RESEARCH
Cloning and study of new DNA methyltransferase M.AluBI modifying adenine in a recognition site AGCT
Clonaje y estudio de una nueva metiltransferasa de ADN M.AluBI que modifica la adenina en el sitio de reconocimiento AGCT
]]>
Vladimir Dedkov, Danila Gonchar, Murat Abdurashitov, Svetlana Udalyeva, Lyubov Urumceva, Valery Chernukhin, Elena Shiryaeva, Sergey Degtyarev
LaborSibEnzyme Ltd., Ak. Timakov str. 2/12, Novosibirsk, 630117, Russia.
ABSTRACT
A fragment of Arthrobacter luteus B DNA carrying the gene of new DNA methyltransferase M.AluBI was cloned and expressed in Escherichia coli. The recombinant plasmid pM.AluBI-16 contains the M.AluBI gene (1515 bp in length), corresponding to a protein of 504 amino acid residues. The amino acid sequence analysis showed that M.AluBI could be an adenine-(N6)-DNA methyltransferase. A recombinant strain was grown up and the enzyme was purified by a consecutive chromatography on P-11 Phosphocellulose, Heparin-Sepharose, Sephacryl S-200 and Hydroxyapatite. M.AluBI specificity was determined by the original method based on blocking of restriction endonucleases cleavage of overlapped sites and on computer modeling. It was first shown that AluBI MTase modifies the adenine residue with formation of 5´-(m6A)GCT-3´ as opposed to its prototype, M.AluI, producing 5´-AG(m5C)T-3´. A comparative sensitivity analysis of different, well known restriction endonucleases to the methylation by M.AluBI and M.AluI was done using λ and T7 phage DNA. The newly acquired data on methylation sensitivity cold be useful for conducting experiments on DNA digestion with restriction endonucleases, and especially with the particular cleavage sensitivity pattern generated with the M.AluBI methyltransferase enzyme.
Keywords: gene cloning, enzyme isolation, bacterial DNA methyltransferase, enzyme specificity, restriction endonuclease, methylation sensitivity.
RESUMEN
Se clonó un fragmento de ADN de Arthrobacter luteus B que contiene el gen de una nueva metiltransferasa de ADN, y se expresó en Escherichia coli. Se obtuvo el plásmido recombinante pM.AluBI-16, que contiene el gen M.AluBI (1515 pb), codificante para una proteína de 504 aminoácidos. El análisis comparativo de la secuencia aminoacídica mostró que M.AluBI pudiera ser una metiltransferasa de ADN adenina-(N6). Se cultivó la cepa recombinante y se purificó la enzima, mediante pasos consecutivos de cromatografía en matrices de Fosfocelulosa P-11, Sefarosa-heparina, Sephacryl S-200 e Hidroxiapatita. La especificidad de M.AluBI se determinó mediante el método original de bloqueo del corte de enzimas de restricción en sitios solapados, y mediante la modelación computacional. Se demostró que la AluBI metiltransferasa modifica el residuo de adenina y genera un producto 5´-(m6A)GCT-3´, a diferencia del producto 5´-AG(m5C)T-3´ obtenido por la acción de su prototipo M.AluI. Posteriormente se analizó comparativamente la sensibilidad a la metilación por M.AluBI y M.AluI, de diferentes endonucleasas de restricción conocidas en el ADN de los bacteriófagos λ y T7. Los nuevos datos de sensibilidad a la metilación obtenidos pueden ser útiles en experimentos de digestión de ADN con enzimas de restricción, en particular los obtenidos con el patrón de sensibilidad generado por la metiltransferasa M.AluBI.
Palabras clave: clonaje de genes, aislamiento enzymático, metiltransferasa de ADN bacteriana, especificidad enzimática, endonucleasa de restricción, sensibilidad a la metilación.
INTRODUCTION
]]> DNA methylation is the most common form of DNA modification in prokaryotic and eukaryotic genomes. DNA methyltransferases (MTases) transfer the methyl group from S-adenosyl-L-methionine (SAM) to cytosine or adenine in double-stranded DNA. The modified bases are 5-methylcytosine (5mC or m5C), N4-methylcytosine (N4mC or m4C) or N6-methyladenine (N6mA or m6A). The MTases differ in their recognition sequence and the base to be modified within this site [1].Most known MTase specificities are exclusively found in bacteria. These enzymes partly belong to restriction-modification (RM) systems. About 350 prototypes of RM systems and thousands of their analogs are known at present time [2]. Each RM system consists of restriction endonuclease (restrictase) and MTase. Both cognate enzymes recognize the same site. The endonuclease cuts a recognition site thus protecting bacterial cell from a foreign DNA invasion. MTase methylates the recognition site in host DNA and protects it from cleavage with a cognate endonuclease [3, 4]. Endonucleases of various bacteria recognizing the same site are called isoschizomers. New MTases are a perspective tool for a study of DNA-protein interactions. One of these studies is a determination of endonucleases sensitivity to a new type of methylation.
There is a well known RM system AluI from Arthrobacter luteus with a recognition site AGCT. M.AluI modifies this site with formation AG(m5C)T [5]. Recently, we described a new RM system AluBI from Arthrobacter luteus B with the same substrate specificity AGCT [6]. RM-systems AluI and AluBI must differ in DNA methylation because AluBI cuts DNA modified by M.AluI [6]. This work was aimed to cloning of M.AluBI and the study of the recombinant enzyme properties and substrate specificity. A comparative study of sensitivity was performed of different endonucleases to DNA methylation by M.AluBI and M.AluI.
MATERIALS AND METHODS
Enzymes, DNA and reagents
M.AluI DNA methyltransferase and S-adenosyl-L-methionine (SAM) were from New England Biolabs Inc, USA. Restriction endonucleases, alkaline phosphatase (calf intestinal) and T4 DNA ligase as well as pUC19, phage l(dam-, dcm-) and T7 DNAs, ATP, reaction buffers were obtained from Sibenzyme Ltd, Russia. Lysozyme was from Helicon, Russia. Bovine serum albumin (fraction V) (BSA) was from Americanbio, USA.
Strains and culture medium
Escherichia coli RR1 (ΔmcrC-mrr recA+) was from New England Biolabs Inc, USA. Arthrobacter luteus B was from Sibenzyme Ltd, Russia. L-broth contains 10 g/L Tryptone (Organotechnie, France), 5 g/L Yeast extract (Organotechnie, France), 5 g/L NaCl, 0.5 g/L MgCl2, 1 mg/L thiamin. For plates L-broth was supplemented with 15 g/L agar (C.T. Roeper GmbH, Germany). Ampicillin was added up to 100 mg/mL and streptomycin up to 25 mg/mL.
Transformation and colony screening procedures
]]> A. luteus B was grown in flasks containing 300 mL L-broth at 30 °C, with 130 rpm for 40 h. Six micrograms of A. luteus B DNA, isolated as described [7], was separately digested in 200 mL of reaction mixtures with 10-20 units of SphI (GCATG^C) or BstNSI (RCATG^Y) for 1 h at 37 °C. The DNA from each digest was purified by phenol extraction and precipitated with ethanol. Hybrid plasmids of pUC19 were constructed by ligation of 2.5 mg of SphI-digested A. luteus B DNA and 5.0 mg of BstNSI-digested A. luteus B DNA with 3.0 mg phosphatase-treated SphI-digested pUC19 by using 2000 units of T4 DNA ligase. The reaction was allowed to proceed for 16 h at 4 °C in 20 mL of 50 mM Tris-HCl, pH 7.8, 10 mM MgCl2, 10 mM DTT, 1 mM ATP. The ligated DNA was precipitated with ethanol and dissolved in 12 mL of water.Electroporation was performed in a mixture of 2.5 mg of ligation product to 50 mL of approximately 3.0 × 109 RR1 cells, by using an EasyJet Prima electroporator according to the manufacturer’s instructions (EquiBio, UK). After electroporation, the mixture was added to 1.0 mL of L-broth and incubated for 1 h at 37 °C. For calculation of transformants, 10 mL of the culture were plated on L-agar with ampicillin. Eighty colonies grew at 37 °C after 16 h. Thus about 8000 recombinant clones were obtained. The genomic library (1.0 mL) was grown in 100 mL of L-broth with ampicillin for 16 h at 37 °C with shaking.
Total plasmid DNA of 8000 recombinant clones from 100 mL culture was isolated by using the Qiagen Plasmid Maxi Kit (Germany) according to the manufacturer’s instructions. Ten micrograms of plasmid DNA preparation was digested with 50 U of AluBI for 4 h at 37 °C, precipitated with ethanol and dissolved in 15 mL of water. Following the protocols [7], digested DNA was incubated with 3.0 × 107 RR1 cells that had been treated with CaCl2. Cells were plated on L-agar containing ampicillin and 16 clones were obtained after 16 h at 37 °C. Each clone was grown in L-broth with ampicillin and plasmid DNA was isolated by Qiagen Plasmid Miniprep Kit (Germany). DNAs from 16 clones were cut with AluBI and analyzed by electrophoresis in 1 % agarose gel. The 16 DNA preparations were resistant against AluBI digestion and carried an insert about 8 kb in length. One of DNAs was cut with Acc65I, ligated with Acc65I-digested pUC19 DNA and used for RR1 retransformation. The obtained clone was named E. coli N16 (pM.AluBI).
DNA sequencing
The sequencing of DNA was carried out with the ABI 3130xI Genetic Analyzer device (Applied Biosystems, USA).
Production and purification of DNA methyltransferase M.AluBI from Escherichia coli N16 (pM.AluBI)
E. coli N16 (pM.AluBI) cells were grown till stationary phase in L-broth with ampicillin in a 20-L Vessel (New Brunswick Scientific, USA) at 30 °C with aeration for 8 h. Cells were harvested at 8000 × g and stored at -20 °C.
The enzyme was purified at 4 °C by using Buffer A (10 mM Tris-HCl, pH 7.5; 0.1 mM EDTA, 7 mM β-mercaptoethanol) and Buffer B (10 mM Potassium Phosphate, pH 7.2; 0.1 mM EDTA; 7 mM β-mercaptoethanol). Harvested cells (100 g) were suspended in 500 mL of Buffer A with 0.2 M NaCl, 5 % glycerol, 0.5 % Triton® X-100 (non-ionic detergent, Sigma) and 0.1 mM PMSF (proteases inhibitor). Cell suspensions (100 ml each) were sonicated on Soniprep 150 (MSE, UK) with a nozzle 2-cm in diameter and receiving 5 maximal impulses for 1 min with cooling in ice for 1 min. Cell debris was removed by centrifugation at 20 000 × g for 1 h. The supernatant was loaded onto a Phosphocellulose P11 (Whatman, UK) of 300 mL bed volume and eluted with 3 L of 0.2 to 0.6 M NaCl gradient in Buffer A for 15 h.
Fractions were assayed for M.AluBI activity as described below. The activity-containing fractions were pooled, dialyzed against 3 L of Buffer A with 0.05 M of NaCl for 4 h and loaded onto a Heparin-Sepharose (Bio-Rad, USA) of 50 mL bed volume column. Protein was eluted with 1 L of a 0.05-0.5 M NaCl gradient in Buffer A. Fractions containing activity were pooled, loaded onto a Sephacryl S-200 (Bio-Rad, USA) of 1 L bed volume column and eluted with 1 L of 0.8 M NaCl in Buffer A. Subsequently, fractions containing activity were loaded onto a Hydroxyapatite (Bio-Rad, USA) of 50 mL bed volume column and eluted with 1 L of a 0.01-0.2 M K-phosphate gradient, pH 7.2 in Buffer B. Finally, fractions containing activity were pooled, concentrated by a dialysis against 1 L of Buffer A with 50 % glycerol, 0.05 M of NaCl and stored at -20 ˚C.
DNA Methyltransferase activity assay
]]> The assay based on a protection of methylated DNA from cognate restriction endonuclease. M.AluBI sample was incubated in 50 mL with 50 mg/mL λDNA (dam-, dcm-), 1 mM SAM, 33 mM Tris-acetate, pH 7.9, 1 mM EDTA, 66 mM potassium acetate 1 mM DTT, 0.1 mg/mL BSA in a well of 96 Well Microplate (Medpolymer, Russia) at 37 °C for 1 h. Then, the microplate was warmed up on a bath at 65 °C for 10 min to inactivate the enzyme. To cleave not modified DNA, the mixture was supplemented with 5 mL of 1000 U/mL AluBI restriction endonuclease diluted with 10 mM Tris-HCl pH 7.6, 50 mM KCl, 0.1 mM EDTA, 200 mg/mL BSA, 1 mM DTT and 100 mM magnesium acetate. After incubation at 37 °C for 1 h, the mixture was analyzed by electrophoresis in 0.8 % agarose gel as described [7]. One unit of M. AluBI activity methylated 1 mg of lDNA in 1 h blocking the activity of AluBI restriction endonuclease.DNA methylation with M.AluBI and M.AluI
The reaction mixture was prepared in a 2 mL Eppendorf tube, containing 0.4 mg/mL lor T7 DNA, 10 mM SAM, 33 mM Tris-acetate pH 7.9, 1 mM EDTA, 66 mM potassium acetate 1 mM DTT, 0.1 mg/ mL BSA and 100 U/mL M.AluBI or M.AluI enzyme, in a total volume of 1.0 mL, and incubated at 37 °C for 20 h. Then, 0.1 mL of 10 % SDS (Sigma) and 0.2 mL of 3 M KCl were added. The mixture was warmed at 65 °C for 10 min to dissolve SDS-precipitate and cooled at 0 °C for 15 min for precipitate formation. The supernatant, pooled after centrifugation at 10 000 × g for 3 min, was mixed with 0.6 volume of isopropanol. DNA pellet was rinsed with 80 % ethanol, dried on air and dissolved in 0.7 mL of TE buffer (10 mM Tris-HCl pH 8.0, 1 mM EDTA) to the 0.5 mg/mL concentration.
DNA cleavage with restriction endonucleases and electrophoresis in agarose gel
Native or M.AluBI- and M.AluI-methylated lor T7 DNA were cleaved in 50 μL of the reaction mixture containing 50 mg/mL DNA, 0.1 mg/mL BSA, appropriate SE buffer, 125 U/mL restriction endonuclease in a well of 96 Well Microplate, incubated for 2 h at the temperature recommended by the manufacturer (Sibenzyme Ltd). The reaction was stopped by addition 10 μL of 0.25 M Na-EDTA, pH 8.5, 50 % sucrose and 0.5 % bromphenol blue. The Microplate was warmed up on a bath at 65 °C for 10 min and cooled on ice. The electrophoresis was carried out in 0.8 % of LE agarose (Segenetic) in TAE buffer with 0.5 mg/mL ethidium bromide (Sigma) at 5 V/cm for 2.5 h as described [7]. The fluorescence of DNA was revealed on the Herolab GmbH device.
Determination of DNA methyltransferase specificity
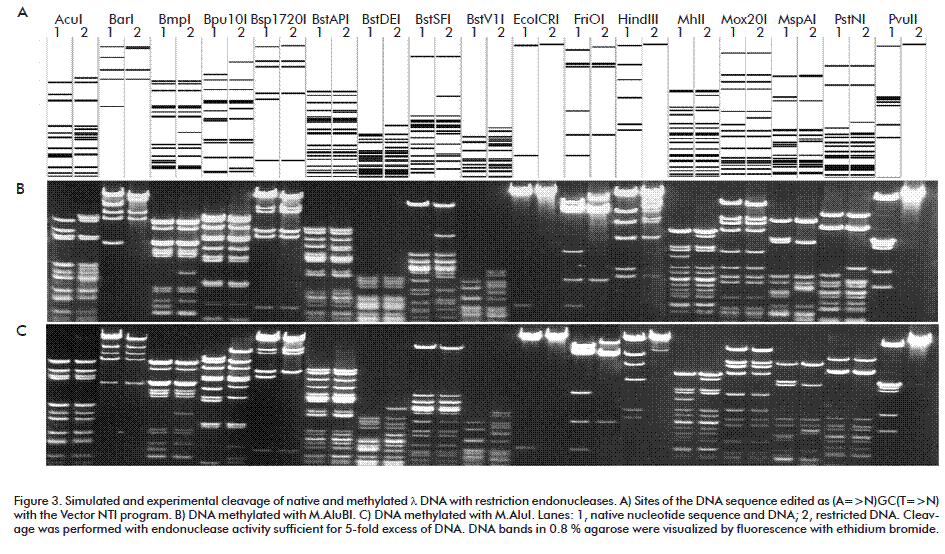
M.AluBI recognition sequence and its methylated basis were defined using methylation sensitivity of restriction endonucleases [2] according to Ded-kov [8]. As described above T7 DNA, as native and M.AluBI-methylated, were cut by restriction endonucleases AluI (AG^CT), AluBI (AG^CT), Tru9I (T^TAA), Fsp4HI (GC^NGC), BstC8I (GCN^NGC) and Bst2BI (CACGAG(-5/-1)). Then, DNA fragments were analyzed by electrophoresis in 0.8 % agarose. Beforehand, blocking of restriction endonucleases by the methylation was modeled to reveal an overlapping of the methylation and the cleavage. Methylation of (mA)GCT was simulated as editing T7 DNA sequences by replacements AGCT with NGCN and designated (A=>N)GC(T=>N). Then, simulated cleavage of native and edited DNA by restriction endonucleases and electrophoresis was performed with Vector NTI program. In the program, a restriction endonuclease didn’t cut the recognition site if any of bases was replaced by N. Thus the methylated site was examined by cleavage with restriction endonucleases. M.AluBI specificity was determined by the analysis of experi-mental and simulated results.
RESULTS AND DISCUSSION
Selection of clones carrying the M.AluBI and DNA sequencing
]]> MTase clones were selected by the resistance of recombinant DNA to the cognate restriction endonuclease according to [9]. A genomic library of A. luteus B DNA was obtained in pUC19/Sph I vector DNA as 8000 E. coli RR1 transformants. A total DNA of hybrid plasmids was digested with AluBI and used for additional RR1 transformation. Plasmid DNAs from 16 clones were cut with AluBI and analyzed by electrophoresis in 1 % agarose gel. All these plasmids were protected from AluBI restriction endonuclease and contained ~8.0-kb insertions of A. luteus B DNA including the M.AluBI gene. Then 8.0-kb insertion was cut with Acc65I and 2101 bp DNA fragment with M.AluBI gene was subcloned into pUC19/Acc65I vector DNA. The obtained clone was named E. coli N16 (pM.AluBI). Final plasmid DNA, pM.AluBI-16, was sequenced. A restriction map of this recombinant plasmid with ORF of M.AluBI gene is shown in Figure 1.The Acc65I-fragment of A. luteus B DNA cloned in pUC19 contained 2101 base pairs with a single open reading frame (ORF) corresponding to M. AluBI of 504 amino acids (in single-letter): MTFTTSWRARIFEEAELLPTDLISDLSIGEVSVLYEYTHAIADADARKTNGQFFTPDDVASFMAGFADRFPRGRWLDPC
AGIGNLSWHLIASQEDPEEFLLERMLLSDTDELALLVARALLTAFQRRHLNLFHEIEENFVVFDFLSVSDSGTSKIGALAAIPSHDFVIVNPPYLATKGRDSRFETAESSDLYAYF
LENVIKTSRGFISVTPQSFTNAKKFMGLRSLLLRSYSNLEILCFDNIPGNLRGVKFGSRNSNTANSIRAAITVALPQPGVPSITSLMRWKGTERQRLFAEVERFKSHVPLTAEFFP
KVSTVFEDLYRWTVELPRLGSLCQRTETEFPLHVPSAPRYFIPALKSPVARSSQRTLYPNSAARDRAYLLINSSLMYWWWRVRDGGMTLSQETLLSMPLPDFAVRDDLVTRLE
LSEETNKVYKLNAGVSQENVKHSPELIADLNAHIVPIHAAPLLLTHLNSEFAQFEYLGRADDAASSARAIVANGAS.
The nucleotide and amino acid sequences of M.AluBI were deposited in NCBI GenBank (http://www.ncbi.nlm.nih.gov/nuccore/LN869917.1). With the aid of Protein Blast (http://blast.ncbi.nlm.nih.gov/ Blast.cgi) the putative conserved motifs of M.AluBI were detected in AdoMet_MTases superfamily with N6_MTase domain hit. This result confirms the data obtained before [6] and shows that M.AluBI belongs to amino-MTases and modifies the site differently than M.AluI (AG(m5C)T) [5]. The structure of MTases is characterized by 9 amino-acid motifs [10]. The main motifs are motif I (F-G-G) for binding SAM and motif IV (earlier – II) for catalysis: (P/F)PC in cytosine (C5) - MTases (C-MTases) and PPY in adenine (N6) - and cytosine (N4) amino MTases. Thus, as the structure of MTases which form m4C and m6A is very similar, а purification of the enzyme is necessary to determine M.AluBI specificity.
M.AluBI purification and characterization
To isolate M.AluBI enzyme preparation, E.coli N16 (pM.AluBI) cells were grown in L-broth with ampicillin. The enzyme was purified from cellular extract by subsequent chromatography on Phosphocellulose P11, Heparin-Sepharose, Sephacryl S-200 and Hydroxyapatite as described at Materials and methods. The traditional chromatography sequence allowed to obtain enzymatic-pure preparation of M.AluBI with activity 10 U/μL, which was suitable for works in molecular biology. Sufficient ionic strength and Phosphocellulose P11 were used to dissociate nucleoprotein complexes and adsorb many DNA-specific enzymes on this resin. The majority of cellular proteins wasn’t adsorbed, it allowed to use rather small column already in the first chromatography.
The substrate specificity of M.AluBI was confirmed by an inability of restriction endonucleases to hydrolyze some sites and a computer simulation of this hydrolysis as described earlier [8] and indicated in Materials and methods. Figure 2 shows that the methylation of T7 DNA with M.AluBI blocks DNA hydrolysis with AluI as well as AluBI restric-tion endonucleases (AGCT). Moreover, M.AluBI blocked a digestion with Tru9I in the case of overlapping with an external adenine in the site TTAA. Thus M.AluBI modifies DNA producing (m6A)GCT. At the same time AluBI-methylation didn’t block Fsp4HI (GC^NGC) and BstC8I (GCN^NGC) which are sensitive to m5C [11, 12]. As shown in Figure 2, Bst2BI is sensitive to the methylation with M.AluBI and doesn’t cleave site CACG(m6A)G.
]]> Thus, the results show that M.AluBI produces 5´- (m6A)GCT-3´ and belongs to adenine-(N6)-DNA methyltransferases (EC 2.1.1.72) [1]. M.AluBI possesses a new specificity among known analogs [2] and may be used in genetic engineering and molecular biology.Methylation sensitivity of restriction endonucleases
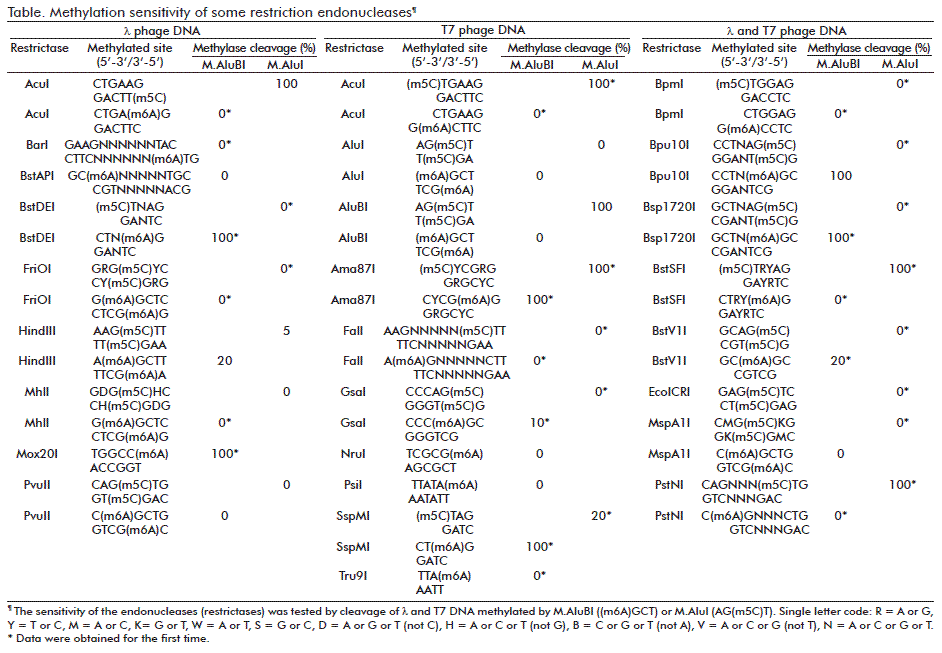
An essential characteristic of restriction endonucleases is methylation sensitivity of these enzymes. A number of endonucleases were tested for methylation sensitivity with land T7 DNAs methylated by M.AluBI or M.AluI with formation of (m6A)GCT or AG(m5C)T, respectively. M.AluBI differs from the similar enzymes such as M.AluI forming AG(m5C)T [5], or M.Mma5219II [2] and M.EsaBC1I [13] which form AG(m4C)T-3´. The model and experimental re-sults are shown in Figure 3 and Figure 4 and are interpreted in the Table. In some cases, the methylation of particular sequences was simulated as described [12].
Some results shown in the Table confirm known data about methylation sensitivity of restriction endonucleases [2]. Other results were obtained for the first time (* noted cleavage %). Methylation sensitivity is one of characteristics of restriction endonucleases [2]. In particular, this property is used in studying cancer. The obtained data expand possibilities of restriction endonucleases for studying DNA methylation.
CONCLUSIONS
The gene of a new DNA methyltransferase AluBI from bacterial strain Arthrobacter luteus B was cloned in E. coli. A recombinant M.AluBI enzyme preparation was isolated and it was shown that this MTase modified the adenine residue in the recognition sequence AGCT producing 5´-(m6A)GCT-3´. M.AluBI differs from the similar enzymes such as M.AluI forming AG(m5C)T or M.Mma5219II and M.EsaBC1I [13] which form AG(m4C)T-3’. The new data on sensitivity of different restriction endonucleases to M.AluBI- and M.AluI-methylation have been obtained.
ACKNOWLEDGEMENTS
This work was carried out with financial support from the Ministry of Education and Science of the Russian Federation under the Agreement No. 14.576.21.0077 of 06.11.2014 (the unique RFMEFI57614X0077 identifier), signed within the Federal Target Program “Research and Development in Priority Directions of Development of a Scientific and Technological Complex of Russia for 2014-2020”.
]]>CONFLICT OF INTEREST STATEMENT
The authors declare that there are no conflicts of interest.
REFERENCES
1. Cheng X, Blumenthal RM. (Eds.). S-Adenosylmethionine-Dependent Methyltransferases. Structures and Functions: Hong Kong: World Scientific Publishing Co. Pte. Ltd.; 1999.
2. REBASE. REBASE - The restriction enzyme database; 2015 [cited 2015 Aug 18]. Available from: http://rebase.neb.com
3. Arber, W, Dussoix, D. Host specificity of DNA produced by Escherichia coli. I. Host controlled modification of bacteriophage lambda. J Mol Biol. 1962;5:18-36.
4. Boyer HW. DNA restriction and modification mechanisms in bacteria. Annu Rev Microbiol. 1971;25:153-76.
5. Kramarov VM, Smolyaninov VV. DNA methylase from Arthrobacter luteus screens DNA from the action of site-specific endonuclease AluI. Biokhimiia. 1981;46(8):1526-9.
6. Chernuhin VA, Boltengagen AA, Tarasova GV, Dedkov VS, Degtjarjov SH. New restriction endonucleasa AluBI from Arthrobacter luteus B - isoschizomer AluI, tolerant to presence 5-methilcytosin in the recognition site AGCT. Vestnik biotehnologii i fiziko-himicheskoj biologii imeni Ju. A. Ovchinnikova. 2007;3(1):21-7.
7. Sambrook J, Fritsch EF, Maniatis, T. Molecular Cloning. A laboratory manual. 2nd ed. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press; 1989.
8. Dedkov VS. Defining specificity of DNA methyltransferase M. Bsc4I in cellular lysate by blocking restriction endonucleases and computer modeling. Mol Gen Microbiol Virol. 2009;24(3):114-20.
]]>9. Mann MB, Rao RN, Smith HO. Cloning of restriction and modification genes in E. coli: the HhaII system from Haemophilus haemolyticus. Gene. 1978;3:97-112.
10. Malone T, Blumenthal RM, Cheng X. Structure-guided analysis reveals nine sequence motifs conserved among DNA amino-methyltransferases, and suggests a catalytic mechanism for these enzymes. J Mol Biol. 1995;253:618-32.
11. Chmuzh EV, Kashirina YG, Tomilova YE, Chernukhin VA, Okhapkina SS, Gonchar DA, et al. The Fsp4HI restriction-modification system: Gene cloning, comparison of protein structures, and biochemical properties of recombinant DNA methyltransferase M.Fsp4HI. Mol Biol. 2007;41(1):37-43.
12. Dedkov VS. Novel M.BstC8I methyltransferase forms 5’-G(m5C)NNGC-3’. Investigation of restriction endonuclease sensitivity to M.BstC8I methylation. Mol Genet Microbiol Virol. 2012;27:40-7.
13. Clark TA, Murray IA, Morgan RD, Kislyuk AO, Spittle KE, Boitano M, et al. Characterization of DNA methyltransferase specificities using single-molecule, real-time DNA sequencing. Nucleic Acids Res. 2012;40(4):e29.
]]>
Received in September 2015.
Accepted in December 2015.
Danila Gonchar. LaborSibEnzyme Ltd., Ak. Timakov str. 2/12, Novosibirsk, 630117, Russia. E-mail: gonchar@sibenzyme.ru.
]]>{kind=link}
{kind=link}
{kind=link}
{kind=link}