RESEARCH
Recombinant hybrid proteins from pertactin type 1 and 2 of Bordetella pertussis are more immunogenic in mice than the original molecules
Proteínas híbridas recombinantes de pertactinas tipo 1 y 2 de Bordetella pertussis son más inmunogénicas en ratones que las moléculas originales
]]>
Diógenes Quintana-Vázquez1, Edelgis Coizeau1, Anabel Alvarez1, Maité Delgado1, Tania Cárdenas1, Yassel Ramos1, Glay Chinea1, Guy A M Berbers2, Gerardo E Guillén1
1 Centro de Ingeniería Genética y Biotecnología, CIGB Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba.
2 Laboratory for Infectious Diseases and Screening, National Institute for Public Health and the Environment. Bilthoven, The Netherlands.
ABSTRACT
The present study explores the concept of hybrid pertactin (PRN) molecules for immunizing against Bordetella pertussis. New molecules were designed using an additive/inclusive approach that comprehends the complete sequences/ regions of two different types of pertactin (Prn). PRN molecules bear the two variable R1 regions from Prn1 and Prn2. The genes of Prn1, Prn2 and six variants of PRN were cloned in Escherichia coli, and PRN proteins were over-expressed at 25-30 % of total protein concentrations using the pET28a/BL21 Codonplus RP expression system. The proteins were purified (> 90 % purity) using the His-tag /Ni-NTA affinity method with amounts of 8-10 mg/g of wet biomass. After refolding, the PRNs were recognized by anti-Prn monoclonal antibodies that bind protective conformational and linear epitopes/regions. Moreover, a panel of ten sera from individuals boosted with a commercial vaccine reacted with the PRN molecules without differences from the P.69 protein. The PRN proteins were highly immunogenic in Balb/c mice, with the induction of the IgG2a and IgG2b subtypes. Particularly, two PRNs (PRN2-lc-1 > PRN2-1) induced highly significant anti-Prn1 antibody levels (p < 0.001). Moreover, the PRN2-lc-1 induced higher levels of antibodies (p < 0.05) against epitopes located at the immunodominant N-terminus region and the variable region R1. The PRN2-lc-1 and PRN2-1 molecules exhibited an enhanced immunological profile in Balb/c mice in terms of level of whole anti-Prn IgG antibodies in respect to natural Prn controls. These two molecules constitute valuable candidates for further evaluation in vivo in acellular vaccine formulations.
]]> Keywords: hybrid pertactin, Bordetella pertussis, acellular vaccine, immune response, antibodies, variable regions.RESUMEN
En este estudio se exploró el concepto de moléculas híbridas de pertactina (PRN) para inmunizar contra Bordetella pertussis. Se diseñaron nuevas moléculas mediante un método aditivo/inclusivo que comprendió las secuencias completas/regiones de dos pertactinas (Prn). Las moléculas PRN portan las dos regiones variables R1 de Prn1 y Prn2. Se clonaron los genes de Prn1, Prn2 y seis variantes de PRN en Escherichia coli. Las PRN se sobre-expresaron a 25-35 % de la concentración de proteínas totales, con el uso del sistema de expresión pET28a/BL21 Codonplus RP. Se purificaron por el método de afinidad His-tag /Ni-NTA proteínas con más del 90 % de pureza, con rendimientos de 8-10 mg/g de biomasa. Tras la reconstitución, las PRN fueron reconocidas por anticuerpos monoclonales anti-Prn contra epitopos y regiones conformacionales y lineales, involucrados en la respuesta inmune protectora. Un panel de 10 sueros de individuos reactivados con una vacuna comercial reaccionaron con las moléculas PRN a niveles similares a los obtenidos con la proteína P.69. Las proteínas PRN fueron altamente inmunogénicas en ratones Balb/c, e indujeron respuesta de anticuerpos IgG2a e IgG2b. Dos proteínas PRN (PRN2-lc-1 > PRN2-1) indujeron niveles significativos de anticuerpos (p < 0.05) contra epitopos localizados en el extremo N-terminal inmunodominante y en la región variable R1. Estas dos proteínas mostraron un perfil de respuesta potenciada de anticuerpos (anticuerpos IgG totales antiPrn) en ratones Balb/c comparadas con los controles de Prn. PRN2-lc-1 y PRN2-1 son valiosos candidatos para evaluaciones posteriores in vivo, como parte de formulaciones de vacunas acelulares.
Palabras clave: pertactina híbrida, Bordetella pertussis, vacuna acelular, respuesta inmune, anticuerpos, regiones variables.
INTRODUCTION
The gram negative bacterium Bordetella pertussis causes a highly contagious disease known as pertussis [1]. The disease was a major cause of infant death before pertussis vaccines were available [2]. The development of vaccines and its early implementation contributed to an overt reduction of pertussis cases worldwide [3]. However, after decades of vaccination the disease reemerged in highly vaccinated populations [4] to become nowadays one of the most prevalent vaccine-preventable disease of developed countries [5].
]]>The number of circulating strains expressing the pertactin variant Prn2 has had a gradual increase in the last decades. However, the strains used in vaccine manufacture bear pertactin types different to Prn2 such as Prn1 [5]. Notably, the differences between Prn1 and Prn2 are only limited to the variable region 1 (R1), which consist of GGXXP repeats. It has been reported that variation in region 1 affects antibody binding and also the efficacy of a commercial whole-cell vaccine. In this study, passive and active immunization indicates that region 1 confers protective immunity and also contains a B-cell epitope recognized both in humans and mice [6]. Functional studies in mice suggest that the incorporation of Prn2 to acellular vaccine formulations could contribute to the efficacy against pertussis [7].
Since current vaccines contain Prn1 and circulating strains, depending on the region or epidemiological scenario, can express Prn1 or Prn2, a technological improvement could be the use of a hybrid pertactin (PRN) containing the relevant epitopes of Prn1 and Prn2. Theoretically, one hybrid molecule could contain two different types of pertactin as a consequence of including the variable R1 region from each type. In addition to expanding the vaccine spectrum, the new molecule may be more immunogenic due to it contains a greater number of functional epitopes, as a consequence to a duplication of a conserved region with immunological relevance. The PRN concept can facilitate the production process and the widespread use of a future vaccine, since a single downstream processing can result in a molecule representing the pertactin types predominant in circulating strains among populations of different epidemiological settings.
The Escherichia coli host has been successfully used to obtain novel molecules in an attempt to simplify pertussis vaccines production [8, 9]. Particularly, the Prn protein has been efficiently expressed in E. coli with the preservation of relevant epitopes [10].
Based on these precedents, the concept of PRN based on Prn1 and Prn2 was experimentally addressed to study the expression levels, folding, purification and the assessment of immunogenicity in Balb/c mice.
MATERIALS AND METHODS
Prn sequences
The extracellular domain sequences of Prn1 (D1-G677, Uniprot Accession Number: Q546U4) and Prn2 (D1-G682, Uniprot Accession Number: O88143) were taken as reference for amino acid positions and are abbreviated as Ref. Prn1 or Ref. Prn2 respectively. Henceforward, amino acid positions are referred to Ref. Prn1 unless otherwise stated.
]]> The protein sequences encoding the extracellular domain of Prn were obtained from Uniprot and Genbank databases: B. parapertussis gen BPP 1150 (Uniprot Accession Number: P24328) and the 13 Prn types described for B. pertussis : Prn1(Uniprot: Q546U4); Prn2 (Uniprot: O88143); Prn3 (Uniprot: O69259); Prn4 (Uniprot: Q9S6M9); Prn5 (Uniprot: Q9S3M8); Prn6 (Uniprot: O69257); Prn7 (Uniprot: Q9S6N1); Prn8 (Uniprot: Q9S6N0); Prn9 (Uniprot: Q93L98); Prn10 (Uniprot: Q8RSU0); Prn11 (Uniprot: Q8GJ68); Prn12 (GenBank: BAF35031.1), which was translated to amino acid sequence and Prn13 (Uniprot: A4UTL5).Bioinformatics tools
Protein sequences were aligned using ClustalW2 [11] available at http://services.uniprot.org. Positions of conserved sequences identified were visualized in CHIMERA visualization system [12].
The structural predictions of mature Prn1 and Prn2 of B. pertussis and Prn of B. parapertussis (henceforward as Prn B. pp) were performed using the I-TASSER algorithm server (http://zhang.bioinformatics.ku.edu/I-TASSER/) [13]. The coordinates of Prn under code 1DAB.pdb from the Protein Data Bank [14] were provided to the server as template.
The sequences encoding the fragments A and B of the hybrid pertactin molecules were selected using VMD 1.8.7beta5.
Bacterial strains, reagents and growth conditions
The E. coli strain Top 10 F’ (Invitrogen) was used for cloning procedures following standard methods [15], and the E. coli strain BL21 (DE3)-codonplus RP (Novagen) for protein expression. Luria-Bertani broth (LB) was used as standard growth medium for both plasmid manipulation and protein expression. When necessary, culture media were supplemented with the appropriate antibiotics: ampicillin (100 μg/ mL), kanamycin (50 μg/mL) and chloramphenicol (34 μg/mL).
Construction of pET28a-prn1 and pET28a-prn2
The DNA fragments encoding the extracellular domain of Prn were amplified by polymerase chain reaction (PCR) from genomic DNA isolated from B. pertussis strain Tohama I (prn1) [16] and a clinical isolate (prn2). The PCR was performed with the primers pET-A0-new-L and pET-D-new as previously reported [10]. The PCR fragments were then digested with endonucleases Nde I and BamH I and inserted into pET28a plasmid (Novagen) to give rise to pET28a-prn1 and pET28a-prn2 expression vectors. The final constructions were selected after satisfactory restriction enzyme digestion and DNA sequence analyses.
Construction of pET28a expression vectors of hybrid pertactin of Prn1 and Prn2
]]> The hybrid pertactin molecules were generated using inverse PCR [17]. In all cases the expression vectors pET28a-prn1 and pET28a-prn2 were used as templates. Primers are shown in the table.The DNA fragments were amplified using 0.1 μg of the appropriate template vector in a 50 μL PCR containing 2.5 units of Platinum Pfx DNA polymerase HF (Invitrogen), 0.3 μM of each primer, 0.3 μM of each dNTP, 1mM MgSO4 and 2× PCRx Enhancer solution with the following parameters: 5 min. initial denaturation at 94 °C; 15 s denaturation at 94 °C, 5:30 min annealing-extension at 68 °C, (10 cycles) and 10 min final extension at 68 °C. The PCR products were gel-purified using Wizard SV Gel and PCR Clean-Up System (Promega). The purified blunt end products were ligated and the plasmids were subsequently introduced into TOP10 E. coli by transformation. Positive clones of the different pET28a hybrid pertactin vectors were subjected to sequence analysis to confirm the absence of point mutations.
The six PRN molecules’ constructs were obtained as described in the following:
Protein expression
The expression vectors for Prn1, Prn2 and hybrid pertactin proteins (PRN1-2, PRN1-sc-2, PRN1-lc-2, PRN2-1, PRN2-sc-1, PRN2-lc-1) were introduced into E. coli BL21-Codonplus (DE3)-RP (Novagen) as previously described [10]. Transformed cells from a single colony were grown at 37 °C under 100 rpm constant agitation in 50 mL LB media containing 50 μg/mL kanamycin and 50 μg/mL chloramphenicol. Cells from the overnight cultures were used to inoculate 300 mL of fresh medium (as described above) at an optical density at 600 nm (OD600) of 0.2 and further grown in one liter flasks at 37 °C and agitation until 0.5 OD600. Protein expression was induced by adding 1 mM IPTG. The cell cultures were further incubated for 4 h and cells were harvested by centrifugation (5000 g for 10 min) and stored at -20 °C until use. Protein expression was estimated by bacterial cells resuspension and rupture, followed by 10 % sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE), Coomassie brilliant blue staining and densitometry analysis. The relative percent of protein expression was calculated using the ImageJ software (National Institutes of Health).
SDS-PAGE
SDS-PAGE was performed to separate proteins as described [18]. Protein molecular size standard RPN5800 (GE Healthcare) were used as reference. The samples at appropriate concentrations were mixed with Tris-glycine SDS sample buffer (2×) and boiled for 3 min before loading onto 10 % Tris-glycine polyacrylamide gels. Bands were visualized by Coomassie brilliant blue staining.
His-tagged affinity chromatography of protein
Frozen bacterial pellets (1 g of wet weight) were suspended in 10 mL of lysis buffer (50 mM Tris HCl pH 8.0, 100 mM NaCl, 5 mM EDTA and 1 % Triton-X-100). The cell suspension was incubated in ice and sonicated for 1 min at a 70 % of amplitude setting, followed by a pause of 5 min. Sonication and pause steps were repeated four times. The insoluble material was collected by centrifugation and subsequently washed three times with a buffer containing 50 mM Tris HCl, 100 mM NaCl, pH 8.0. The pellet was then solubilized in 10 mL of solubilization buffer (4 M Urea, 10 mM benzamidine, 500 mM NaCl, and 50 mM Tris HCl, pH 8.0). After centrifugation, the supernatant was diluted to a final protein concentration of 3 mg/mL in equilibrium buffer (4 M Urea, 500 mM NaCl and 50 mM Tris HCl, pH 8). Then, 10 mL of the diluted sample was loaded onto a Ni-NTA affinity column (2 mL column volume; GE Healthcare, USA) using a peristaltic pump at a flow rate of 1 mL/min. The column was washed with 10 column volumes of equilibration buffer and subsequently with a similar volume of 10 mM imidazole in equilibration buffer. The pertactin proteins were eluted from the column in 10 mL of elution buffer (4 M Urea, 500 mM NaCl, 50 mM Tris HCl, pH 8.0 and 50 mM imidazole).
]]> Monoclonal antibodies and human seraMurine monoclonal antibodies (mAbs) PeM1, PeM4, PeM19 and PeM29 were previously described [19]. A stored panel of 10 sera from healthy individuals (adults) was used. The sera were previously obtained after one month of a booster vaccination with Adacel acellular vaccine (Sanofi Pasteur).
Refolding of proteins
The solution containing the purified unfolded proteins was diluted 10-fold in refolding buffer (50 mM Tris HCl pH 8.8, 100 mM NaCl, 1mM EDTA and 200 mM L-arginine) by slowly adding single drops into the stirred refolding buffer (at a rate of 1 mL/min.). Proteins were allowed to fully refold during 24 h dialysis at 4 °C against 2 L of refolding buffer. The refolded proteins were dialyzed against 50 mM Tris HCl, pH 8.8, using Spectra/Por 7 dialysis membranes with a molecular weight cut-off of 10 kDa (Spectrum Laboratories, CA). The proteins were concentrated on an Amicon concentrator with a 30 kDa cut-off (Millipore) to a final concentration of 0.4 mg/mL. The proteins were stored at -20 °C in the presence of 10 mM of benzamidine (Applichem, Germany).
The refolding procedure was evaluated by ELISA using the conformational mAbs PeM1 and PeM29. The mAbs were normalized to 0.5 mg/mL and assayed against P.69 Prn1 purified from B. pertussis (kindly provided by Chiron-Biocine, Siena, Italy) and the refolded Prn1 (recombinant) protein.
ELISA for antigenicity
Polystyrene high binding 96-well ELISA plates (Costar® Corning Incorporated, USA) were coated 3 h at 37 °C by adding 100 μL of 0.05 M carbonate buffer, pH 9.6 containing 2 μg protein/mL to each well. The wells were washed four times with PBS supplemented with 0.05 % Tween 20 (PBST) and then blocked with 4 % skim milk (Oxoid) in PBST for 1 h at 37 °C. In a previous experiment the mAbs were normalized to 0.5 mg/mL and titrated against P.69 (Prn1) using two fold serial dilutions in blocking solution and starting at 1:2000. As result the following dilution were used for the mAbs: PeM1 in 1:12 000; PeM4 in 1:4000; PeM19 in 1:6000 and PeM29 in 1:28 000, which give optical densities values of 0.7 to 1.0 in a linear interval (r2 = 0.997). Then, the plates were incubated for 2 h at 37 °C followed by four washings as described above. Bound mAbs were detected by using horseradish peroxidase-conjugated goat anti-mouse total IgG (Sigma, St. Louis, MI). The human sera were assayed at 1:100 dilution and a horseradish peroxidase-conjugated anti-human total IgG was used (Sigma). The optical density at 492 nm was measured with a multiscan plate reader (Merk). The mAb PeM19 was used to verify that equal amounts of protein were coated in all of the ELISA experiments (CV < 5 %).
The mAb PeM29 and the specific sera for P.69, Prn2, PRN1-lc-2 and PRN2-1 were evaluated against these same four proteins, both folded and unfold-ed. The proteins were denatured by heating 5 min at 95 °C.
The different binding of each mAb to hybrid molecules were established using a cut-off value established as follows: the optical densities from the hybrid pertactin proteins with coefficient of variation (CV) greater than 5.0 % in respect to P.69, Prn1 or Prn2 were considered with different (positive) binding activity. For human sera, the D´Agostino-Pearson normality test, Bartlett’s test for equal variances and ANOVA with Newman-Keuls multiple comparison test were performed. The statistic analyses were performed with Graphpad Prism 5.0.
Mass spectrometric analysis
]]> The identity of the different proteins was confirmed by mass spectrometric analysis using a hybrid quadrupole-orthogonal time-of-flight mass spectrometer with a nanospray ion source (QTOF-2TM, Micromass, UK).The SDS-PAGE protein bands (stained) were faded and in-gel digested with trypsin as previously described [20]. The mixture of tryptic peptides was loaded into the borosilicate nanoflow tips and submitted to 900 and 35 V of capillary and cone voltage, respectively. To acquire the ESI-MS/MS spectra, the first quadrupole was used to select the precursor ion within a window of 3 Th approximately. Argon gas was used in the collision chamber at ~3 10-2 Pa pressure and collision energies between 20 and 30 eV were set to fragment precursor ions. Data acquisition and processing were performed using MassLynx v3.5 (Micromass, UK).
Protein identification and concentration
Monoisotopic masses of tryptic peptides were used to identify the protein in a non-redundant database by peptide mass fingerprinting (PMF) with MASCOT (http://www.matrixscience.com). Additionally, the most intense signals of every spectrum were sequenced by collision induced dissociation. The MS/MS ion search option of the MASCOT software was used to confirm the protein identity. Parameter settings considered were the enzyme treatment, up to one missed cleavage, the molecular mass error (0.1 Da) and the modification on cysteine (propionamide cysteine). Other (variable) modifications such as methionine sulfoxide and the deamidation of glutamine and asparagine were also taken into account. The molecular weight of the protein and taxonomy were not restricted.
Protein concentration was estimated using BCA™ Protein Assay (Thermo Scientific).
Immunogenicity assay in Balb/c mice
Groups of female mice (n = 9) were immunized with the following proteins P.69, Prn1, Prn2, PRN1-2, PRN1-sc-2, PRN1-lc-2, PRN2-1, PRN2-sc-1, PRN2- lc-1 and a placebo (buffer phosphate instead of protein). The inoculations were given subcutaneously with 100 μL of a formulation containing 0.2 μg of protein adsorbed to 80 μg of aluminum hydroxide gel adjuvant (HCL Biosector). Mice received three doses at days 0, 14 and 28 and were bled at days -7, 14, 28 and 40. The anti-Prn1 IgG titles in sera were estimated by the ELISA described below.
Antibody response determination
The antibody response was determined in Polystyrene high binding 96-well ELISA plates (Costar® Corning Incorporated, USA). Briefly, plates were coated overnight at 4 °C by adding 100 μL of 0.05 M carbonate buffer, pH 9.6 containing 2 μg of Prn1 protein/mL to each well. The wells were washed four times with PBS supplemented with 0.05 % Tween 20 (PBST) and then blocked with 4 % skim milk (Oxoid) in PBST for 1 h at 37 °C. Then, the plates were incubated for 2 h at 37 °C with an appropriate dilution of serum. An anti-Prn1 serum was included as reference curve with 100 arbitrary title units. After four washings, bound IgG were detected by using horseradish peroxidase-conjugated goat anti-mouse total IgG (Sigma, St. Louis, MI). The optical density at 492 nm was measured with a multiscan plate reader (Merck). The statistic analyses were performed with Graphpad Prism 5.0. The D´Agostino-Pearson normality test, Bartlett’s test for equal variances and ANOVA with Newman-Keuls multiple comparison test were performed.
Immunoglobulin classes and subclasses were determined using a mouse isotyping kit (Sigma).
]]> The sera from groups immunized with P.69, Prn1, Prn2, PRN1-lc-2 and PRN2-1 were also tested against the following linear peptides (coating, 5 μg of peptide/mL): P1, RGDAPAGGAVPGGAVPGGAVPGGFGPGGFG PVLDGW, located at variable region R1 of Prn1; P2, RGDAPAGGAVPGGAVPGGFGPGGFGPGGFGPGGFGPVLDGW, located at variable region R1 of Prn2; P3, PQPGPQPPQPPQPQPEAPAPQPPAG, located at region R2 and P4, IVKTGERQHGIHIQGSDP, located at the N-terminus immunodominant domain. The differences observed were analyzed using Kruskal-Wallis statistic and Dunn’s multiple comparison tests.
RESULTS
Structural predictions
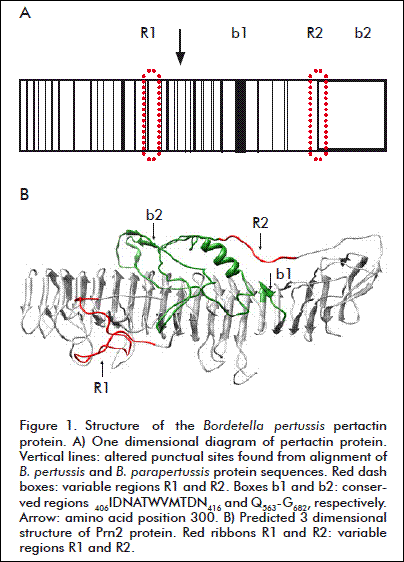
The structure of the Prn protein has been previously determined for the first 539 amino acids [14]. For this reason the conserved region Q558-G677 was not characterized by X-ray crystallography. In this work the structure of the whole extracellular domains of Prn1, Prn2 and Prn B. pp were predicted using I-TASSER [13]. The obtained models were structurally similar for the three proteins although the program estimated the Prn2 model as the more significant with a quality score of -1.21. As it is shown in figure 1, the conserved carboxyl region Q563-G682 (Ref. Prn2) was predicted to fold as a domain towards the amino end. In this region the predominant conformation was coil (90 %). The domain appeared connected to the helical stem by the variable region 2 (Figure 1B, R2).
Molecular design
The design of a PRN of Prn1 and Prn2 considered the following aspects. 1) The frequency of point amino acid mutations at the amino terminus. In the first 300 amino acids (D1-G300) appeared 66 % (35/53) of the altered sites (Figure 1A). 2) The conserved regions G377-L439 and Q558-G677. The region G377-L439 includes the sequence 401IDNATWVMTDN411, which locates near a domain involved in folding functions [21] while Q558-G677 sequence comprises the carboxyl end. 3) The structural proximity of the N-terminus and conserved C-terminus. This is in agreement with a previous report [19]. 4) The internal conserved domain involved in the folding function of Prn [21, 22]. Altogether, the regions mentioned comprise almost the entire Prn molecule. Hence, a reliable approach for designing a PRN should satisfy that the complete sequences/regions of the natural Prn protein are represented. This could be achieved with an additive/inclusive design, which means to add another/different variable R1 region to a Prn variant (Prn1 or Prn2).
From the structure analysis it was predicted that region V81-S269 (Ref Prn1) or V81-S274 (Ref Prn2) contains the smaller number of amino acids needed to preserve the variable loop (R1) natively structured. Therefore, if Prn2 or Prn1 protein frames are open at T80 and a V81-S269 (Ref Prn1) or V81-S274 (Ref Prn2) fragment is respectively inserted in the same orientation of the helical stem, PRN1-2 or PRN2-1 are obtained. For a practical purpose, as illustrated in figure 2, the molecules can also be obtained by the fusion of two fragments. For example, PRN1-2 can be formed by fusion of the fragments A = D1-S269 (Ref. Prn1) and B = V81-G682 (Ref. Prn2) to result in A-B (PRN1-2). Similarly, the PRN2-1 protein can be obtained by fusion of the fragments A = D1-S274 (Ref. Prn2) and B = V81-G677 (Ref. Prn1) to generate A-B (PRN2-1). Another feature considered in the design was the similarity of the helical cross section between the ends of the fragment to be linked. In proteins PRN1-2 or PRN2-1 there is a difference of 5 Å between the N-terminus of fragment B and the C-terminus of fragment A. To shorten this difference a chimeric rung IDNATWVMTDNIDNATWVMTDN was inserted as a long linker between fragments A and B which originated the PRN1-lc-2 and PRN2-lc-1 proteins. The chimeric rung is a tandem duplication of the conserved sequence IDNATWVMTDN. This sequence was also inserted as a short linker between fragment A and B to obtain PRN1-sc-2 and PRN2-sc-1.
Cloning, expression and mass spectrometric characterization of hybrid pertactin molecules
]]> The expression vectors for the expression of His-tag PRN proteins were constructed using inverse PCR [17]. Figure 3 exemplifies the procedure followed to obtain pET28a-prn1-2 vector. The other five vectors resulted from the same approach but combining different primer couples and vector templates (see Materials and methods for details). The constructs fulfilled restriction analysis with Nde I and BamH I endonucleases as well as DNA sequencing (data not shown). The expression vectors were used to transform the strain BL21(DE3) Codonplus RP. The overexpressed bands represented 25-30 % of the whole cell proteins (Figure 2C) and were analyzed by mass spectrometry. The signals of highest intensities were selected for MS/MS analysis and submitted for automatic search with the MASCOT software. This program indentified all the proteins with a high level of confidence (score: 900-1300) with peptide sequences corresponding to Prn of B. pertussis. The amino (GSHMDWNNQSIVK, m = 1514.71) and carboxyl ends (GDRGFTGDGGG, m = 994.46) of Prn were confirmed by manual inspection of the MS/MS spectra. Sequence coverage greater than 80 % was achieved for all PRN proteins.Purification and refolding of recombinant Prn1, Prn2 and the six hybrid pertactin molecules
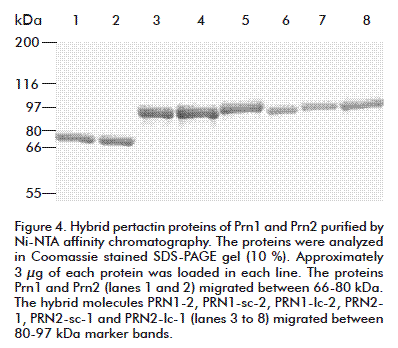
The His-tag recombinant proteins Prn1, Prn2 and the six PRNs were associated to the pellet of rupture and represented approximately the 45-49 % of the precipitated proteins. The insoluble aggregates were efficiently solubilized in the extraction buffer containing 4 M urea. After Ni-NTA affinity chromatography the proteins were obtained at levels of purity higher than 90 % (Figure 4).
The refolding procedure was verified comparing the reactivity of purified Prn1 (from E. coli) and natural pertactin P.69 Prn1 (from B. pertussis) with the conformational mAbs PeM1 and PeM29 as previously reported [10]. As depicted in figure 5A, both mAbs showed similar Abs values against the evaluated proteins (CV < 5 %).
The recovery of the purification procedure was approximately 60 %. The protein loss associated to column pass through, column washes, refolding (dialysis) and concentration were less than 1 %. The described method allowed the rapid purification of approximately 8-10 mg of refolded proteins per gram of wet biomass.
Antigenicity evaluation of the refolded hybrid Prn molecules
Figure 5 shows the positive recognition of PRN variants by four anti-Prn1 mAbs. As expected, the mAb PeM19 (linear) recognized similarly the Prn1, Prn2, P.69 (Prn1) and the six PRN proteins (CI 95 %: 1.026- 1.063; CV = 3.55 %). The binding of PeM29 (conformational) followed the order: PRN1-sc-2 > Prn1, Prn2, P.69 (Prn1), PRN1-2, PRN1-lc-2 and PRN2-1 (CI 95 %: 0.809-0.855; CV = 4.42 %) > PRN2-lc-1 > PRN2-sc-1. Similarly, the binding of PeM1 (conformational) followed the order: PRN1-sc-2, PRN1- lc-2 (CI 95 %: 1.317-1.389; CV = 1.68 %) > Prn1, Prn2, P.69 Prn1, PRN2-1, PRN2-lc-1(CI 95 %: 0.979- 1.039; CV = 4.15 %) > PRN2-sc-1. The linear mAb PeM4 recognized the Prn molecules in the following order: PRN2-1 > PRN1-sc-2, PRN1-lc-2, PRN2-sc-1, PRN2-lc-1 (CI 95 %: 1.732-1.1.855; CV = 4.82 %) > PRN1-2, Prn2 (CI 95 %: 1.527-1.667; CV = 2.76 %) > Prn1 > P.69 (Prn1).
After thermal treatment at 95 °C, the recognition of P.69, Prn1, Prn2, PRN1-lc-2 and PRN2-1 was reduced as average 12- and 2.5-fold for PeM29 and the specific antiserum, respectively.
Sera from individuals vaccinated with the Adacel vaccine recognized similarly the Prn1, Prn2, P.69 (Prn1) and the PRN proteins.
]]> Immunogenicity of P.69, Prn1, Prn2 and the six PRN variants in Balb/c miceThe sera of immunized mice (all groups) exhibited a significant IgG levels (p < 0.001) in respect to mice who received placebo (data not shown). After the third dose, the mice immunized with PRN molecules showed higher levels of antibodies as compared to the natural Prn controls P.69, Prn1 and Prn2 (Figure 6A). Particularly, PRN1-lc-2 and PRN2-1 induced very highly significant IgG levels (p < 0.001) as compared to groups immunized with the Prn controls. As shown in figure 6B, the profile of anti-Prn1 immunoglobulin subclass (analyzed from pooled sera) was IgG1 > IgG2a > IgG2b > IgG3 = IgM = IgA in all immunized groups. The lowest values of the ratio IgG1/IgG2a (1.07) and IgG1/IgG2b (1.48) were detected in sera from mice immunized with PRN1-lc-2.
The sera induced by the pertactin controls P.69, Prn1, Prn2 and the two more immunogenic hybrid molecules, PRN1-lc-2 and PRN2-1, were further evaluated against linear peptides from the variable region R1 of Prn1, R1 of Prn2, the variable region R2 and the P4 peptide (IVKTGERQHGIHIQGSDP) sequence located in the N-terminus immunodominant domain. The region R2 was marginally recognized by 2 out of 9 sera from mice immunized with both P.69 and PRN1-lc-2 (data not shown). The sera from groups immunized with PRN1-lc-2, PRN2-1 and Prn2 recognized the variable region 1 of Prn1 and Prn2 with a significantly higher activity than sera from mice that received P.69 and Prn1 (Figure 7). Only the sera from mice immunized with PRN1-lc-2 recognized the peptide P4 (N-terminus region) with significant differences compared to P.69 and Prn1 control groups (Figure 7C). The immunoglobulin subclasses were also evaluated with peptides P1, P2 and P4. The profile was similar to the anti-Prn1 immunoglobulin subclass corresponding to the protein PRN1-lc-2 the lowest IgG1/IgG2a and IgG1/IgG2b ratio (data not shown).
DISCUSSION
The purpose of this study was to design and develop hybrid molecules of Prn1 and Prn2. The concept could be advantageous since a hybrid Prn could induce a more immunogenic/protective response and be more effective against different strains. Technologically, a hybrid molecule also could impact on reducing production costs by avoiding the separate production of the antigens. This strategy can also contribute to extend the effectiveness of a vaccine formulation in different regions and epidemiological settings.
Recent molecular epidemiological studies have demonstrated the circulation of strains expressing different types of Prn. In Europe and the Americas prevail strains harboring Prn2 [23-25] unlike regions in Asia where the displacement of Prn1 strains has been more gradual and both types of strains coexist [26]. In mouse models, Prn1 strains colonize advantageously mice devoid of immunity, unlike Prn2 strains that settle favorably in vaccinated animals, a fact consistent with the predominance of Prn1 strains in unvaccinated populations [27]. There is also involved the gap between vaccine and circulating strains, which adds fuel to the complexity of pertussis disease. A great proportion of vaccinated individuals with current vaccine (Prn1 strains) could be more susceptible to be infected with Prn2 strains, mainly, at waning immunity. Consequently, this could increase the number of trans-mitters/reservoirs of pertussis disease. In this regard, efforts should be made to update the current vaccines to circulating strains.
Considering these facts, PRN molecules were designed with the support of bioinformatics and structural analysis. The sequence examination of different variants of Prn from B. pertussis and B. parapertussis showed a polarized pattern of single amino acid substitutions towards the N-terminus region despite the high degree of similitude/similarity among them. The accumulation of mutations proximal to the amino end of the Prn sequence (first 300 aa.) is in close relation with previously observed immunological aspects. The N-terminus region was recently described as immunodominant, particularly in humans, with a 97 % of sera from infected and vaccinated individuals recognizing the first 110 aa.[28]. In addition, the first 230 aa. of this region contain dominant and subdominant helper CD4+ T cell epitopes [29]. These features highlight the importance of including the domain in the design of new molecules. Additionally, the structural prediction using I-TASSER showed the conserved C-ter-minus end folded in direction of the N-terminus end. This prediction is in agreement with previous epitopic mapping with monoclonal antibodies that suggested that the variable regions R1 and R2 are structurally close [19].
Considering the structural and sequence data, the concept of PRN from Prn1 and Prn2 was explored (see the Molecular design section in RESULTS). Six variants of PRN were cloned and expressed in E. coli. This host previously showed to be convenient for the expression and purification of several natural variants and deletion mutants of Prn [10]. Here, in agreement with the previous reports, the PRN and the control proteins (Prn1 and Prn2) were similarly expressed at high levels. For the PRN, the molecular size increase in 18 kDa had negligible influence in protein expression.
The six PRN, Prn1 and Prn2 were purified at levels of purity > 90 % using His-tagged affinity chromatography. Subsequently, the proteins were refolded and the proper conformation of the PRN proteins was evaluated by ELISA using mAbs against the discontinuous PeM1 and PeM29 epitopes. The mAbs PeM1 and PeM29 positively reacted with the refolded Prn1 (recombinant/E. coli) protein and the native P.69 (Prn1) purified from B. pertussis. The equivalence in binding activity for both mAbs suggests that the folding procedure was appropriate. Previous studies showed that Prn folds efficiently during the gentle subtraction of 6 M guanidine hydrochloride [10]. Here, similar results were obtained using 4 M urea.
]]> The six PRNs were also positively recognized by the discontinuous mAbs, although some variation was observed in respect to Prn1 and P.69 controls. Previous studies showed that the bindings of PeM1 and PeM29 are respectively reduced or abolished when deletions or point modifications are made in the first 110 residues of the amino end [22]. Here, the binding of PeM1 and PeM29 mAbs was unaffected or improved for the PRNs with the exception of PRN1- sc-2/PeM1 which binding was reduced by half. Only after thermal treatment at 95 °C a lack of binding was observed for both natural and hybrid molecules, suggesting an appropriate refolding of the PRNs. Additionally, the human sera from vaccinated individuals were able to recognize the PRNs without differences respect to Prn1, Prn2 and P.69. All the above results suggest that the refolded PRN molecules and natural Prn share a considerable number of epitopes.The immunogenicity of the PRNs was further evaluated in Balb/c mice. After the third dose, mice immunized with the PRN molecules exhibited equal or higher levels of IgG respect to mice immunize with Prn controls. Particularly, the IgG levels induced by PRN1- lc-2 and PRN2-1 were highly significant (p < 0.001) with the presence of the subtypes IgG1, IgG2a and IgG2b. Specifically in mice, IgG2a and IgG2b have been involved in cellular host responses to bacterial infections [30, 31]. Furthermore, in humans, the anti-Prn antibodies have been shown as crucial, mediating B. pertussis phagocytosis [32]. Additionally, the PRN1-lc-2 and PRN2-1 molecules elicited a significant response against relevant epitopes associated to protection, such as the variable region R1 [6] and the immunodominant N-terminal end domain [28].
Noteworthy, the immunological relevance of these results must be verified in functional studies. Preliminary results showed significantly higher survival (p < 0.001) in OF1 mice immunized with the proteins PRN1-lc-2 or Prn2-1 (alone) against the virulent strain 18323 (Prn7) respect to non immunized mice in the modified intracranial challenge assay (data not shown).
The differences in IgG titers induced by a natural Prn and PRN molecules may be due to several interrelated factors. An aspect that may be influencing the antibody response is the duplication of the region V81-S274 (Ref PRN2) in the hybrid proteins. As mentioned before, several helper CD4+ T cell epitopes have been indentified in this region. Probably, the duplication of these epitopes improves their presentation in the context of MHC-II promoting the activation of effector B cells. It is suggestive that the mAb PeM4 recognizes favorably the GGFGPGGFGP epitope (R1 region) in 5 out of 6 PRNs in respect to pertactin controls. The hybrid pertactin molecules have two variable R1 regions and, therefore, two copies of this epitope. As shown by structural analyses, the distance (> 40 Å) between the duplicated epitope facilitates the simultaneous binding of two antibody molecules or B cell receptor molecules, favoring B cell signaling. Interestingly, the GGFGPGGFGP epitope was also better recognized in Prn2 than in Prn1/P.69 proteins. Since the R1 region in Prn2 has a longer loop compared to Prn1 [33], the level of surface exposure of this epitope could also be favoring its recognition by PeM4. Also in agreement to this, mice immunized with Prn2 elicited higher levels of IgG against the region 1 than the Prn1/P.69 proteins, suggesting that also the level of exposure of this region could be promoting B cell activation. On the other hand, it has been proposed that the C-terminal end of pertactin could play a concealing role over N-terminal end [22]. Due to the design of the PRN molecules, the GGFGPGGFGP epitope that localized closer to the N-terminal could be more accessible as a consequence of the remoteness from the C-terminal domain. Similarly, this statement could be fulfilled in the PRN1-lc-2 protein which elicits significant levels of IgG against an N-terminal epitope. In relation with all above, the structural arrangement specific to each protein may play a role in epitope accessibility and consequently in the better titers and quality of the antibody responses observed for PRN molecules.
Further studies aimed at evaluating the protective capabilities of the hybrid molecules are currently in progress. These hybrid molecules should be studied in the context of combined formulations containing other pertussis components such as pertussis toxoid and filamentous hemagglutinin, and also the diphtheria tetanus toxoid. The different components in a combined formulation can modulate the induction of an effective immune response against pertussis.
In conclusion, here we report for the first time the genetic engineering of PRN molecules from the B. pertussis Prn. These hybrid proteins constitute promising candidates to further in vivo immunization studies.
REFERENCES
1. Bordet J, Gengou O. Le microbe de la coqueluche. Ann Inst Pasteur (Paris). 1906;20:731-41.
]]>2. Willems RJL, Mooi FR. From whole cell to acellular pertussis vaccines. Rev Med Microbiol. 1996;7:13-21.
3. Mattoo S, Cherry JD. Molecular pathogenesis, epidemiology, and clinical manifestations of respiratory infections due to Bordetella pertussis and other Bordetella subspecies. Clin Microbiol Rev. 2005;18(2):326-82.
4. He Q, Mertsola J. Factors contributing to pertussis resurgence. Future Microbiol. 2008;3(3):329-39.
5. Mooi FR. Bordetella pertussis and vaccination: the persistence of a genetically monomorphic pathogen. Infect Genet Evol. 2010;10(1):36-49.
6. King AJ, Berbers G, van Oirschot HF, Hoogerhout P, Knipping K, Mooi FR. Role of the polymorphic region 1 of the Bordetella pertussis protein pertactin in immunity. Microbiology. 2001;147(Pt 11):2885-95.
7. Komatsu E, Yamaguchi F, Eguchi M, Watanabe M. Protective effects of vaccines against Bordetella parapertussis in a mouse intranasal challenge model. Vaccine. 2010;28(27):4362-8.
8. Aminian M, Sivam S, Lee CW, Halperin SA, Lee SF. Expression and purification of a trivalent pertussis toxin-diphtheria toxin-tetanus toxin fusion protein in Escherichia coli. Protein Expr Purif. 2007;51(2):170-8.
9. Jinyong Z, Xiaoli Z, Weijun Z, Ying G, Gang G, Xuhu M, et al. Fusion expression and immunogenicity of Bordetella pertussis PTS1-FHA protein: implications for the vaccine development. Mol Biol Rep. 2011;38(3):1957-63.
10. Hijnen M, van Gageldonk PG, Berbers GA, van Woerkom T, Mooi FR. The Bordetella pertussis virulence factor P.69 pertactin retains its immunological properties after overproduction in Escherichia coli. Protein Expr Purif. 2005;41(1):106-12.
11. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23(21):2947-8.
12. Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. UCSF Chimera--a visualization system for exploratory research and analysis. J Comput Chem. 2004;25(13):1605-12.
13. Zhang Y. I-TASSER server for protein 3D structure prediction. BMC Bioinformatics. 2008;9:40.
14. Emsley P, Charles IG, Fairweather NF, Isaacs NW. Structure of Bordetella pertussis virulence factor P.69 pertactin. Nature. 1996;381(6577):90-2.
15. Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. 2nd ed.New York: Cold Spring Harbor Laboratory Press; 1989.
16. Kasuga T, Nakase Y, Ukishima K, Takatsu T. Studies on Haemophilis pertussis. III. Some properties of each phase of H. pertussis. Kitasato Arch Exp Med. 1954;27(3):37-47.
17. Imai Y, Matsushima Y, Sugimura T, Terada M. A simple and rapid method for generating a deletion by PCR. Nucleic Acids Res. 1991;19(10):2785.
]]>18. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680-5.
19. Hijnen M, Mooi FR, van Gageldonk PG, Hoogerhout P, King AJ, Berbers GA. Epitope structure of the Bordetella pertussis protein P.69 pertactin, a major vaccine component and protective antigen. Infect Immun. 2004;72(7):3716-23.
20. Castellanos-Serra L, Ramos Y, Huerta V. An in-gel digestion procedure that facilitates the identification of highly hydrophobic proteins by electrospray ionization-mass spectrometry analysis. Proteomics. 2005;5(11):2729-38.
21. Oliver DC, Huang G, Nodel E, Pleasance S, Fernandez RC. A conserved region within the Bordetella pertussis autotransporter BrkA is necessary for folding of its passenger domain. Mol Microbiol. 2003;47(5):1367-83.
22. Hijnen M, de Voer R, Mooi FR, Schepp R, Moret EE, van Gageldonk P, et al. The role of peptide loops of the Bordetella pertussis protein P.69 pertactin in antibody recognition. Vaccine. 2007;25(31):5902-14.
]]>23. Mosiej E, Augustynowicz E, Zawadka M, Dabrowski W, Lutynska A. Strain variation among Bordetella pertussis isolates circulating in Poland after 50 years of whole-cell pertussis vaccine use. J Clin Microbiol. 2011;49(4):1452-7.
24. Borisova O, Kombarova SY, Zakharova NS, van Gent M, Aleshkin VA, Mazurova I, et al. Antigenic divergence between Bordetella pertussis clinical isolates from Moscow, Russia, and vaccine strains. Clin Vaccine Immunol. 2007;14(3):234-8.
25. Bottero D, Gaillard ME, Fingermann M, Weltman G, Fernandez J, Sisti F, et al. Pulsed-field gel electrophoresis, pertactin, pertussis toxin S1 subunit polymorphisms, and surfaceome analysis of vaccine and clinical Bordetella pertussis strains. Clin Vaccine Immunol. 2007;14(11):1490-8.
26. Zhang L, Xu Y, Zhao J, Kallonen T, Cui S, Hou Q, et al. Effect of vaccination on Bordetella pertussis strains, China. Emerg Infect Dis. 2010;16(11):1695-701.
27. van Gent M, van Loo IH, Heuvelman KJ, de Neeling AJ, Teunis P, Mooi FR. Studies on Prn variation in the mouse model and comparison with epidemiological data. PLoS One. 2011;6(3):e18014.
28. Hijnen M, He Q, Schepp R, Van Gageldonk P, Mertsola J, Mooi FR, et al. Antibody responses to defined regions of the Bordetella pertussis virulence factor pertactin. Scan J Infect Dis. 2008;40(2):94-104.
29. Stenger RM, Poelen MC, Moret EE, Kuipers B, Bruijns SC, Hoogerhout P, et al. Immunodominance in mouse and human CD4+ T-cell responses specific for the Bordetella pertussis virulence factor P.69 pertactin. Infect Immun. 2009;77(2):896-903.
30. Skerry CM, Mahon BP. A live, attenuated Bordetella pertussis vaccine provides long-term protection against virulent challenge in a murine model. Clin Vaccine Immunol. 2011;18(2):187-93.
31. Nimmerjahn F, Ravetch JV. Fcgamma receptors: old friends and new family members. Immunity. 2006;24(1):19-28.
32. Hellwig SM, Rodriguez ME, Berbers GA, van de Winkel JG, Mooi FR. Crucial role of antibodies to pertactin in Bordetella pertussis immunity. J Infect Dis. 2003;188(5):738-42.
33. Mooi FR, He Q, van Oirschot H, Mertsola J. Variation in the Bordetella pertussis virulence factors pertussis toxin and pertactin in vaccine strains and clinical isolates in Finland. Infect Immun. 1999;67(6):3133-4.
Received in March, 2013.
Accepted in December, 2013.
]]>
Diógenes Quintana-Vázquez. Centro de Ingeniería Genética y Biotecnología, CIGB Ave. 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba. E-mail: diogenes.quintana@cigb.edu.cu.
]]>
{kind=link}
{kind=link}
{kind=link}
{kind=link}