl (75 mmol.L-1). Se evidenció que ambos brasinoesteroides lograron recuperar la germinación de las semillas estresadas con NaCl, mientras que el Brz2001 con NaCl inhibió la germinación. En el caso del crecimiento se evidenció un efecto protector de la EBL en la longitud del tallo y la masa seca de las plántulas estresadas, mientras que en la longitud de la raíz tuvo un efecto negativo. Los niveles de prolina y la actividad peroxidasa aumentaron en todos los tratamientos excepto el control, mientras que los niveles de MDA se mantuvieron inalterables en todos los tratamientos. En el caso de la prolina la EBL logró reducir estos niveles en las plántulas estresadas.
Brassinosteroids (BRs) are currently considered the sixth class of plant hormones. In recent years, several authors have studied the BR protection conferred to the plants to certain types of biotic and abiotic stress. In this study, germination, early growth, levels of proline and malondialdehyde (MDA) and the peroxidase activity of two tomato varieties (INCA 9(1) and Amalia) subjected to salt stress and treated with a natural brassinosteroid (24-epibrassinolide, EBL), an spirostanic analogue of brassinosteroid (Biobras-16) and an inhibitor of the biosynthesis of brassinosteroids (Brz2001) were evaluated. Seeds of these varieties were treated for four hours with EBL (0,1µmol.L-1) Biobras-16 (0,1µmol.L-1) Brz2001 (10 µmol.L-1), and the combination of brassinosteroids with the inhibitor. Later, they were then placed in Petri dishes for seven days in the presence or absence of NaCl 75 mmol.L-1. It was evidenced that both brassinosteroids regained the germination of NaCl stressed-seeds, while the Brz2001 with NaCl inhibited germination. In the case of growth, EBL protected the stem length and the dry mass of the stressed seedlings, while it had a negative effect on root length. Proline levels and peroxidase activity increased in all treatments except the control, whereas MDA levels remained unchanged in all treatments. EBL reduced proline levels in stressed seedlings.
Los brasinoesteroides (BRs) están considerados actualmente la sexta clase de hormonas vegetales. Estos compuestos naturales, que están ampliamente distribuidos en el reino vegetal, tienen efectos pleiotrópicos en las plantas, entre los que se destaca su efecto estimulador del crecimiento (1) por estimulación del alargamiento celular y regeneración de la pared celular provocando un incremento de la biomasa y del rendimiento. Además, se encuentran involucrados en la desdiferenciación de protoplastos y la regulación de la diferenciación de elementos traquearios (2).
En los últimos años, varios autores han estudiado la protección que los BRs le confieren a las plantas ante determinados tipos de estrés tanto bióticos como abióticos (3, 4). Entre estos se encuentran altas y bajas temperaturas (5), sequía y salinidad (3) y exceso de metales pesados (6, 7). Además disminuyen los efectos del estrés causado por la falta de nutrientes y también incrementan la resistencia a herbicidas y a agentes patógenos (2).
La 24-epibrasinólida (EBL) es el brasinoesteroide natural más usado internacionalmente para estudiar los efectos de esta familia de compuestos en las plantas, y diversas han sido las formas de aplicación y las concentraciones empleadas. También, para realizar experimentos actualmente se utilizan análogos, que son portadores de modificaciones en su estructura y, por tanto, en su actividad biológica, como es el Biobras-16 (BB-16). Este último es una formulación que se sintetiza en nuestro país, en el Centro de Estudios de Productos Naturales de la Facultad de Química de la Universidad de la Habana, que ha tenido muy buenos resultados en la agricultura (8).
El Brz2001 es un inhibidor de la biosíntesis de los brasinoesteroides. Este inhibidor induce cambios morfológicos en las plantas tales como enanismo y alteraciones en las hojas con encrespamiento de los bordes y color verde oscuro. Estos cambios son revertidos por la brasinólida. A pesar de que la estructura del Brz2001 es similar al uniconazol, un inhibidor de la biosíntesis de giberelina, las plantas tratadas con Brz2001 mostraron muy poca recuperación con la adición de ácido giberélico. La comparación de las respuestas en plantas de berro tratadas con brasinazol y Brz2001 han demostrado que este último es un inhibidor más especifico de la biosíntesis de los brasinoesteroides. Su efecto se ha demostrado en plantas dicotiledóneas como Arabidopsis thaliana (L.) Heynh., Nicotiana tabacum L.y Lepidium sativum L., pero en monocotiledóneas como arroz no produce ningún cambio. El Brz2001 se ha utilizado como complemento a los mutantes deficientes de BRs para dilucidar diferentes mecanismos de acción de estos compuestos en las plantas (9). En este estudio se utiliza para disminuir el contenido endógeno de brasinoesteroides y observar el efecto que esta deficiencia pudiera tener en la respuesta de las plántulas al estrés salino. ]]>
La salinidad es uno de los estrés más perjudiciales para las plantas. Las semillas son las primeras en enfrentar las condiciones de estrés, ya que la salinidad afecta tanto la reanudación del crecimiento activo del embrión como el crecimiento inicial de la plántula, a través de su influencia sobre diferentes procesos fisiológicos y bioquímicos. Los principales efectos que causa la presencia de sales en el medio son la disminución de la absorción de agua por las raíces (efecto osmótico), la absorción específica de iones que pueden provocar toxicidad y el desbalance nutricional provocado por modificaciones en la absorción de nutrientes esenciales (10).
Existen algunos estudios que plantean la inducción de tolerancia al estrés salino con la utilización de brasinoesteroides. Sin embargo, no se ha estudiado cómo la alteración de los niveles de hormona endógena puede afectar la respuesta a este estrés. Además de que existen pocos estudios en tomate sobre los efectos de los brasinoesteroides en la tolerancia al estrés salino.
Teniendo en cuenta estos antecedentes se realizó el siguiente trabajo con el objetivo de evaluar el efecto de la aplicación exógena de un brasinoesteroide natural y un análogo; así como el de un inhibidor de la biosíntesis, lo cual debe reducir la concentración endógena de estas hormonas, en la respuesta de plántulas de dos genotipos de tomate a la salinidad.
MATERIALES Y MÉTODOS
Para realizar el experimento se utilizaron dos variedades cubanas de tomate (Solanum lycopersicum), Amalia e INCA 9(1), dos brasinoesteroides, uno natural (24-epibrasinólida, EBL) y un análogo espirostánico, 25(R)-3a, 5a-dihidroxi- 5a-espirostan-6-ona cuya formulación se conoce como Biobras-16 (BB-16) y un inhibidor de la biosíntesis de brasinoesteroides, Brz2001. Las semillas de ambas variedades se sumergieron, durante cuatro horas, en soluciones de 24-epibrasinólida o Biobras-16 (0,1 µmol.L-1), sin y con el inhibidor Brz2001 (10 µmol.L-1), utilizándose la imbibición en agua como tratamiento control. Además se incluyó un tratamiento solamente con Brz2001 (10 µmol.L-1). Luego las semillas tratadas se pusieron a germinar sobre papel de filtro en placas Petri en solución de NaCl (75 mmol.L-1) o en agua destilada (control). En la tabla se muestra la descripción detallada de cada uno de los tratamientos. Se colocaron en cada placa 30 semillas y se utilizaron seis placas por tratamiento. Las mismas se mantuvieron bajo condiciones de oscuridad durante una semana a 25ºC. Al cabo de los siete días se evaluaron los siguientes indicadores: número de semillas germinadas, longitud de la raíz y del vástago, así como las masas fresca y seca de las plántulas. En el caso de las masas seca y fresca se conformaron tres muestras de cinco plantas cada una por tratamiento.
Además, al finalizar el experimento se realizaron algunas determinaciones bioquímicas a las plántulas de los tratamientos cuyo crecimiento lo permitió. Para ello, se tomaron 0,25 g de tejido fresco de cada tratamiento y se maceraron en nitrógeno líquido, adicionándole 2,5 mL de tampón fosfato 100 mmol.L-1, pH 7 y centrifugándose posteriormente a 10000 rpm por 20 minutos a 4ºC. Con el sobrenadante obtenido se estimó la peroxidación lipídica mediante la cuantificación del malondialdehído (MDA) (11), la actividad de las enzimas peroxidasas totales (12) y las proteínas solubles totales por el método de Bradford (13). El precipitado obtenido de los extractos, después de lavado tres veces con tampón fosfato, se trató con NaCl 1 mol.L-1 por 1 hora para extraer las peroxidasas unidas iónicamente a la pared celular (14), posteriormente se centrifugó a 10000 rpm por 15 minutos. Con el sobrenadante obtenido se realizó el ensayo de la actividad enzimática peroxidasa utilizando pirogalol como sustrato (12). Una unidad de actividad enzimática peroxidasa se definió como la cantidad de enzima que causó un incremento de 0,1 unidades de absorbancia por minuto a 25ºC. ]]>
Además, para la determinación del aminoácido prolina se tomaron 0,25 g de tejido fresco de cada tratamiento y se maceraron en nitrógeno líquido, adicionándole 10 mL de agua a 100ºC. El contenido de prolina se cuantificó a través del método descrito por Bates et al. (15) y se utilizó L-Prolina (Sigma) como patrón.
El experimento se repitió en dos ocasiones con comportamientos similares. Se muestran los resultados de una de las repeticiones. El procesamiento estadístico de los datos obtenidos consistió en el cálculo de las medias, las desviaciones estándar y los intervalos de confianza a p≤0,05.
RESULTADOS Y DISCUSIÓN
Para evaluar el efecto de la aplicación exógena de los brasinoesteroides y sus análogos; así como el de un inhibidor de la biosíntesis en plántulas de tomate ante estrés salino, se determinaron diferentes indicadores relacionados con la germinación y el crecimiento de plántulas etioladas de tomate de las variedades INCA 9(1) y Amalia.
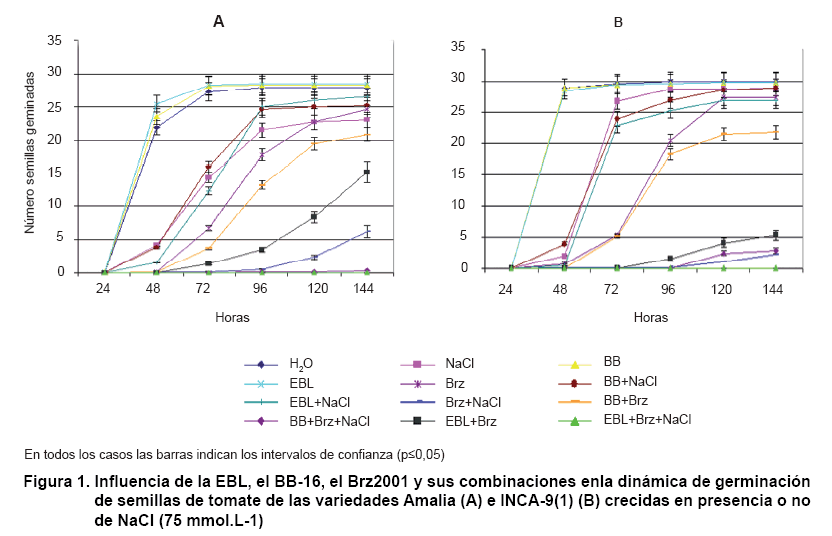
La dinámica de la germinación demostró que las semillas de tomate de ambas variedades cubanas, Amalia e INCA 9(1), tratadas con el inhibidor de la biosíntesis de los BRs, Brz2001 y crecidas en medio salino, disminuyen su porcentaje final de germinación, siendo más pronunciado cuando se utiliza el inhibidor. El NaCl disminuyó el porcentaje de germinación en la variedad Amalia y no así en la INCA 9(1). El tratamiento con Brz2001 retrasa la germinación, pero al final no hay diferencia con el control. El tratamiento con BRs no revirtió la reducción que la sal provocó en la variedad Amalia y el tratamiento con BRs conjuntamente con Brz inhibió el proceso de germinación de las semillas. Cuando las plántulas crecieron en medio salino y las semillas habían sido tratadas con Brz, se inhibió la germinación y esta respuesta se intensificó cuando se adicionaron los BRs (Figura 1).
]]>
Está bien establecido que los brasinoesteroides promueven la germinación de las semillas, particularmente en tomate (16) presumiblemente por la interrupción de la latencia de las semillas mediada por ABA y estimulando la germinación por la vía de la expansión del embrión (17).
Se ha demostrado que en los estadios tempranos de la germinación los niveles endógenos de brasinoesteroides como la brasinólida y la castasterona se elevan, así como los transcriptos de dos C-6 oxidasas (CYP85A1 y CYP85A6), enzimas relacionadas con la biosíntesis de los BRs (18).
Otros autores (19) han informado que el porcentaje de germinación de semillas de Brassica juncea se incrementó significativamente con la aplicación de 24-epibrasinólida especialmente a las concentraciones más bajas utilizadas (10-9 y 10-11 mol.L-1).
En experimentos realizados con el análogo de brasinoesteroide BB-6 en la germinación de la variedad de tomate Campbell-28, en condiciones salinas, no se modificó la dinámica de germinación aunque se encontraron ligeros aumentos en los porcentajes finales de germinación (8).
En experimentos realizados por nuestro grupo de trabajo se mostró un incremento en la velocidad de germinación con BB-16 (0,1 µmol.L-1) en semillas de tomate de la variedad INCA 9(1) sometidas a estrés salino, aunque no hubo diferencias en los porcentajes finales de germinación (20). ]]>
En este trabajo los brasinoesteroides no estimularon la germinación ni en condiciones normales ni en presencia de NaCl. Esto podría deberse a la concentración utilizada en los ensayos, que es 100 veces superior a las informadas en la literatura para la estimulación de la germinación. Se ha planteado que concentraciones altas (1 µmol.L-1) de estas hormonas provocan retraso de la germinación (20). Además , se ha observado que la administración de cantidades excesivas de brasinólida y castasterona provoca crecimiento anormal de los órganos vegetales como inflamación y distorsión de los hipocótilos y raíces, tanto en la luz como en la oscuridad (21).
También las condiciones en que se realizó el experimento pueden estar relacionadas con los resultados, pues estas son plantas crecidas en la oscuridad. Algunos autores han reportado que, en la oscuridad, los mutantes de Arabidopsis deficientes o insensibles a los BRs presentan un fenotipo des-etiolado y una des-represión de los genes inducidos por la luz (2).
Recientemente, se han obtenido evidencias de que estos compuestos regulan negativamente la fotomorfogénesis, a través de una compleja red regulatoria que incluye la interacción con otras hormonas (22).
Además, el Biobras-16 logró revertir parcialmente los efectos adversos del Brz2001, en ambas variedades, mientras la EBL no tuvo este comportamiento. Esto podría deberse a las diferencias estructurales entre estos compuestos, pues el Biobras-16 es un análogo de la castasterona, el brasinoesteroide más activo en tomate. La castasterona (CS) es el penúltimo compuesto de la biosíntesis de los brasinoesteroides y el precursor de la brasinólida (BL). La CS se convierte a BL por una reacción de oxidación en el C-6 (Baeyer-Villiger), lo que resulta en la formación de un inusual anillo lactona de siete miembros. Se ha postulado que la castasterona es el brasinoesteroide más activo en tomate durante el crecimiento vegetativo y que en este estadio los niveles de brasinólida son muy bajos (23).
Cuando las semillas se tratan con Brz y se colocan en presencia de NaCl, los BRs no pueden revertir el fuerte efecto inhibitorio de la combinación del estrés salino y la disminución del contenido endógeno de hormona. ]]>
En la Figura 2 se muestran las longitudes del vástago y la raíz de las plantulas de tomate de ambas variedades. Se puede observar el efecto negativo de la sal, en una concentración de 75 mmol.L-1, y el efecto inhibitorio de Brz2001 a una concentración de 10 µmol.L-1 en las plántulas etioladas de ambas variedades de tomate (Figura 2A). Se demostró que el tratamiento con el BB-16 incrementó la longitud del tallo de plántulas de la variedad Amalia sometidas al estrés salino (Figura 2A).
Se evidenció que en plántulas etioladas de tomate, la imbibición de las semillas con EBL, tuvo actividad biológica (inhibición de la raíz) (Figura 2B), pero no mostró los efectos clásicos de la elongación del tallo (Figura 2A). Esta respuesta puede deberse a la concentración utilizada, pues generalmente las concentraciones estimuladoras son más bajas. También este BR, aunque no mejoró la tolerancia de las plántulas a la salinidad, revirtió un poco la inhibición que provocó el Brz; sin embargo, este efecto no se observó en la germinación.
Además, se comprobó que en general, la aplicación de la EBL incrementó la inhibición del crecimiento de la raíz provocada por Brz2001, tanto en condiciones normales como en condiciones de estrés salino (Figura 2B).
Los efectos de los BRs en el crecimiento de la raíz son contradictorios, en algunos casos detectándose promoción y en otra inhibición. En general, los niveles de brasinoesteroides como la castasterona en raíces son más bajos que en los meristemos, y al parecer los requerimientos en este órgano también son menores (24). Los primeros informes mostraron que la EBL tenía efectos inhibitorios en la formación de las raíces en maíz, tomate, Arabidopsis y plántulas de soya (25). Sin embargo, otros autores han informado incrementos en la longitud de la raíz en plantas de berro (Lepidium sativum) crecidas en la oscuridad y en raíces primarias de maíz (26).
Se ha planteado que esta respuesta es dependiente de la concentración. Aplicación exógena de bajas concentraciones de BRs (24-epibrasinólida, 0,1 nmol.L-1 y 24-epicastasterona, 10 nmol.L-1) estimularon el crecimiento de la raíz de plantas de genotipos silvestres de Arabidopsis y restablecieron los valores normales en mutantes deficientes de brasinoesteroides (27). Las altas concentraciones de estos compuestos resultaron inhibitorias (10 y 500 nmol.L-1 de EBL y ECS, respectivamente) probablemente por estimulación de la biosíntesis de etileno, el cual es bien conocido que inhibe la elongación de las raíces (28). En este trabajo se utilizó 100 nmol.L-1 de EBL, por lo tanto es posible que ocurra algo similar a lo informado en la referencia anterior. ]]>
El compuesto Brz2001 se ha caracterizado como un inhibidor específico de la biosíntesis de los BRs (9). La capacidad de reversión del efecto inhibitorio por la EBL, sugiere que los BRs pueden contrarrestar, parcialmente la deficiencia de la hormona endógena. Este resultado coincide con hallazgos previos obtenidos en nuestro laboratorio (29) pero, contrario a previas observaciones, no se detectó un restablecimiento parcial del crecimiento por la EBL (Figura 2A). Esto pudiera deberse a las diferencias estructurales ya explicadas, así como a la concentración empleada en los ensayos.
Por otra parte, el BB-16 incrementó la longitud de la raíz de las plántulas de la variedad Amalia, con el control (agua), y este mismo comportamiento se evidenció cuando las plántulas crecieron en medio salino, lo que sugiere que el BB-16 puede inducir la activación de mecanismos de tolerancia al NaCl en las raíces.
En la Figura 3 se muestran las masas fresca y seca de las plántulas de ambas variedades de tomate. Como se puede observar existe una afectación en la masa fresca de las plántulas de la variedad Amalia crecida en medio salino, mientras que la de la var. INCA 9(1) no tuvo diferencias significativas. En cuanto a la masa seca sucedió todo lo contrario, mostrándose la afectación en las plántulas de la variedad INCA 9(1). Por otra parte, no se apreció un efecto protector de los brasinoesteroides utilizados en la masa seca de las plántulas etioladas de ambas variedades sometidas a estrés salino (Figura 3). Los efectos del inhibidor Brz2001 a la concentración empleada se mostraron en la masa fresca de las plántulas de la variedad Amalia y en la masa seca de las plántulas de ambas variedades.
La adición de Brz2001 a cultivos de Wolfia arrhiza inhibió su crecimiento después de siete días de cultivo. La inhibición del crecimiento pudo ser revertida parcialmente por la adición de la 24-epibrasinólida, pero no se recuperaron los niveles del control, especialmente a las concentraciones de Brz2001 entre 5×10-5-10-4 mol.L-1 (30). En este trabajo la EBL y el BB-16 revirtieron la inhibición inducida por el Brz2001 en la masa seca de la var. Amalia.
Otros autores reportaron que el análogo BB-6 a la concentración de 0,001 mg.L-1 estimuló las masas fresca y seca de las radículas de plántulas de tomate var. Campbell-28 crecidas en NaCl (50 y 100 mmol.L-1)(8). ]]>
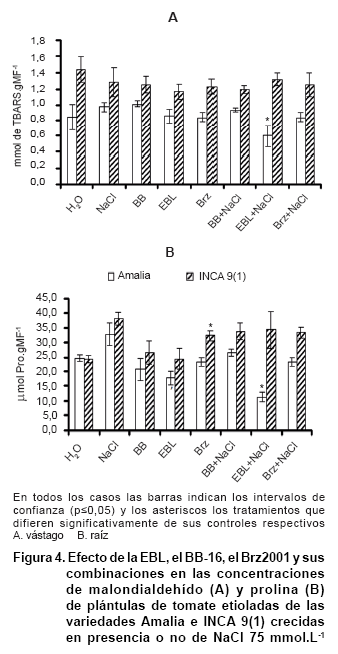
En cuanto a los indicadores bioquímicos evaluados en las plántulas de tomate etioladas, es de destacar que la salinidad no provocó un incremento en la concentración de malondialdehido (MDA), indicador de peroxidación lipídica de las membranas provocada por el daño oxidativo. Los niveles de MDA se mantuvieron menores en las plántulas estresadas de la variedad Amalia tratadas con EBL (Figura 4A); sin embargo, este comportamiento no influyó en la respuesta del crecimiento de esta variedad ante el estrés salino.
El incremento de los niveles de MDA ante estrés salino ha sido informado por otros autores pero en raíces de variedades susceptibles y ha sido correlacionado con el decremento en las actividades de enzimas antioxidantes como catalasa y glutationa reductasa y una actividad basal de superóxido dismutasa y ascorbato peroxidasa. Sin embargo, en variedades tolerantes ha sucedido todo lo contrario (31, 32).
El tratamiento de posturas de Robinia pseudoacacia con brasinólida (BL) disminuyó el contenido de malondialdehído, lo cual fue asociado con una mayor tolerancia al déficit hídrico (33) . En cuanto al estrés salino se ha observado una disminución de la peroxidación lipídica en plantas de frijol, maíz y mostaza tratadas con diferentes concentraciones de brasinoesteroides , fundamentalmente 24-epibrasinólida y homobrasinólida (34-36).
Los niveles de prolina aumentaron con el estrés salino en la variedad INCA 9(1). Sin embargo, la EBL en presencia de NaCl en la variedad Amalia disminuyó la prolina en comparación con el control con NaCl. Un comportamiento diferente mostró la var. INCA 9(1), donde los niveles se mantienen iguales entre las plántulas estresadas tratadas con EBL y las control (Figura 4B). Sin embargo, aunque se observa esta respuesta en la variedad Amalia, que pudiera estar asociada a un posible efecto protector de la prolina, este no influyó en el crecimiento.
Es interesante destacar que en el tratamiento con Brz solo aumentaron los niveles de prolina con respecto al control. Esto pudiera sugerir que la deficiencia de hormona endógena incrementa la biosíntesis de la prolina, lo cual pudiera corroborar la hipótesis de que los brasinoesteroides modulan negativamente la biosíntesis de este aminoácido (2). ]]>
Las plantas superiores pueden acumular prolina en respuesta a diversos estrés abióticos como sequía y salinidad, bajas temperaturas o radiación ultravioleta (10). Aunque la acumulación de prolina ha sido observada en diversos cultivos en condiciones de estrés salino, su rol exacto es muy controversial, pues es considerada más un síntoma de daño que un indicador de tolerancia al estrés. La acumulación de prolina ha sido correlacionada con el grado de deterioro inducido por el estrés y hay autores que plantean que el aumento de este aminoácido podría deberse a una necesidad de la planta de eliminar el exceso de amonio (10).
Por otra parte, la prolina es sintetizada del glutamato o la ornitina, y la primera es considerada la vía predominante en el estrés salino, ya que en estas condiciones disminuyen las actividades de la alanina y aspartato aminotransferasas acumulándose glutamato (37). En esta vía, la biosíntesis de la prolina comienza con la fosforilación y reducción del glutamato a glutamil-5-semialdehído (G5SA) por la enzima bifuncional-pirrolina-5-carboxilato sintasa (P5CS). La actividad Υ-glutamil quinasa de P5CS representa el paso limitante de la velocidad de reacción de esta vía y es sensible a retroinhibición por Pro (38). La expresión de P5CS fue fuertemente inducida por el estrés salino. Esto sugiere que P5CS en la biosíntesis de la prolina juega un papel protector bajo las condiciones de estrés, quizás controlando el potencial redox y estimulando la vía de las pentosas fosfato para la regeneración del NADP+. Algunos autores sugirieron que la acumulación de prolina inducida por el estrés salino puede ser causada por la expresión de P5CS (39).
Por otro lado, en Arabidopsis thaliana, la sequía y el estrés salino activan diferencialmente la expresión de dos genes de P5CS: AtP5CS1 (At2g39800) y AtP5CS2 (At3g55610). AtP5CS1 es responsable de la acumulación de prolina durante el estrés salino y la sequía. La expresión de AtP5CS1 es activada por una vía de traducción de señales dependiente de ABA y modulada por la luz y los brasinoesteroides (40). Esto pudiera sugerir un posible mecanismo por el cual los brasinoesteroides disminuyen los efectos adversos provocados por la salinidad. Sin embargo, esto no se evidenció en este trabajo.
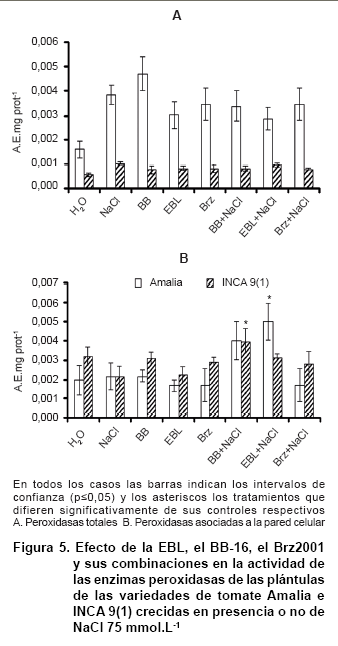
En cuanto a los niveles de actividad de las enzimas peroxidasas totales, es la variedad Amalia la que tiene una mejor respuesta. Las plántulas sometidas al estrés salino tienen niveles superiores a las plántulas crecidas en agua, y aunque en el resto de los tratamientos se aprecia un incremento, los niveles alcanzados no superan a los controles (Figura 5A). En cuanto a las peroxidasas unidas iónicamente a la pared celular se observa un incremento en las plántulas estresadas tratadas con ambos brasinoesteroides (Figura 5B).
Las peroxidasas de la clase III son óxido-reductasas específicas de plantas, que están implicadas en varios procesos fisiológicos como la desintoxicación del peróxido de hidrógeno, el catabolismo de las auxinas, la lignificación, la suberización, la respuesta a estrés y la senescencia (41). ]]>
El estrés salino aumenta la actividad de la enzima peroxidasa en plantas de arroz (32). Por otra parte, en Brassica juncea, la aspersión foliar de plantas con EBL provocó que las plantas sometidas a estrés salino mostraran una disminución de la peroxidación lipídica, un incremento de la actividad de la CAT; sin embargo, no hubo modificaciones en la actividad POX en comparación con las plantas controles (36).
Se ha encontrado un incremento dependiente de la concentración de NaCl de las peroxidasas PER22 (At2g38380) y PER23 (At2g38390), dos enzimas pertenecientes a las peroxidasas de la clase III, en raíces de Arabidopsis (42).
Se ha postulado que las peroxidasas apoplásticas tienen dos funciones opuestas en la remodelación de la pared celular. Por un lado, permiten el entrecruzamiento de los componentes de la pared celular logrando la rigidez y por otro, están involucradas en la generación de radicales hidroxilo, los cuales pueden romper los polisacáridos de la pared celular permitiendo el ablandamiento de la misma. Se ha observado que tiene un papel importante en la germinación y el crecimiento de plántulas de berro (43). Se ha informado que el aumento de la rigidez de la pared celular, la formación de una barrera física y el proceso de entrecruzamiento de diversos compuestos mediados por las peroxidasas de la clase III puede proteger a las raíces de las plantas de la deshidratación provocada por el déficit hídrico (44).
Además de que actúan como antioxidantes, las peroxidasas pueden estar involucradas en la producción de especies reactivas de oxigeno. Esto puede deberse a que las peroxidasas de la clase III, bajo ciertas condiciones fisiológicas, producen los radicales superóxido e hidroxilo (45).
De forma general, no se encontraron trabajos publicados sobre los efectos del inhibidor de la biosíntesis de brasinoesteroides Brz2001 en el contenido de MDA y prolina, ni en la actividad de las enzimas peroxidasas por lo que estos resultados constituyen los primeros pasos en el estudio de los mecanismos de acción de los brasinoesteroides en la inducción de tolerancia al estrés salino alterando los niveles endógenos de estas hormonas. ]]>
De estos resultados se puede concluir que el inhibidor de la biosíntesis de BRs, Brz2001, fue más potente que el NaCl en retrasar la germinación de las semillas y en reducir la elongación del tallo de las plántulas de tomate de las variedades Amalia e INCA 9(1); sin embargo, no alteró la acumulación de la masa seca de las plántulas. Los resultados sugirieron que la inhibición del crecimiento inducido por la salinidad y por el déficit de hormona endógena transcurren por mecanismos distintos. Además, se ratificó la importancia del genotipo en la capacidad de influir en la respuesta al estrés salino.
REFERENCIAS
1. Yang, C. J.; Zhang, C.; Lu, Y. N.; Jin, J. Q. y Wang, X. L. The mechanisms of Brassinosteroids’ action: from Signal Transduction to plant development. Molecular Plant, 2011, vol. 4, no. 4, p. 588-600.
2. Bajguz, A. y Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiology and Biochemistry, 2009, vol. 47, no. 3, p. 1-8.
3. Kagale, S.; Divi, U. K.; Krochko, J. E.; Keller, W. A. y Krishna, P. Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta, 2007, vol. 225, p. 353-364.
4. Núñez, M.; Mazorra, L. M.; Martínez, L.; González, M. C. y Robaina, C. Análogos de brasinoesteroides revierten parcialmente el impacto del estrés salino en el crecimiento inicial de las plántulas de dos genotipos de arroz (Oryza sativa I.). Cultivos Tropicales, 2007, vol. 28, vol. 2, p. 95-99. ]]>
5. Ogweno, J. O.; Song, X. S.; Shi, K.; Hu, W. H.; Mao, W. H.; Zhou, Y. H.; Yu, J. Q. y Nogués, S. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J. Plant Growth Regul., 2008, vol. 27, p. 49-57.
6. Hayat, S.; Ali, B.; Hasan, S. A. y Ahmad, A. Brassinosteroid enhanced the level of antioxidants under cadmium stress in Brassica juncea. Environmental and Experimental Botany, 2007, no. 60, p. 33-41.
7. Alam, M. M.; Hayat, S.; Ali, B. y Ahmad, A. Effect of 28-homobrassinolide treatment on nickel toxicity in Brassica juncea. Photosynthetica, 2007, vol. 45, p. 139-142.
8. Nuñez, M. Análogos de brasinoesteroides cubanos como biorreguladores en la agricultura. Informe final de Proyectos de Investigaciones. Código 00300047. PNCT. Biotecnología Agrícola. CITMA. La Habana, junio, 2000.
9. Sekimata, K.; Kimura, T.; Kaneko, I.; Nakano, T.; Yoneyama, K.; Takeuchi, Y.; Yoshida, S. y Asami, T. A specific brassinosteroid biosynthesis inhibitor, Brz2001: evaluation of its effects on Arabidopsis, cress, tobacco, and rice. Planta, 2001, vol. 213, p. 716-721.
10. Munns, R. y Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol., 2008, vol. 59, p. 651-681.
11. Hodges, M. D.; DeLong, J. M.; Forney, C. F. y Prange, R. K. Improving the thiobarbituric acid-reactive-substances assay for lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 1999, vol. 207, p. 604-611.
12. Bergmeyer, H. U. Peroxidase. En: Methods of Enzymatic Analysis. New York and London: Academic Press Inc. 1974. p. 685-690.
13. Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 1976, vol. 72, p. 248-254.
14. Lin, C. C. y Kao, C. H. Cell wall peroxidase against ferulic acid, lignin, and NaCl-reduced root growth of rice seedlings. Journal of Plant Physiology, 2001, vol. 158, p. 667-671.
15. Bates, L. S.; Waldren, R. P. y Teare, I. D. Rapid determination of free proline for water stress studies. Plant Soil, 1973, vol. 39, p. 205-207.
16. Rao, S. S. R.; Vardhini, B. V.; Sujatha, E.; Anuradha, S. Brassinosteroids - A new class of phytohormones. Current Science, 2002, vol. 82, no. 10, p. 1239-1245.
17. Steber, C. M. y McCourt, P. A Role for Brassinosteroids in Germination in Arabidopsis. Plant Physiology, 2001, vol. 125, p. 763-769.
18. Nomura, T.; Ueno, M.; Yamada, Y.; Takatsuto, S.; Takeuchi, Y. y Yokota, T. Roles of brassinosteroids and related mRNAs in pea seed growth and germination. Plant Physiology, 2007, vol. 143, p. 1680-1688.
19. Sharma, P. y Bhardwaj, R. Effect of 24-epibrassinolide on seed germination, seedling growth and heavy metal uptake in Brassica juncea L. Gen. Appl. Plant Physiology, 2007, vol. 33, no. 1-2, p. 59-73.
20. Reyes, Y.; Mazorra, L. M.; Martínez, L. y Núñez, M. Efecto del análogo de brasinoesteroide (Biobras-16) en la germinación y el crecimiento inicial de las plantas de dos variedades de tomate en condiciones de estres salino. Cultivos Tropicales, 2010, vol. 31, no. 3, p. 82-88.
21. Tanaka, K.; Nakamura, Y.; Asami, T.; Yoshida, S.; Matsuo, T. y Okamoto, S. Physiological roles of brassinosteroids in early growth of Arabidopsis: Brassinosteroids have a synergistic relationship with gibberellin as well as auxin in light-grown hypocotyl elongation. J. Plant Growth Regul., 2003, vol. 22, p. 259-271.
22. Song, L.; Zhou, X. Y.; Li, L.; Xue, L. J.; Yang, X. y Xue, H. W. Genome-wide analysis revealed the complex regulatory network of brassinosteroid effects in photomorphogenesis. Molecular Plant, 2009, vol. 2, no. 4, p. 755-772.
23. Nomura, T.; Kushiro, T.; Yokota, T.; Kamiya, Y.; Bishop, G. J. y Yamaguchi, S. The last reaction producing brassinolide is catalyzed by cytochrome P-450s, CYP85A3 in tomato and CYP85A2 in Arabidopsis. Journal of Biological Chemistry, 2005, vol. 280, no. 18, p. 17873-17879.
24. Symons, G. M.; Ross, J. J.; Jager, C. E. y Reid, J. B. Brassinosteroid transport. Journal of Experimental Botany, 2008, vol. 59, no. 1, p. 17-24.
25. Sasse, J. M. Physiological actions of brassinosteroids: an update. Plant Growth Regulation, 2003, vol. 22, p. 276-288.
26. Kim, S. K.; Chang, S. C.; Lee, E. J.; Chung, W. S.; Kim, Y. S.; Hwang, S. y Lee, J. S. Involvement of brassinosteroids in the gravitropic response of primary root of maize. Plant Physiology, 2000, vol. 123, p. 997-1004.
27. Müssig, C., Shin, G. H. y Altmann, T., Brassinosteroids promote root growth in Arabidopsis. Plant Physiology, 2003. vol. 133, p. 1261-1271.
28. Divi, U. K.; Rahman, T. y Krishna, P. Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways. BMC Plant Biology, 2010, vol. 10, 151 p.
29. Mazorra, L. M.; Núñez, M.; Nápoles, M. C.; Yoshida, S.; Robaina, C.; Coll, F. y Asami, T. Effects of structural analogs of brassinosteroids on the recovery of growth inhibition by a specific brassinosteriod biosynthesis inhibitor. Plant Growth Regulation, 2004, vol. 44, no. 2, p. 183-185.
30. Bajguz, A. y Asami, T. Suppression of Wolffia arrhiza growth by brassinazole, an inhibitor of brassinosteroid biosynthesis and its restoration by endogenous 24-epibrassinolide. Phytochemistry, 2005, vol. 66, no. 15, p. 1787-1796.
31. Demiral, T. y Turkan, I. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environmental and Experimental Botany, 2005, vol. 53, p. 247-257.
32. Khan, M. H. y Panda, S. K. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiol. Plant, 2008, vol. 30, p. 81-89.
33. Li, K. R.; Wang, H. H.; Wang, Q. J. y Fan, J. Effects of brassinolide on the survival, growth and drought resistance of Robinia pseudoacacia seedlings under water stress. New Forests, 2008, vol. 35, p. 255-266.
34. Rady, M. M. Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress. Scientia Horticulturae, 2011, vol. 129, p. 232-237.
35. Arora, N.; Bhardwaj, R.; Sharma, P. y Arora, H. K. 28-homobrassinolide alleviates oxidative stress in salt treated maize (Zea mays L.) plants. Brazilian. Journal of Plant Physiology, 2008, vol. 20, p. 153-157.
36. Ali, B.; Hayat, S.; Fariduddin, Q. y Ahmad, A. 24-Epibrassinolide protects against the stress generated by salinity and nickel in Brassica juncea. Chemosphere, 2008, vol. 72, p. 1387-1392.
37. Jaleel, C. A.; Manivannan, P.; Lakshmanan, G. M. A.; Sridharan, R. y Panneerselvam, R. NaCl as physiological modulator of proline metabolism and antioxidant potential in Phyllant husamarus. C. R. Biologies, 2007, vol. 330, p. 806.
38. Dea-Wook, K.; Junko, S.; Ganesh, K. A.; Shinsuke, F.; Hitoshi, I.; Du Hyun, K.; Ie-Sung, S. y Randeep, R. Gene transcription in the leaves of rice undergoing salt-induced morphological changes (Oryza sativa L.). Mol. Cells, 2007, vol. 24, no. 1, p. 45-59.
39. Kant, S.; Kant, P.; Raveh, E. y Barak, S. Evidence that differential gene expression between the halophyte Thellungiella halophila, and Arabidopsis thaliana is responsible for higher levels of the compatible osmolyte proline and tight control of Na+ uptake in T. halophila. Plant Cell Environ., 2006, vol. 29, p. 1220-1234.
40. Nounjan, N. y Theerakulpisut, P. Effects of exogenous proline and trehalose on physiological responses in rice seedlings during salt-stress and after recovery. Plant Soil Environ., 2012, vol. 58, no. 7, p. 309-315.
41. Almagro, L.; Gómez Ros, L. V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A. y Pedreño, M. A. Class III peroxidases in plant defence reactions. Journal of Experimental Botany, 2009, vol. 60, no. 2, p. 377-390.
42. Jiang, Y.; Yang, B.; Harris, N. S. y Deyholos, M. K. Comparative proteomic analysis of NaCl stress-responsive proteins in Arabidopsis roots. Journal of Experimental Botany, 2007, vol. 58, no. 13, p. 3591-3607.
43. Muller, K.; Linkies, A.; Vreeburg, R. A. M.; Fry, S. C.; Krieger-Liszkay, A. y Leubner-Metzger, G. In vivo cell wall loosening by hydroxyl radicals during cress (Lepidium sativum L.) seed germination and elongation growth. Plant Physiology, 2009, vol. 150, p. 1855-1865.
44. Gong, P.; Zhang, J.; Li, H.; Yang, C.; Zhang, C.; Zhang, X.; Khurram, Z.; Zhang, Y.; Wang, T.; Fei, Z. y Ye, Z. Transcriptional profiles of drought-responsive genes in modulating transcription signal transduction, and biochemical pathways in tomato. Journal of Experimental Botany, 2010, vol. 61, no. 13, p. 3563-3575.
45. Minibayeva, F.; Kolesnikov, O.; Chasov, A.; Beckett, R. P.; Luthje, S.; Vylegzhanina, N.; Buck, F. y Bottger, M. Wound-induced apoplastic peroxidase activities: their roles in the production and detoxification of reactive oxygen species. Plant, Cell and Environment, 2009, vol. 32, p. 497-508.
Recibido: 13 de abril de 2012 Aceptado: 20 de febrero de 2013
Lic.Yanelis Reyes, Aspirante a Investigador, departamento de Fisiología y Bioquímica Vegetal. Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, La Habana, CP 32700. Email: yanelisrg@inca.edu.cu
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}