Dinámica de la humedad del suelo y la fitomasa de raíces en ecosistemas de la Sierra del Rosario, Cuba
Dynamics of soil moisture and root phytomass in ecosystems of Sierra del Rosario, Cuba
L. Hernández y J. A. Sánchez
Instituto de Ecología y Sistemática, Ministerio de Ciencia, Tecnología y Medio Ambiente, Carretera de Varona Km 3 ½ Capdevila, Boyeros, C.P. 11900, La Habana 19, Cuba
E-mail: wicho@ecologia.cu ]]>
Resumen
Se estudió la dinámica de la humedad del suelo y la fitomasa de raíces finas (< 1,0 mm) en siete ecosistemas, con diferentes condiciones de suelo y tipo de vegetación, de la Reserva de la Biosfera Sierra del Rosario, Cuba. Cinco sitios provenían de bosques tropicales siempreverdes mesófilos (vegetación predominante en la reserva), otro perteneció a un bosque tropical siempreverde micrófilo y también se incluyó una sabana arbustiva. La humedad del suelo se determinó cada semana durante un año, a 0-5 y 5-15 cm de profundidad; cada semana se colectaron cinco muestras por localidad. Para la determinación de la fitomasa de raíces se muestrearon cuatro meses del año en estudio (dos en la estación poco lluviosa y dos en la lluviosa), en los mismos puntos donde se estudió la humedad del suelo. Esta última varió de acuerdo con la localidad y fue menor en los sitios secos (bosque micrófilo y sabana); entre los bosques mesófilos estas diferencias fueron menos pronunciadas. En general, la capa de suelo 0-5 cm, en todas las localidades, mantuvo una mayor humedad y producción de raíces finas que la capa de 5-15 cm. La fitomasa de raíces cambió con las estaciones climáticas y los valores más altos se hallaron en el bosque micrófilo, donde la humedad del suelo es menor. Los resultados demostraron la importancia de la primera capa del suelo en la dinámica de la humedad del sustrato y de la fitomasa de raíces finas.
Palabras clave: bosque tropical, contenido de agua en el suelo, pastizales, raíces.
Abstract
The dynamics of soil moisture and fine root phytomass (< 1,0 mm) was studied in seven ecosystems, with different soil conditions and vegetation types, of the Sierra del Rosario Biosphere Reserve, Cuba. Five sites were from tropical evergreen mesophyllous forests (prevailing vegetation in the reserve), another one belonged to a tropical evergreen microphyllous forest and a shrubby savanna was also included. Soil moisture was determined to a depth of 0-5 cm, and 5-15 cm every week during a year. Each week five samples were collected per site. For the determination of the root phytomass samplings were made in four months of the year under study (two in the dry season and two in the rainy season), in the same spots where soil moisture was studied. This last variable varied according to the site, and it was lower in the dry sites (i.e., microphyllous forest and savanna); among evergreen forests these differences were less pronounced. In general, the 0-5cm soil layer, in all sites, maintained higher humidity and fine root production than the 5-15 cm layer. Root phytomass changed with season, and the highest values were found in the mycrophyllous forest where soil moisture is lower. The results showed the importance of the first soil layer in the dynamics of soil moisture and fine root phytomass.
Key words: tropical forest, soil water content, pasturelands, roots.
]]> INTRODUCCIÓN
El agua en forma de lluvia es un recurso vital para el desarrollo de la vegetación en la mayor parte del mundo. Las características principales del agua de lluvia, en cuanto a su utilidad, son: su cantidad, su frecuencia y su intensidad, cuyos valores varían considerablemente de un lugar a otro (Bucci et al., 2004; Hernández y Sánchez, 2009). En Cuba, un país subtropical, la estacionalidad de las lluvias es muy marcada y tienen gran variabilidad los factores antes mencionados. También las lluvias varían de acuerdo con los años, debido al paso de los ciclones y los frentes fríos.
Estos aspectos climáticos determinan el tipo de formación vegetal que caracteriza un sitio, pero el suelo también influye significativamente en esta distribución. De hecho, se conoce que los cambios hidrológicos proporcionan el desarrollo de diferentes comunidades vegetales, que están estrechamente relacionadas con las variaciones drásticas de los tipos de suelo (Herrera, 2007). Además, la interacción planta-suelo que se establece en las formaciones boscosas difiere considerablemente de la que ocurre en las formaciones herbáceas, lo que implica que estas comunidades vegetales tengan características edafoclimáticas bien distintas (Bazzaz, 1996).
Para la región occidental de Cuba, existe una base teórico-práctica de la ecología de los bosques húmedos y secos de la Reserva de la Biosfera Sierra del Rosario (RBSR), donde aparece información del clima, el suelo, la vegetación y los eventos ecofisiológicos de regeneración y crecimiento de las plantas de la región (Sagué y Hernández, 1978; Herrera et al., 1988a; Herrera et al., 1997; Mancina et al., 2007; Hernández y Sánchez, 2009; Sánchez et al., 2009). También en la RBSR se ha estudiado la humedad del suelo y su relación con la producción de fitomasa subterránea de las comunidades herbáceas (Sagué y Hernández, 1978; Herrera et al., 1988b); sin embargo, estos estudios aún son muy escasos para las formaciones vegetales de la RBSR y para otras regiones submontanas y montañosas de Cuba (Hernández et al., 2006; Hernández y Rodríguez, 2011). En general, a nivel mundial también son muy limitados los datos publicados sobre la producción y renovación de las raíces finas en los bosques tropicales de montaña, aunque se conoce que el tamaño, la estructura y la dinámica del sistema radical de los árboles son factores importantes para la estabilidad de la pendiente y la retención de los nutrientes del suelo (Hertel et al., 2003). Esto evidencia la necesidad de conocer la estructura y el funcionamiento del sistema radical de los bosques tropicales, para proteger y manejar adecuadamente dichos ecosistemas, que están entre los más vulnerables de la Tierra (FAO, 2011).
Por su parte, las grandes diferencias en la cantidad y la actividad de las raíces finas en los sistemas montañosos (Priess et al., 1999; Hertel et al., 2003) pueden ser el resultado de las diferencias en la composición de las especies, los tipos de suelo, la elevación del sitio, la humedad del suelo y el impacto humano (Hertel et al., 2003), tal como ha sido informado para distintas vegetaciones herbáceas cubanas; pero este aspecto aún está por definir para la vegetación boscosa de la llanura y de la montaña de la RBSR.
Por lo tanto, el objetivo de este estudio fue caracterizar la dinámica de la humedad del suelo y la producción de raíces finas (< 1,0 mm) en siete ecosistemas de la RBSR, que representan distintas formaciones boscosas y una sabana típica de la región.
MATERIALES Y MÉTODOS
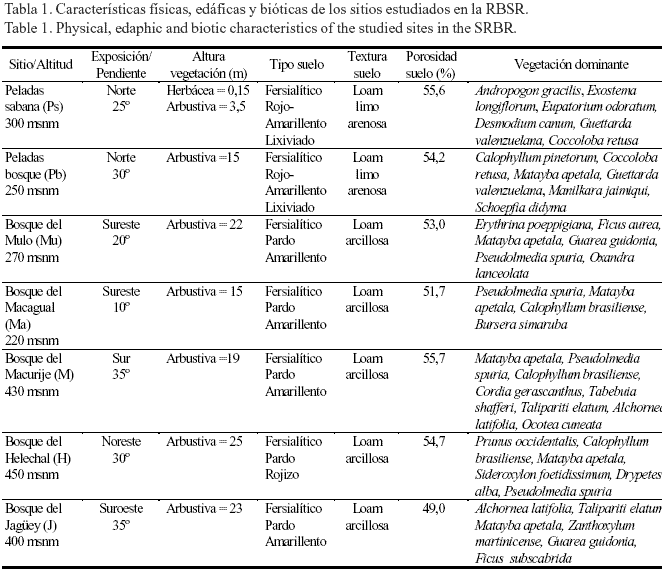
Descripción general de sitio. La RBSR se localiza en la parte más oriental de la Sierra del Rosario, Artemisa, Cuba (22º 45' N, 82º 50' O) y tiene una extensión de 25 000 ha. El sistema montañoso está fuertemente diseccionado y las alturas varían entre 100 y 565 m. La precipitación total durante el año de estudio (abril/1987- marzo/1988) fue de 1 631 mm (abril, mayo y diciembre fueron los meses más secos y las máximas lluvias cayeron durante agosto, septiembre y octubre); este año puede considerarse muy seco, dado que en la RBSR la precitación total anual promedio es de 2 013 mm, con una pronunciada estación seca desde diciembre hasta marzo (Herrera et al., 1997; Hernández y Sánchez, 2009). La temperatura media anual del aire fue de 24,2ºC. La flora, la vegetación y la geología de la RBSR se describieron ampliamente por Herrera et al. (1988a). Los suelos de la reserva son variados y según Hernández et al. (1988) predominan los Fersialíticos y los Pardos. La vegetación más extendida es el bosque tropical siempreverde mesófilo, con árboles de hasta 25 m de altura (Herrera et al., 1997).
Humedad del suelo y fitomasa de las raíces. Para determinar la humedad del suelo y la fitomasa de las raíces finas, se seleccionaron siete localidades representativas de la vegetación de la RBSR (tabla 1), entre ellas cinco localidades de bosque siempreverde mesófilo: dos en las subcuencas del sitio del Mulo (Mu) y el Macagual (Ma) y tres en localidades del Salón: Macurije (M), Helechal (H) y Jagüey (J). En la subcuenca del sitio conocido como Las Peladas se escogieron dos áreas: una perteneciente a un bosque siempreverde micrófilo (Pb) y otra a una sabana arbustiva (Ps). En todos los ecosistemas se escogió una parcela de 20 x 20 m, representativa de la comunidad vegetal. La humedad del suelo (base seca) se determinó durante un año, a 0-5 y 5-15 cm de profundidad; cada semana se colectaron cinco muestras con un cilindro de 5 cm de diámetro. Para determinar la fitomasa de las raíces finas (<1,0 mm) se muestreó en cuatro meses, dos en la estación seca (noviembre del 1987 y febrero del 1988) y dos en la estación lluviosa (mayo y julio del 1987), en los mismos puntos donde se tomaron las muestras para la determinación de la humedad del suelo. Las raíces se procesaron acorde con lo informado por Hernández (1999). La tasa de renovación (TR) de la fitomasa de las raíces finas se estimó según Singh y Singh (1981) para cada ecosistema, como la diferencia entre el valor máximo y el valor mínimo de la fitomasa, dentro de cada profundidad de suelo, dividido entre el valor promedio de la fitomasa subterránea. ]]>
También en los sitios de colecta de las raíces se tomaron muestras de suelo para el análisis físico, que se realizó en coordinación con el laboratorio de Química del Instituto de Ecología y Sistemática y con el laboratorio de Física del Instituto de Suelos, La Habana, Cuba. La fitomasa del mantillo (hojarasca y esteras de raíces) en los sitios boscosos estudiados fue muy variable: la hojarasca presentó valores entre 478 y 945 gm-2, y las esteras de raíces variaron desde 27,1 hasta 99,4 gm-2 (Hernández et al., 2006).Análisis de los datos. La normalidad de los datos se verificó mediante la prueba de bondad de ajuste Kolmogorov-Smirnov y la homogeneidad de varianza a través de la prueba de Bartlett. Las variables porcentaje de humedad del suelo y de la biomasa subterránea total de raíces finas se procesaron mediante análisis de varianza de clasificación simple, con arreglo factorial de los tratamientos (localidad por profundidad por mes). No se aplicaron pruebas de comparación múltiple de medias a posteriori, debido a que las variables independientes representaban combinaciones de tratamientos cualitativos no estructurados (localidad y mes) con tratamiento cuantitativo (profundidad) (Blanco, 2001). Para detectar posibles agrupamientos entre localidades, de acuerdo con la humedad del suelo o la fitomasa subterránea, se usó el método de conglomerados (o análisis de cluster jerárquico). En estos análisis se utilizaron los datos originales de humedad del suelo, por profundidad y por meses estudiados; en el caso de la fitomasa se hizo un procesamiento similar, pero además se realizó una matriz con la información de la fitomasa total de raíces obtenidas en todas las profundidades. El dendrograma se construyó a partir de una matriz de distancia euclidiana y por el método de enlace de ligamiento promedio no ponderado (UPGMA). En este último análisis, mientras menor sea la distancia entre las unidades (localidades) que se analizan, mayor será su similitud; es decir, donde la distancia tiene valor 0, indica máxima similitud entre las localidades. La relación entre la fitomasa total de las raíces y la humedad del suelo, obtenida por localidad, se exploró mediante el coeficiente de correlación lineal de Pearson.
RESULTADOS Y DISCUSIÓN
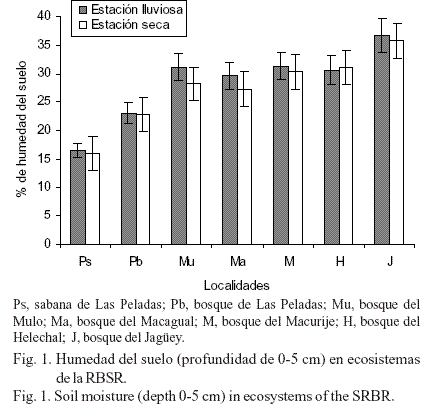
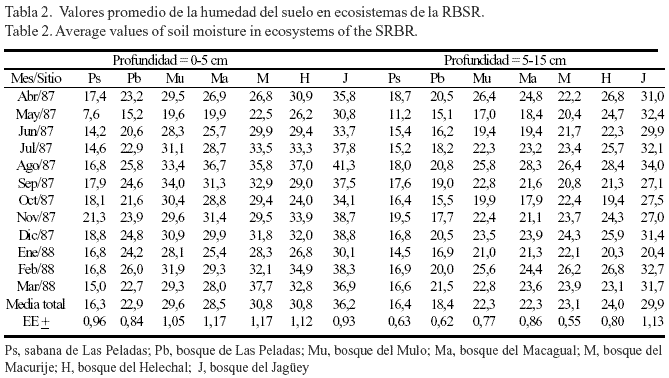
Humedad del suelo. La humedad del suelo cambió considerablemente con la profundidad del sustrato, la localidad y los meses del año (tabla 2), y la interacción de estos tres factores también afectó significativamente (F= 7,58; P< 0,001) el comportamiento de esta variable. La menor humedad del suelo se obtuvo en la sabana de Las Peladas, durante el mes de mayo, en la profundidad de 0-5 cm (tabla 2); en esta misma profundidad se alcanzaron los máximos valores de humedad en el suelo del bosque del Jagüey, en agosto, que coincidió con el inicio del período más lluvioso durante el presente estudio. Por otra parte, independientemente de la localidad, la humedad del suelo fue siempre superior en 0-5 cm con relación a los valores alcanzados en 5-15 cm, excepto para la localidad de la sabana de Las Peladas, donde no existieron diferencias.
Cuando se analizó esta variable por estación hidrológica, en la profundidad de 0-5 cm no existieron diferencias significativas entre las estaciones (fig. 1) para ninguno de los ecosistemas, y tampoco en la profundidad de 5-15 cm (datos no mostrados). Este resultado demostró que, en los ecosistemas estudiados, el análisis de la humedad del suelo debe realizarse por meses, en lugar de emplear la estación hidrológica como factor independiente, debido a que la estacionalidad de las lluvias no evidenció la variabilidad sufrida a través del año. Sin embargo, el agrupamiento por estación del año no solapó las diferencias que existieron entre localidades, tal como se demostró en el análisis anterior. De hecho, las localidades boscosas del Mulo, Macagual, Macurije, Helechal y del Jagüey siempre presentaron valores de humedad del suelo significativamente superiores a los obtenidos en la sabana y el bosque de Las Peladas (fig.1), y entre estos últimos sitios también existieron diferencias considerables.
El resultado anterior se corroboró, igualmente, cuando se determinó el agrupamiento de los ecosistemas por los valores de humedad del suelo de todo el año y por profundidad (fig.2). En el dendrograma para la profundidad de 0-5 cm (fig. 2A), se apreció la formación de dos grandes grupos. El primero estuvo constituido por los sitios boscosos más húmedos dentro de la Sierra del Rosario (Jagüey, Helechal, Macagual, Mulo y Macurije); en este grupo se observaron, además, diferentes grados de similitud entre sus unidades. La localidad del Jagüey presentó los máximos valores de humedad del suelo, con lo que se separó del resto de los sitios antes mencionados. Sin embargo, en este mismo conjunto se agruparon, a menor distancia, los ecosistemas boscosos del Mulo y Macurije, lo que demostró que fueron las localidades más similares de acuerdo con la variable estudiada. El segundo grupo estuvo formado por los sitios de Las Peladas, que presentaron los menores valores de humedad. En la profundidad de 5-15 cm, el dendrograma (fig. 2B) mostró un resultado similar al anterior, pero se evidenciaron más las diferencias entre los sitios húmedos y secos. De hecho, el bosque del Jagüey se separó por completo del resto de los sitios y se unió al mismo nivel de aquellos procedentes de sitios húmedos y secos, aunque los de Las Peladas (los más secos) ocuparon una posición intermedia entre los húmedos.
Los resultados anteriormente mencionados se explican por sí solos, si se tiene en cuenta que la textura del suelo y su estructura pueden influir notablemente en el grado de retención de agua (Kaúrichev, 1980), aunque también otros factores bióticos y abióticos influyen significativamente en la humedad del suelo (Herrera et al., 1988b). En ambas formaciones vegetales del sitio de Las Peladas (sabana y bosque) los suelos presentaron una textura arenosa (tabla 1), lo que sin dudas ofrece una menor resistencia a perder el agua, comparado con los suelos de textura loam arcillosa (mayor capacidad de retención de agua) encontrados en los sitios del Mulo, Macagual, Macurije, Helechal y del Jagüey. También en Las Peladas los suelos tuvieron un mayor porcentaje de porosidad (tabla 1), lo que unido a otras características funcionales de estos ecosistemas (Herrera et al., 1988b) los ubican entre los más tensionados dentro de la RBSR. Según Herrera et al. (1988b), en el bosque del Jagüey sucede lo contrario con relación al bosque micrófilo de Las Peladas, debido a que el primero se sustenta sobre un suelo Fersialítico Pardo Amarillento con una estructura loam arcillosa y con presencia de montmorillonita; ello facilita la humedad casi permanente del suelo y el desarrollo de un bosque con estructura continental y mesófilo, que no pudiera existir en Las Peladas.
Fiala et al. (1991) informaron que en una ladera de un pastizal, al noreste de la RBSR, la humedad del suelo varió de la parte más alta a la más baja (30,3-38,7%, respectivamente), donde, además, las características físicas y químicas de los suelos eran diferentes. Por su parte, Sagué y Hernández (1978) determinaron que este indicador es mayor debajo del bosque que en los pastizales, lo que es más evidente en la capa de suelo de 0-20 cm. Sin embargo, estos autores observaron que en los horizontes más profundos el déficit de humedad es mayor en el bosque que en el pastizal, lo cual pudiera estar dado por la mayor transpiración de la vegetación en el primero que en el segundo, aspectos que se deben tener en cuenta para la repoblación de las áreas de las cuencas con embalse. Por otra parte, Sarmiento y Vera (1976) señalaron que en los bosques y sabanas de Venezuela los regímenes de humedad son muy similares, y el del bosque casi siempre es más seco, probablemente como consecuencia de la mayor utilización de agua por este tipo de vegetación, en comparación con la sabana. ]]>
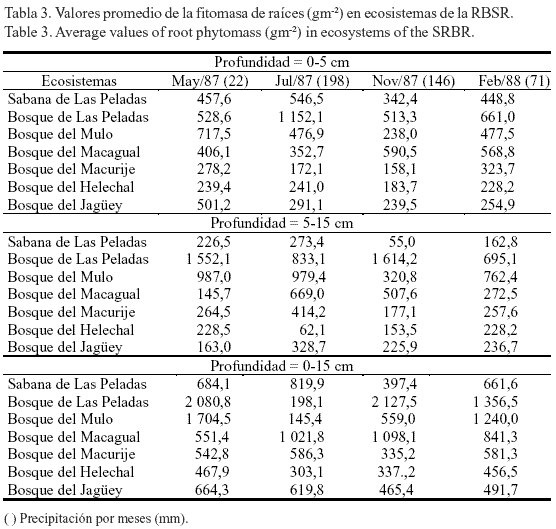
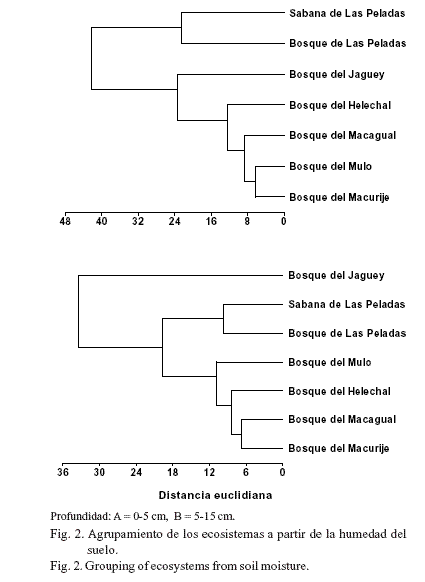
Sagué et al. (1979), en estudios realizados durante diez días en las estaciones poco lluviosa y lluviosa, en áreas del Salón (sitio de la RBSR), observaron que en ambas estaciones la humedad del suelo fue mayor en el área de bosque, seguida de los pastizales y, por último, las terrazas.La raíces en los ecosistemas. La fitomasa de raíces finas cambió considerablemente con la profundidad del sustrato, la localidad y los meses del año (tabla 3), y la interacción de estos tres factores también afectó significativamente (F = 34,6; P < 0,001) el comportamiento de esta variable. El menor valor de raíces finas (55 gm-2) se obtuvo en los suelos de las sabanas de Las Peladas, durante noviembre de 1987, en la profundidad de 5-15 cm (tabla 3); los máximos valores en esta misma profundidad se alcanzaron en el bosque de Las Peladas, en mayo (1 552,1 gm-2) y en noviembre de 1987 (1 614,2 gm-2); este último se correspondió con el período poco lluvioso, aunque en el mes de noviembre la lluvia caída fue alta (146 mm). Por su parte, en el bosque del Mulo aparecieron los máximos valores de biomasa subterránea durante julio de 1987 (mes más lluvioso de los cuatro estudiados) y en febrero. En la profundidad de 0-5 cm la tendencia de la producción de biomasa cambió con relación a la obtenida en 5-15 cm. Los mayores valores se obtuvieron en el bosque de Las Peladas en julio de 1987 y febrero de 1988; en el bosque del Mulo en mayo de 1987, y en el bosque del Macagual en noviembre de 1987. Por último, la producción total de biomasa subterránea (profundidad de 0-15 cm) siguió una tendencia muy similar a la obtenida en la profundidad de 0-5 cm, con la diferencia de que en los meses de mayo y noviembre los mayores valores de raíces finas aparecieron también en el bosque de Las Peladas. Al parecer, las condiciones de estrés hídrico y nutrimental, creadas fundamentalmente por la estructura del suelo (arenoso), indujeron la mayor producción de raíces finas, como vía para minimizar las tensiones ambientales. Ello fue informado para este ecosistema por Herrera et al. (1988b), quienes plantearon además que quizás el bosque de Las Peladas base su funcionamiento en condiciones edáficas estresantes, debido a su gran producción de raíces, la formación de esteras radicales y su elevada esclerofilia y esclerorrizia.
La distribución vertical de las raíces finas tendió a ser mayor en la capa de suelo de 0-5 cm (tabla 3), tal como se ha informado en diferentes comunidades herbáceas y arbóreas (Herrera et al., 1988b; Hernández y Rodríguez, 2011). Sin embargo, esta tendencia no quedó muy clara para algunos sitios estudiados, ni para los bosques de Las Peladas y el Mulo, que tuvieron la máxima producción de raíces finas en la capa de 5-15 cm para los meses de mayo y noviembre; aunque se debe tener en cuenta que esta capa duplica en área y volumen a la primera, lo que posiblemente permita esa producción de raíces (Sagué y Hernández, 1978; Herrera et al., 1988b; Fornaris el al., 2006).
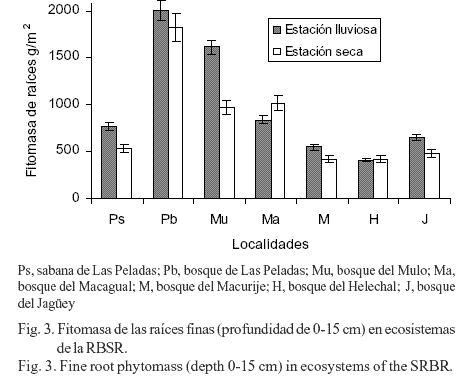
Por su parte, cuando se estudió la producción de fitomasa de raíces por estación hidrológica, en la profundidad de 0-15 cm solo existieron diferencias significativas para las localidades de la sabana de Las Peladas, y los sitios boscosos del Mulo, Macurije y del Jagüey (fig. 3). En el resto de los sitios no hubo diferencias entre las estaciones, pero tendieron a ser mayores en el bosque de Las Peladas durante la estación lluviosa y menor en el bosque del Macagual en esta misma época del año; en el bosque del Helechal los valores promedio de la fitomasa de las raíces no cambiaron con la estación. En general, este
resultado demostró la variabilidad que sufre la fitomasa de las raíces con la disponibilidad de agua. Sin embargo, es importante señalar que en el presente estudio solo se determinó la cantidad de fitomasa de las raíces en dos meses de cada estación, lo que sin duda pudo afectar los resultados, aunque es conocido los cambios que sufre la biomasa de las raíces con la disponibilidad de agua del sustrato (Sagué y Hernández, 1978).
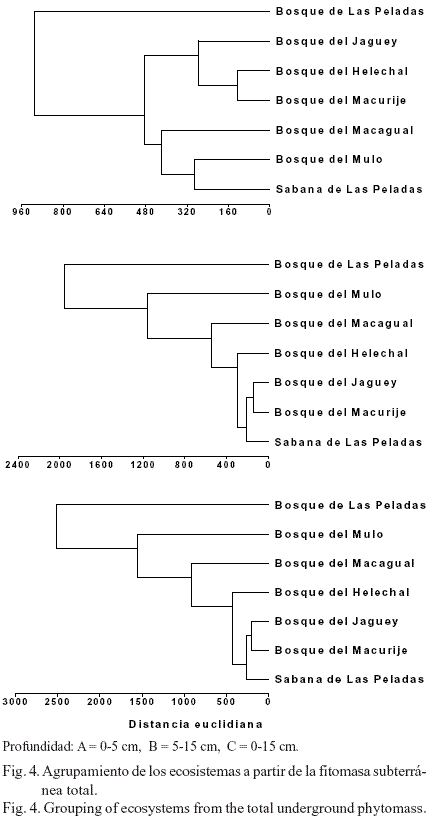
Los dendrogramas realizados con los datos de la fitomasa de las raíces, igualmente mostraron variabilidad entre localidades cuando se analizaron para las distintas profundidades (fig. 4). En la profundidad de 0-5 cm (fig. 4A) surgieron tres agrupaciones. El primer grupo, constituido por el bosque de Las Peladas, presentó los mayores valores de biomasa de las raíces y se separó por completo del resto de las localidades. En el otro extremo de este agrupamiento apareció un segundo grupo constituido por aquellas localidades que tuvieron valores medios (bosques del Macagual y el Mulo, y la sabana de Las Peladas). El tercer agrupamiento estuvo formado por los sitios que presentaron valores bajos (bosques de Jagüey, Helechal y Macurije), y que ocuparon en el dendrograma una posición intermedia entre los sitios de mayor producción de fitomasa de las raíces.
En la profundidad de 5-15 cm, el dendrograma mostró un nuevo agrupamiento de los ecosistemas (fig. 4B). Los sitios con mayor cantidad de fitomasa de las raíces (bosque de Las Peladas y del Mulo) se situaron en un extremo de este agrupamiento y se diferenciaron de aquellos con una producción media o baja (bosques del Macagual, Helechal, Jagüey y Macurije, y sabana de Las Peladas). Finalmente, el análisis total de la producción de la fitomasa de las raíces finas (fig. 4C; profundidad de 0-15 cm) mostró un agrupamiento muy similar al obtenido en 5-15 cm, lo que evidenció que la cantidad de fitomasa subterránea encontrada en esta última profundidad define su distribución espacial, al menos en los primeros 15 cm de profundidad del suelo, para los sitios estudiados.
En los pastizales, las primeras capas de suelo desempeñan un papel fundamental en la distribución vertical de las raíces (Hernández, 1999). La mayor producción de fitomasa subterránea total estuvo concentrada en la capa superficial del suelo (0-5 cm) y representó el 65,0% de este perfil; en la capa 5-15 cm se concentró el 77,9% (1 262 gm-2) de esta fitomasa. Para la capa 0-15 cm el 84% de la fitomasa estuvo concentrada en la capa de suelo 0-5 cm, lo que equivale a 1 060 gm-2. En un pastizal de Cynodon nlemfuensis, Hernández et al. (1998) encontraron, en los primeros 15 cm de suelo (para un perfil de 95 cm de profundidad), el 70% de la fitomasa de las raíces y el 86% de los detritos. En estudios realizados en los bosques del Mulo, en la RBSR, Sagué y Hernández (1978) determinaron que el 63% de las raíces finas se encontraba en la capa de suelo 0-10 cm y hasta los 20 cm tenían valores por encima del 73%. Por su parte Hertel et al. (2003), en bosques de Costa Rica con diferente estatus sucesional, señalaron que la mayor cantidad de raíces finas se concentra en la capa orgánica del suelo y no en los primeros 10 cm de la capa mineral. ]]>
Relación entre la humedad del suelo y la cantidad de fitomasa de raíces por localidades. Existió una correlación significativa y negativa entre la humedad del suelo y la cantidad de fitomasa producida en los sitios boscosos (fig. 5), o sea, al aumentar la humedad del suelo disminuyó la producción de raíces. De este modo, el sitio boscoso más seco (bosque de Las Peladas) tuvo la mayor producción de raíces; por su parte, los bosques más húmedos (del Jagüey y el Helechal) mostraron los valores medios más bajos de fitomasa de las raíces. Ello no ocurrió al incluir el sitio de la sabana de Las Peladas (datos no mostrados), lo que evidencia un comportamiento diferencial en esta relación entre sitios boscosos y sabanas.En los bosques tensionados de la RBSR, como el de Las Peladas, existe una gran cantidad de fitomasa de esteras radicales y la relación raíz/tallo (biomasa subterránea/biomasa aérea) es alta (Herrera et al., 1988b), lo cual apoya el resultado previamente discutido. Por su parte, la fitomasa de las raíces promedio en la capa de materia orgánica (mantillo) en los sitios boscosos, al inicio de la investigación, fue muy variable y presentó un valor medio de 703 ± 93 gm-2 para la hojarasca, y de 49 ± 16 gm-2 para las esteras radicales. Este último fue superior (479 gm-2) en la región minera de Moa, que es un sitio muy tensionado por la falta de nutrientes disponibles y la sequía edáfica (Hernández et al., 2003), lo que a su vez representó el 21% de todo el perfil estudiado. Las áreas de pluvisilva de Moa presentaron los valores más altos de las esteras radicales (34,1% del perfil); el promedio de la fitomasa de las raíces para los diferentes tipos de bosque que se estudiaron (Horizonte O + capa 0-20 cm) fue de 2 275 gm-2, que representó el 54,7% de la materia orgánica del suelo. De acuerdo con lo planteado por Fornaris et al. (2006), la presencia de la estera radical no siempre depende de una alta esclerofilia, y solo se desarrolla cuando hay suficiente humedad ambiental.
En estudios realizados acerca de las raíces finas (< 0,1 mm), en distintas formaciones boscosas y plantaciones de Cuba (Hernández et al., 2006), se observaron diferencias notables en las primeras capas de suelo (0-10 y 0-15 cm). En los manglares este indicador fluctuó entre 5,3 y 8,6 t/ha, en los bosques siempreverdes de 3,4 a 4,7 t/ha y en los semideciduos fue de 2,9 t/ha; los valores bajos (1,8 t/ha) se encontraron en las plantaciones de majagua (Talipariti elatum) y los más bajos (1,0 t/ha) en los pinos (Pinus cubensis y P. caribaea).
Por otra parte, en los pastizales al noreste de la RBSR, Hernández y Fiala (1992) hallaron valores de fitomasa aérea total entre 626 y 1 358 gm-2 y de fitomasa subterránea total entre 1 182 y 1 735 gm-2. Las variaciones fueron ocasionadas principalmente por las diferencias de relieve y pendiente, las cuales determinaron cambios en el contenido de humedad y nutrientes del suelo. En la parte más baja de la ladera las condiciones favorecieron los mayores valores de fitomasa aérea viva (649 gm-2) y de rizomas (1 023 gm-2), y la menor cantidad de raíces vivas. A través de la pendiente se produjo un incremento en la proporción total de fitomasa aérea (de 34,1 a 43,9%), mientras que la fitomasa subterránea disminuyó (de 65,9 a 56,1%) de la parte más alta a la más baja; sin embargo, la proporción de rizomas vivos se incrementó (de 38,5 a 57,7%) y decreció la fitomasa de raíces vivas (de 17,8 a 5,6%). Estos autores también determinaron una correlación lineal negativa entre la fitomasa de las raíces y la humedad del suelo, en las capas superficiales.
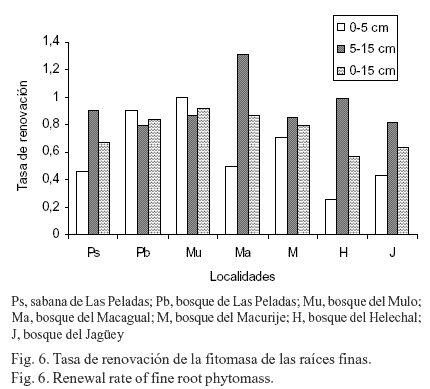
Tasa de renovación. La menor tasa de renovación (TR) se produjo en la localidad sabana de Las Peladas (fig. 6), en la capa de suelo de 0-5 cm, pero en la de 5-15 cm estuvo entre las más altas de las localidades estudiadas. Existieron diferencias notables en la TR entre las capas de suelo y entre las localidades, lo que pudiera estar dado por la humedad del suelo y las grandes diferencias que existen en las montañas en cuanto a la exposición, la pendiente, la altura de la vegetación y las especies predominantes, así como por las actividades antrópicas en cada área (Caldwell y Virginia, 1989). Excepto en dos de las áreas (el bosque de Las Peladas y del Mulo), la TR fue mayor en la capa de suelo 5-15 cm, debido posiblemente a la menor variabilidad de la humedad del suelo en esta capa. La TR promedio (± EE) de la fitomasa de las raíces para todas las localidades estudiadas fue de 0,60 (± 0,10) para la capa 0-5 cm y de 0,92 (± 0,07) en la capa de suelo de 5-15 cm (volumen dos veces mayor que la capa superficial); este resultado también podría ser consecuencia, entre otros factores, de una mayor estabilidad de la humedad del suelo en esta capa y la mayor proporción de raíces finas (Hernández, 1999).
En conclusión, el presente estudio evidenció que la humedad del suelo desempeña un papel importante en la producción de raíces finas en los sistemas boscosos, tal como ha sido informado para diferentes sistemas herbáceos (Hernández y Fiala, 1992; Hernández, 1999). Este estudio, igualmente, corroboró la importancia de la primera capa del suelo en la dinámica de la humedad del sustrato y de la fitomasa de las raíces finas, aspectos que se deben considerar para el funcionamiento adecuado de estas formaciones vegetales en la RBSR y otras localidades con características afines, y también para su manejo futuro.
AGRADECIMIENTOS
Dejamos constancia de nuestro agradecimiento al Dr. Héctor Sagué Díaz, quien iniciara los estudios hidrológicos y sus procesos en la RBSR; a los compañeros del antiguo Departamento de Ecología Forestal de la Academia de Ciencias de Cuba (Leudelio Lastres, Justo Arteaga y Rolando Francés); a la dirección del Plan Las Terrazas, por el financiamiento y la construcción de las instalaciones que sirvieron de base para las observaciones y los resultados de este trabajo. A los doctores Ricardo A. Herrera y Maritza García, y a los demás compañeros de la Estación Ecológica, quienes hicieron posible la toma de las muestras y el proceso para la determinación de la humedad del suelo. Esta investigación también fue parcialmente financiada por la Fundación Internacional para la Ciencia (IFS), mediante el donativo D/3536-2 a J. A. Sánchez. ]]>
REFERENCIAS BIBLIOGRÁFICAS
1. Bazzaz, F.A. Plants in changing environments: linking physiological, population, and community ecology. Cambridge University Press. Cambridge, UK. 320 p. 1996
2. Blanco, F.A. Métodos apropiados de análisis estadísticos subsiguientes al análisis de varianza (ANDEVA). Agronomía Costarricense. 25:53. 2001
3. Bucci, S.J. et al. Functional convergence in hydraulic architecture and water relations of tropical savanna trees: from leaf to whole plant. Physiology. 24:891. 2004
4. Caldwell, M.M. &. Virginia, R.A. Root systems. In: Plant physiological ecology. (Eds. R. Pearcy et al.). Chapman and Hall. New York. p. 367. 1989
5. FAO. State of the world´s forests. FAO. Rome, Italy. 164 p. 2011
6. Fiala, K. et al. Biomass partitioning in Paspalum notatum stands on the slope relief in the anthropic savanna of Cuba. Preslia. 63:269. 1991
7. Fornaris E.G. et al. Características fisonómicas y funcionales del bosque siempreverde mesófilo de las cuchillas del Toa, Guantánamo, Cuba. Biodiversidad Cuba Oriental. 4:36. 2006
8. Hernández, A. et al. Los suelos de la Reserva. En: Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Proyecto MAB No. 1, 1974-1987. (Eds. R.A. Herrera et al.) ROSTLAC, Montevideo. p. 88. 1988 ]]>
9. Hernández, L. Fitomasa subterránea en un pastizal de Paspalum notatum, en la Sierra del Rosario, Cuba. Tesis presentada en opción al título de M.Sc. en Ecología. Instituto de Ecología y Sistemática. Ciudad de La Habana, Cuba. 85 p. 1999
10. Hernández, L. & Fiala, K. Root biomass dynamics in the savanna communities of Paspaletum notati in Cuba. Ekológia. 1:153. 1992
11. Hernández, L. & Rodríguez, María. La fitomasa subterránea en bosques naturales y en áreas de rehabilitación de la región minera de Moa, Holguín, Cuba. Acta Bot. Cub. 211:36. 2011
12. Hernández, L. & Sánchez, J.A. 2009. Distribución del agua de lluvia y factores microclimáticos en ecosistemas de la Reserva de la Biosfera «Sierra del Rosario», Cuba. [cd rom] Memorias del V Congreso de Meteorología. Ciudad de La Habana, Cuba
13. Hernández, L. et al. Caracterización espacial de la biomasa subterránea en pastizales del Instituto de Ciencia Animal. Acta Bot. Cub. 116:1. 1998
14. Hernández, L. et al. Biomasa vegetal subterránea en áreas naturales, contaminadas y reforestadas de la región minera de Moa, Cuba. En: Memorias de la Cuarta Conferencia Internacional sobre Ecología de la Serpentina. http://www.sciencereviews2000.co.uk/. 2003
15. Hernández, L. et al. Fine root biomass of various Cuban forests indicating the resistance to disturbance. Memorias. Symposium, Solvakia. http://www.uke.sav.sk. p. 240. 2006
16. Herrera, P. Flora y vegetación. En: Biodiversidad de Cuba. (Ed. H. González). Ediciones Polymita. La Habana, Cuba. p. 142. 2007
17. Herrera, R.A. et al. Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Proyecto MAB No. 1, 1974-1987. ROSTLAC, Montevideo. 760 p. 1988a
18. Herrera, R.A. et al. Caracterización y dinámica de la fitomasa de raíces y micorrizas vesículo-arbusculares en la Sierra del Rosario. En: Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Proyecto MAB No. 1, 1974-1987. (Eds. R.A. Herrera et al.). ROSTLAC, Montevideo. p. 447. 1988b ]]>
19. Herrera, R.A. et al. Ecotechnologies for the sustainable management of tropical forest diversity. Nature and Resources. 33:2. 1997
20. Hertel, D. et al. Size and structure of fine root systems in old-growth and secondary tropical montane forest (Costa Rica). Biotropica. 35:143. 2003
21. Kaúrichev, I.S. Prácticas de edafología. Editorial MIR. Moscú. 287 p. 1980
22. Mancina, C.A. et al. Habitat use by phyllostomid bat assemblages in secondary forests of the «Sierra del Rosario» Biosphere Reserve, Cuba. Acta Chiropterologica. 9:203. 2007
23. Priess, J.A. et al. Litter and fine-root production in three types of tropical premontane rain forest in SE Venezuela. Plant Ecol. 143:171. 1999
24. Sagué, H. & Hernández, L. Estructura del sistema radical de dos comunidades vegetales de suelos montañosos. Inf. Cient. Téc. 41:9. 1978
25. Sagué, H. et al. Balance hídrico y erosión en Sierra del Rosario. Voluntad Hidraúlica. 49:50. 1979
26. Sánchez, J.A. et al. Ecological grouping of tropical trees in an evergreen forest of the Sierra del Rosario, Cuba. Acta Bot. Cub. 204:14. 2009
27. Sarmiento, G. & Vera, M. La marcha anual del agua en el suelo en sabanas y bosques tropicales en los llanos de Venezuela. Agronomía Tropical. 27 (6):629. 1976
28. Singh, K.P. & Singh, R.P. Seasonal variation biomass and energy of small roots in tropical dry deciduos forest, Varanasi, India. Oikos 37:88. 1981
Recibido el 10 de mayo del 2011
Aceptado el 19 de septiembre del 2011 ]]>
{kind=link}
{kind=link}
{kind=link}
{kind=link}