REPORT
Identification of a new trypsin from Spodoptera frugiperda involved in a defensive mechanism against the Bacillus thuringiensis Cry1Ca1 toxin
Identificación de una nueva tripsina de Spodoptera frugiperda que participa en un mecanismo de defensa contra la toxina CryCa1 de Bacillus thruringiensis
]]>
Lianet Rodríguez1, Orlando Borrás2, Pilar Téllez1, Ivis Morán1, Milagros Ponce1, Yanileysis Fernández1, Camilo Ayra1
1 Laboratorio de Biotecnología Ambiental.
2 Laboratorio de Genómica Funcional de Plantas. División de Plantas, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 10600, AP 6162, La Habana, Cuba.
ABSTRACT
Both, insecticidal formulations based on the entomopathogenic bacterium Bacillus thuringiensis (Bt) and transgenic plants expressing Bt Cry toxins, have been threatened by the potential appearance of insect resistance in major crop pests. With the aim of identifying genes and mechanisms triggered against a Bt Cry1-class toxin in the midgut of fall armyworm Spodoptera frugiperda, the major pest of maize in Cuba, subtractive cDNA libraries of the molecular interaction insect-Bt Cry1Ca1 toxin were constructed. Among those genes specifically regulated in response to the intoxication, one coding for a new trypsin-like serin proteinase (SfT6) was identified. Gene function analysis using RNA interference showed SfT6 plays a crucial role for the Cry1Ca1 toxicity against S. frugiperda; gene expression suppression caused a reduction of the proteolytic processing of Cry1Ca1 by the larval midgut juice and a reduced susceptibility of insects in bioassays. Our study represents the first report on S. frugiperda midgut genes differentially-expressed in response to Cry1Ca1 intoxication. Besides, we have identified and cloned the full-length cDNA sequence of a novel serine proteinase whose regulation is linked to the natural process of insect adaptation to Bt in order to survive the pathogenic process.
Keywords: Bacillus thuringiensis, Spodoptera frugiperda, Cry1Ca1, serine-proteinase, trypsin, RNA interference.
]]>
INTRODUCTION
For decades, commercial formulations of the sporogenic bacterium Bacillus thuringiensis (Bt) and its parasporal crystal inclusions (composed mainly by the insecticidal Cry proteins) have been sprayed on economically important crops, sometimes indiscriminately, for the control of major insect pests [1]. More recently, Bt cry genes have been introduced into cultivable plants, particularly cotton and maize, to produce insect-resistant Bt-crops that in 2009 were grown on 50.4 million ha worldwide [2]. The major concern of such widespread use of Bt products deals with the possibility of the that developed resistance by key pest species. To date, three lepidopteran pest species have evolved substantial resistance to Bt sprays under field conditions [3, 4] while the first cases of field-evolved resistance to Bt-crops have been recently documented [5].
The Bt-insect interaction is a complex multi-step process ending up in larval death when it’s effective. In Lepidoptera, ingested Bt spore and crystals reach the extremely alkaline and reducing environment of the larval midgut where spore germination is activated and crystal inclusions solubilize to inactive protoxins of 130-140 kDa in size. Then, midgut serine-proteinases transform protoxins into active toxins of 55-65 kDa after removing approximately 600 residues from the C-terminus and the first 28 N-terminal residues [6]. Activated monomers of the Cry toxins can cross the peritrophic matrix barrier to bind specific membrane receptor(s) on epithelial target cells, where further proteolysis may take place before pore-forming structures are formed by the plasma membrane inserted into to disturb the ionic and osmotic balances of the cell [7]. Finally, germinated spores swing into the insect hemolymph through the gaps in the epithelial cells barrier left by Cry-bursted cells to colonize the entire larval body causing septicemic death.
The molecular study of host – pathogen interaction can reveal important features of the pathogenic process as well as identify those components of defensive mechanisms triggered in the host. For insect-Bt interaction, such defensive responses could lead to insect adaptation to the entomopathogen after continuous exposures inducing a considerable resistance level in successive generations. One effective approach for identifying insect genes differentially expressed during exposure to Bt Cry toxins is suppression subtractive hybridization (SSH) [8]. SSH is based on an efficient subtraction step followed by a suppression step. PCR amplification allows the enrichment of the differentially expressed genes but suppression of those that are not differentially expressed. In addition, a normalization step is incorporated to equalize the abundance of individual mRNAs, in order to provide a better chance for detection of rare transcripts. Since SSH allows the isolation of differentially expressed cDNAs without a prior knowledge of their sequence, it is highly desirable for studying differential gene expression in systems where information on the genomic sequence is scarce (i.e., insects). Further, gene function studies can be accomplished in vivo by RNA interference where specific gene expression is suppressed or completely silenced after feeding larvae with a double-stranded RNA (dsRNA) synthesized from a large fragment of the correspondent gene [9].
In the present work, the molecular response elicited by Bt Cry1Ca poisoning in Spodoptera frugiperda (Lepidoptera: Noctuidae), the major pest of maizein Cuba, is investigated. Subtractive cDNA libraries enriched for S. frugiperda midgut genes differentially regulated during the exposure to a sublethal dose of the toxin were generated. Expression analysis of SSH clones showing a significant hit in the BLAST analysis and which reflected the distribution of molecular functions in the subtractive cDNA libraries allowed the identification of a new trypsin-like serin proteinase (SfT6) that was chosen for gene function analysis by RNAi. Gene expression suppression of SfT6 decreased the pathogenicity of Bt Cry1Ca1 in S. frugiperda larvae and the protoxin activation rate by larval gut juice corroborating its crucial role for the Bt Cry1Ca1 toxic pathway in this important Lepidoptera.
RESULTS AND DISCUSSION
The present study aimed at identifying genes and mechanisms of the lepidopteran larval gut involved in the specific response against Cry1-class toxins of B. thuringiensis. To achieve this goal, we performed the molecular analysis of S. frugiperda-Cry1Ca1 toxin interaction and investigated the role of a new trypsin-like serine-protease (SfT6) differentially expressed in the larval midgut during the process of Bt intoxication. To this end, the following strategies were used: i) construction and characterization of subtractive cDNA libraries from the larval midgut tissue during the interaction S. frugiperda-Cry1Ca1, ii) cloning of the cDNA full-length sequence of a novel S. frugiperda trypsin, named here as SfT6, and its sequence analysis, iii) RNAi-mediated gene silencing of SfT6 for function analysis through insect bioassays and in vitro proteolytic processing of the Cry1Ca1 protoxin.
Since a sublethal dose of 2 µg Cry1Ca1/cm2 caused a complete inhibition of the larval growth and less than 5% mortality in S. frugiperda third-instar larvae [10], this toxin concentration was used for studying the early transcriptional response of midgut cells from larvae fed with Cry1Ca1. The construction of subtractive cDNA libraries enriched in larval midgut genes differentially regulated during the intoxication process yielded fragments ranging in size from 250 to 1000 base pairs (bp). The quality of subtracted cDNA libraries was validated by RT-PCR amplification of an internal fragment of the constitutively expressed β-actin transcript, and by hybridization with radioactive 32P-labeled probes obtained from the unsubtracted cDNA pool [10]. In addition, the analysis of signal differentials between unsubtracted/subtracted cDNA hybridizations allowed the identification of 86 clones that were the most relevant according to their regulation. The DNA sequence of selected 86 clones was compared with GenBank/EMBL databases using the BLASTN/BLASTX algorithms. According to results, 23% of the cDNA fragments showed significant similarity to gene products involved in known biological processes; with disease/defense, metabolism (catalytic activities) and oxidative stress being the most frequently identified by using a Gene Ontology software.
]]> One of the most interesting clones of the libraries according to its expression regulation, with a significant hit in the BLAST analysis, was a serine-proteinase named as SfT6. The full-length cDNA sequence of SfT6 was PCR cloned by RACE. A fragment of 904 bp length was obtained that contained a 765 bp Open Reading Frame (ORF), spanning nucleotides 57 to 821. The ORF encoded a protein consisting of 254 amino acids with a calculated molecular mass for the mature protein of 25.1 kDa and isoelectric point of 8.6. Multiple alignment and phylogenetic relationship analysis of predicted amino acids for SfT6 mature polypeptide denoted that the cloned fragment belongs to a trypsin-like serine-proteinase gene. The phylogenetic tree showed that the S. frugiperda SfT6-predicted polypeptide is in the same branch as trypsin-like deduced proteins from most lepidopteran annotated sequences (Figure 1A). Specific alignment of SfT6 with partial sequences of S. frugiperda trypsin-like serine proteinases found in the integrative database SPODOBASE [11] yielded only a marginal identity of 20% among them. RT-PCR reactions conducted to investigate the developmental expression pattern of SfT6 showed that this gene is transcribed during the actively feeding stages of the insect life (larvae and adult) while it is totally absent in embryonic and pupal stages (Figure 1B). The accumulation rate of SfT6 dramatically increased during the progression from neonates to sixth larval instar suggesting an important role of this protease for food digestion during larval growth and development. However, the expression of SfT6 was visibly affected when larvae were exposed to Cry1Ca1 toxin (Figure 1C). SfT6 mRNA levels were shown to rapidly decline by 15 min of intoxication while the expression of other two larval trypsins (Sf1f02950-3-1 and Sf1f04740-5-1) remained invariable. With this result, we show for the first time, S. frugiperda can mount a defensive response to a Bt Cry1-class toxin based on, among others, a decreased expression in one particular serine proteinase from the midgut [12].Considering the crucial role of midgut serine proteinases for the proteolytic activation of Cry1-class proteins during the Bt toxic pathway in Lepidoptera [6, 13], the specific downregulation of SfT6 expression in S. frugiperda larval midgut during the early stages of Cry1Ca1 intoxication suggests a defensive mechanism adopted by the insect to survive the pathogenic process. To prove this hypothesis, we performed RNAi-mediated gene silencing in S. frugiperda by feeding newly molted fourth-instar larvae with double-stranded RNA (dsRNA) produced from a fragment of SfT6 gene. As controls, two groups of insects fed with either buffer alone (10 mM Tris-Cl, pH 7.5; 10 mM EDTA) or a nonspecific dsRNA obtained from the β-glucuronidase (gus) gene from Escherichia coli were used. Both, insects’ susceptibility to Cry1Ca1 and in vitro activation of Cry1Ca1 protoxins by larval midgut juice proteases, were evaluated in SfT6-knockdown insects and compared to insects from control groups. The efficacy of SfT6-knockdown was investigated 48 h after the corresponding RNAi treatment by qRT-PCR analysis on larval midgut total RNA. The results revealed that the oral administration of dsRNA-SfT6 caused a significant reduction of SfT6 gene expression in the midgut (approx. 90% of control), while feeding with dsRNA-Gus showed no effect on the SfT6 levels, confirming the specificity of the process for the target gene [12].
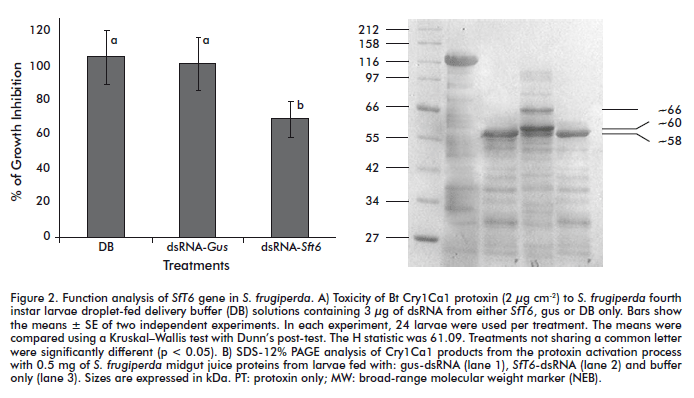
The results of Cry1Ca1 toxicity are shown in Figure 2A, expressed as percentage of growth inhibition(%GI) and determined in dsRNA-treated and untreated larvae 48 h after the dsRNA feeding assay. No differences in %GI values were found between larvae that were only fed with buffer (mocked) or nonspecific dsRNA-Gus. In contrast, larvae from the group fed with dsRNA-SfT6 were ~30% less affected by the Cry1Ca1 protoxin in terms of %GI at the same tested concentration. On the other hand, in vitro protoxin processing experiments showed that after 1 h of Cry1Ca1 incubation with gut juice from larvae fed with nonspecific dsRNA-Gus, a single band of ~58 kDa was generated that did not differ from that obtained with gut juice from mocked (buffer-fed) larvae (Figure 2B, lanes 1 and 3). On the contrary, a moderate but clear decrease in protoxin activation rate was found when midgut juice from dsRNA-SfT6-fed larvae was used in the reaction (Figure 2B, lane 2). In this case, the midgut juice failed to completely process the Cry1Ca1 protoxin to the ~58 kDa product (the apparent activated form of Cry1Ca1 toxin in S. frugiperda) but produced in addition two major partially processed products of ~60 and ~66 kDa (Figure 2B, lane 2).
In summary, our results represent the first study on molecular defensive mechanisms triggered in the midgut cells of S. frugiperda larvae after intoxication with a B. thuringiensis Cry1-class toxin [10, 12]. Also, we have identified, cloned and characterized a novel trypsin gene from the S. frugiperda larval midgut that participates in a defensive mechanism elicited in response to the Bt Cry1Ca1 pathogenic process [12]. We have shown that the specific down-regulation of SfT6 serin-proteinase increases larval survival after exposure to Cry1Ca1 in bioassays and affects in vitro proteolytic processing of the protoxin in the presence of larval gut juice. Down-regulation of SfT6 could represent a putative defensive mechanism to reduce the amounts of activated toxin monomers that can reach target gut cells, providing with a survival advantage for insects feeding on Cry1Ca1-contaminated diets. In this way, unprocessed protoxins cannot pass the peritrophic matrix and can be rapidly eliminated during the gut clearance process. Regulation of SfT6 expression could become a target in a proteinase-mediated Bt-resistance mechanism. Indeed, significantly reduced mRNA levels of the SfT6 ortholog in Plodia interpunctella were found in a resistant population of this insect [14].
ACKNOWLEDGMENTS
The authors appreciate Mr. Rafael Maura assistance on insect manipulation and dissection during RNAi experiments.
REFERENCES
1. Federici BA. Insecticidal bacteria: an overwhelming success for invertebrate pathology. J Invertebr Pathol. 2005;89: 30-8.
]]>2. Clives J. Brief 41: Global Status of Commercialized Biotech/GM Crops: 2009. Ithaca (NY): ISAAA; 2009 [cited 2011 Jan 20]. ISAAA Brief No. 41. Available from: http://www.isaaa.org/resources/publications/briefs/41/default.asp.
3. Tabashnik BE, Cushing NL, Finson N, Johnson MW. Field development of resistance to Bacillus thuringiensis in diamondback moth (Lepidoptera: Plutellidae). J Econ Entomol. 1990;83:1671-6.
4. Janmaat AF, Myers J. Rapid evolution and the cost of resistance to Bacillus thuringiensis in greenhouse populations of cabbage loopers, Trichoplusia ni. Proc Biol Sci. 2003;270(1530):2263-70.
5. Tabashnik BE, Van Rensburg JB, Carrière Y. Field-evolved insect resistance to Bt crops: definition, theory, and data. J Econ Entomol. 2009;102(6):2011-25.
6. Lightwood DJ, Ellar DJ, Jarrett P. Role of proteolysis in determining potency of Bacillus thuringiensis Cry1Ac delta-endotoxin. Appl Environ Microbiol. 2000; 66(12):5174-81.
]]>7. Bravo A, Gill SS, Soberón M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon. 2007;49(4):423-35.
8. Diatchenko L, Lau YF, Campbell AP, Chenchik A, Moqadam F, Huang B, et al. Suppression subtractive hybridization: a method for generating differentially regulated or tissue-specific cDNA probes and libraries. Proc Natl Acad Sci USA. 1996; 93(12):6025-30.
9. Turner CT, Davy MW, MacDiarmid RM, Plummer KM, Birch NP, Newcomb RD. RNA interference in the light brown apple moth, Epiphyas postvittana (Walker) induced by double-stranded RNA feeding. Insect Mol Biol. 2006;15(3):383-91.
10. Rodríguez-Cabrera L, Trujillo-Bacallao D, Borrás-Hidalgo O, Wright DJ, Ayra-Pardo C. Molecular characterization of Spodoptera frugiperda-Bacillus thuringiensis Cry1Ca toxin interaction. Toxicon. 2008; 51(4):681-92.
11. Fournier P. SPODOBASE [Internet]. Paris: INRA. c2003 - [cited 2010 Mar 17]. Available from: http://bioweb.ensam.inra.fr/spodobase/
12. Rodríguez-Cabrera L, Trujillo-Bacallao D, Borrás-Hidalgo O, Wright DJ, Ayra-Pardo C. RNAi-mediated knockdown of a Spodoptera frugiperda trypsin-like serine-protease gene reduces susceptibility to a Bacillus thuringiensis Cry1Ca1 protoxin Environ Microbiol. 2010;12(11):2894-903.
13. Oppert B. Protease interactions with bacillus thuringiensis insecticidal toxins. Arch Insect Biochem Physiol. 1999;42(1):1-12.
14. Zhu YC, Oppert B, Kramer KJ, McGaughey WH, Dowdy AK. cDNA sequence, mRNA expression and genomic DNA of trypsinogen from the indianmeal moth, Plodia interpunctella. Insect Mol Biol. 2000; 9(1):19-26.
Camilo Ayra. Laboratorio de Biotecnología Ambiental. División de Plantas, Centro de Ingeniería Genética y Biotecnología, CIGB. Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 10600, AP 6162, La Habana, Cuba. E-mail: camilo.ayra@cigb.edu.cu .
]]>{kind=link}