RESEARCH
A homologous DNA recombination pathway alternative to RecFOR and RecBCD induced by gamma irradiation in Salmonella typhimurium
Ruta de recombinación homóloga del ADN alternativa a RecFOR y RecBCD inducida por radiación gamma en Salmonella typhimurium
]]> Elizabeth Cuétara1, Angel Sánchez-Lamar2, Regina Montero-Montoya3, Javier Espinosa-Aguirre3, Rafael Camacho-Carranza3
1 Departamento de Farmacología Clínica Experimental, División de Investigaciones, Instituto Nacional de Oncología y Radiobiología. Ave 29 esq. F, CP 10400, La Habana, Cuba.
2 Laboratorio de Genética Toxicológica, Facultad de Biología, Universidad de La Habana. Ave 25, No. 455. e/ J e I, Vedado, La Habana, Cuba.
3 Departamento de Medicina Genómica y Toxicología Ambiental, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, UNAM. Apartado Postal 70228, D.F. 04510, México.
ABSTRACT
Understanding how cells repair DNA damage is a key issue in molecular and cell biology. Ionizing radiation induces DNA damage mainly repaired by a universal mechanism, homologous recombination (HR). Since DNA is chemically the same in all organisms and repair machinery is highly conserved, bacterial models are suitable for assaying genotoxicity, due to their plasticity and low cost. Traditionally, the assays that measure HR use double-strand ends to initiate the recombination events, favoring the RecBCD pathway. The present work was aimed to evaluate gamma rays-induced HR in Salmonella typhimurium, in terms of segregation rates by means of the duplication-segregation assay (seg-dup), which does not favor any particular DNA repair pathway. RecA-independent recombination events were detected at high doses of gamma radiation (150 Gy), which were partially dependent on RecB, SbcCD and RecQ. The seg-dup assay was efficient to elucidate the proteins involved in repairing radiation induced injury. Moreover, due to its simplicity, low costs and the versatility of S. typhimurium strains available, it could be useful to assess the mechanism of action of novel drugs that exert their action by interacting with cell HR machinery.
Keywords: recombination, Salmonella, gamma rays, DNA damage.
RESUMEN
La comprensión sobre cómo las células reparan los daños en el ADN es un aspecto esencial para la biología molecular y celular. Los daños inducidos por la radiación ionizante se reparan a través de un mecanismo universal, conocido como recombinación homóloga (RH). El ADN es químicamente homogéneo entre los organismos y su maquinaria de reparación es altamente conservada, por lo que los modelos bacterianos son apropiados para estudiar la genotoxicidad, dada su mayor plasticidad y bajo costo. Tradicionalmente, los ensayos que miden la RH emplean segmentos terminales de doble cadena para iniciar los eventos recombinatorios, y favorecen la ruta de RecBCD. En este trabajo se estudió RH inducida por radiación gamma en Salmonella typhimurium, según su tasa de segregación evaluada mediante el ensayo de segregación-duplicación (seg-dup). Este ensayo no favorece a ninguna ruta de recombinación. Se detectaron eventos de recombinación independientes de RecA, inducidos por altas dosis de radiación gamma (150 Gy), y parcialmente dependientes de recB SbcCD y RecQ. El ensayo seg-dup dilucidó de forma eficiente cuáles fueron las proteínas involucradas en la reparación del daño inducido por la radiación. Dada la simplicidad y los bajos costos del ensayo, y la versatilidad de las cepas de S. typhimurium generadas, estas pudieran permitir la evaluación del mecanismo de acción de fármacos cuya acción esté mediada por la interacción con la maquinaria celular de RH.
Palabras clave: recombinación homóloga, Salmonella, rayos gamma, daño en el ADN.
INTRODUCTION
It is widely accepted that homologous recombination (HR) is involved in the maintenance of genomic integrity, the generation of genetic diversity, and the proper segregation of chromosomes. The links between recombination and replication have been appreciated for decades and it is now generally accepted that these two fundamental aspects of DNA metabolism are inseparable: In fact, homologous recombination is essential for completion of DNA replication and vice versa [1].
In bacteria, the process of homologous recombination is asymmetric in terms of the genetic contributions made by donor and recipient cells, which contrast the well-studied mechanism of crossing over in eukaryotic sexual reproduction, by which both parental contribute equally [2]. Bacterial HR primary functions in the repair of stalled or collapsed replication forks [3, 4]. At an inactivated replication fork, the HR machinery remodels the forked structure, creating a recombination intermediate recognized by PriA which recruits a replication restart complex; this allows the successive assembly of the primosome and of the replisome so that the fork restarts [5]. It is estimated that around 1 % of cells, in a population growing under normal conditions, experience either a double strand break (DSB) or a single strand break (SSB) due to impediments to replication forks [6, 7]. HR has been traditionally studied by using bacteriophages, bacteria and fungi. In these model organisms, many proteins participating in the process were identified and several pathways proposed to explain their interactions [8].
]]> DNA damage is originated mainly by endogenous sources, like reactive oxygen species (ROS: •OH, •H, •O2- and H2O2) and replication fork arrest, instead of exogenous agents which are a minor and occasional threats [9]. There is increasing interest in radiation-induced DNA damage due to the widespread use of radiation in clinical practice. Exposure to ionizing radiation generates a random deposition of this kind of energy and induces a wide spectrum of damage, by interacting with cell components (lipids, proteins and DNA) directly or mediated by ROS generation [10- 12]. Due to the highly efficient nature of the multiple and overlapping DNA repair mechanisms available to cells, the vast majority of the resultant DNA lesions are accurately repaired and do not lead to mutations [13]. DSBs are a particularly lethal form of DNA damage that prevents chromosome replication if not repaired and, in most bacteria, HR is the dominant pathway for its repair [14], with RecA protein playing a central role. At the beginning of a repair or recombination process, a region of single stranded DNA (ssDNA) is formed at a DSB. This incoming strand, is covered by a RecA protein forming a helical filament and searches the genome for sequence homology by sampling DNA in three dimensions and rapidly binding and unbinding to double-stranded DNA (dsDNA) until homology is found [15,16].Since the 1970’s, the bacterium S. typhimurium has been used as biological model to assay genotoxicity [17] using the Ames’ test. Dr. Ames’ work was critical in linking mutations in DNA to carcinogenesis. Most of the knowledge about recombination was obtained from bacterial assays based on conjugation, transformation and transduction. Zieg y Kushner [18], in 1977, designed an assay in Escherichia coli K-12, with duplication (not in tandem) of two partially deleted Lac Z operons. Since deletions did not overlap, the only way to obtain a Lac+ mutant was by recom-bination, detected by agar tetrazolium lactose platting (white colonies in a red colonies background) [18]. In 1996, Miesel y Roth used an assay base on phage P22 transduction with a small homology sequence (approximately 3-kb long) flanked by markers [19]. Almost a decade after, Fernández-López et al. developed in 2005 an automated assay to measure conjugation by bioluminescence, using the plasmid R388 and an E. coli strain [20]. However, all the assays had in common that they initiated recombination using foreign DNA double strand ends, then favoring the RecBCD pathway, which could have occluded any alternative pathway, if present, mediating on HR. Advantageously, a non-sexual system was designed in S. typhimurium by Galitski and Roth to study recombination, based on phage transduction [21], which does not favor any HR pathway.
Therefore, this work was aimed to evaluate gamma rays induced-homologous recombination in S. typhimurium, in terms of segregation rates, using the duplication-segregation assay (seg-dup) which does not favor any particular HR repair pathway and reconsidering all the previous mechanistic knowledge on HR in this model species. For this purpose, we used a collection of strains, proficient and deficient in enzymatic activities related to different steps and pathways of recombination. A RecA-independent pathway was identified in S. typhimurium, activated in response to radiation injury.
MATERIALS AND METHODS
Strains obtainment
Strains were obtained by phage transduction as first described by Galitski and Roth [21]. The base strain RC1596 (sty(LT2) sulA31 DUP1731 [(leuA1179)*MudA*(nadC220)]), was constructed by using a strain of S. enterica serovar typhimurium LT2 with a duplication (36.2 kb) and ampicillin resistance TT18931 as recipient of transduction from the donor TT19027 strain. Mutations were introduced in seven genes involved in HR in this base strain (Table 1), by using the same protocol, with the RC1596 strain as recipient and “TT” strains as donors.
Reagents and culture conditions
Culture media were purchased from Difco. The chromogenic β-galactosidase substrate X-gal (5-bromo- 4-chloro-3-indolyl-beta-D-galacto-pyranoside) and its solvent N, N-Dimethylformamide were obtained from Sigma.
The complex medium used was nutrient broth (NB), 8 g/L (Difco Laboratories) with NaCl (5 g/L). Solid medium contained agar at 1.5 % (Merck). Phage plates used to score plaque morphologies contained 1 % Bactotryptone (Difco), 0.8 % NaCl and 1.2 % agar or 0.7 % for phage top agar. MacConkey Agar Base (40 g/L, Difco) was supplemented with 1 % lactose. Antibiotics (Sigma Chemical, Co.) were used at the following final concentrations: 30 mg/mL sodium ampicilin (Ap) for single-copy elements (MudA), 20 mg/mL chloramphenicol (Cm), 20 mg/mL tetracycline hydrochloride (Tc), 50 mg/mL kanamycin sulfate (Km). Sterile 0.85 % NaCl (saline) was used to dilute cultures. All incubations were done at 37 °C and liquid cultures were shaken at 150 rpm.
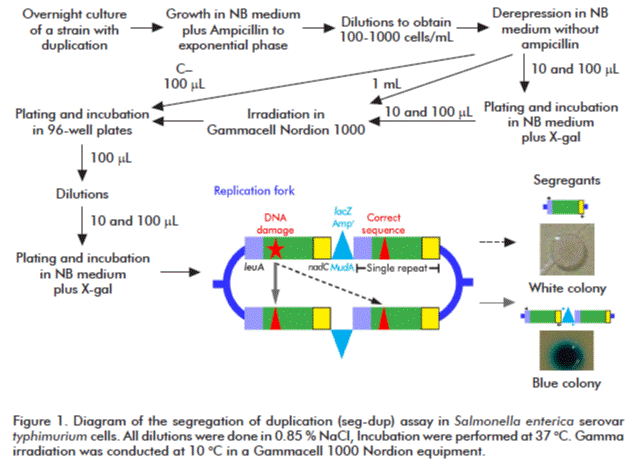
]]> Seg-dup assayThe seg-dup assay was performed exactly as described by Cuétara et al.[22]. Briefly, 200 μL of a bacterial overnight culture was inoculated in 10 mL of NB medium supplemented with the corresponding antibiotics, and it was allowed to grow until exponential phase (O.D. 600 nm = 0.4). It was then diluted 1:10 000 in saline solution followed by a 1:10 dilution in fresh NB medium. This sample was divided in two portions: one to determine spontaneous mutations and the other, to be treated with ionizing radiation to determine induced mutation. Two cultures per treatment (10 and 100 μL each) were kept as controls to guarantee the absence of mutants before radiation treatment.
The experiments were then set-up, with twenty independent cultures per treatment, by placing 100 μL of cells in 96-well plates and incubating them for 16 h. Subsequently, cells were diluted 1:1×106, and 10 and 100 μL of such dilutions were plated in boxes that contained NB plus X-gal (40 μg/mL), and were allowed to grow until colonies with the different phenotypes appeared, either blue or white, the white colonies being the segregants. Segregation rates (μ) were established by calculating the blue/white colonies ratios, by the Luria and Delbrück’s method [23], further extended by Espinosa-Aguirre et al. [24]. The estimator of the number of mutations (m) was calculated by a maximum likelihood method [20], the variance of m established according to the Lea and Coulson method [25]. To set μ, the m obtained was divided by the final number of cells in the culture minus the initial number of cells seeded. The standard deviation of μ was the square root of the m variance divided by the final number of cells in the culture minus the initial number of seeded cells. The 99 % confidence intervals (CI 99 %) were further calculated for each estimate with tree times the standard deviation of μ (μ ± 3sμ); non-overlapping intervals were considered statistically different [23, 25] (Figure 1).
Irradiation with gamma rays
One eppendorf tube containing a milliliter of cells, corresponding to each point of every irradiation doses, were placed in a circular rack that was adjusted to the irradiation chamber. Irradiation was performed using a Gammacell 1000 Nordion equipment (Nordion, Canada). The temperature inside the chamber was set to 10 ºC.
Two dose-response curves (four independent experiments with five replicas each), were conducted per strain (base strain and recA1 deficient) to characterize the response of the base strain, the recA1 deficient strain the expected most sensitive mutant. LD50 was calculated, for both strains, from the curve of better adjustment to the experimental data obtained by using the Origin version 7.0 (OriginLab(TM) NAS Software Partner). From those experiments, it was decided to use 50 and 150 Gy for the strains that exhibited a spontaneous segregation rate similar to that of the base strain and, to avoid lethality, 20 and 50 Gy for the mutants with spontaneous segregation rates an order below the base strain.
RESULTS AND DISCUSSION
Mutation in genes related to HR affects seg-dup rates
Strain recA1 and the double mutation (recB-, recF-) diminished in a significant way the spontaneous segregation rate (μ) of Salmonella (1.9 ± 0.1 × 10-4 and 3.2 ± 0.4 × 10-4, respectively), in comparison with the base strain (7.0 ± 0.4 × 10-3). The effect of the recQ mutation was also significant, diminishing the μ by 7.5 times. Such findings emphasize the important role attributed to RecQ proteins in generating an initiating signal that can recruit RecA for SOS induction and recombination at stalled replication forks, which are required for resumption of DNA replication [26]. The recF mutation significantly reduced in 2/3 the ability of S. typhimurium to recombine (2.6 ± 0.2 × 10-3) while recB mutation significantly increased it (1.5 ± 0.9 × 10-2). Mutations in enzymes of the late steps of recombination distinctively affected the process: the ruvAB mutants exhibited the maximal μ (1 ± 0.1 × 10-2), while ruvC deletion significantly diminished μ (6.9 ± 4.2 × 10-4). The suppressor effect of the sbcCD mutation was previously observed in recBC mutants restoring its recombination ability [27, 28].
]]> In our experiments, the absence of SBCCD notoriously increased the ability of base strain cells for recombination (9.7 ± 0.5 × 10-2). Ivančid-Bade et al., [29] found that some recB mutants exhibited a constitutive expression of the SOS regulon, probably as an adaptive mechanism to cope with endogenous damage, mainly SSBs [29, 30]. They identified the existence of a passive mechanism, independent of RecBCD and RecFOR, to load RecA in a RecB-background, which included RecJ [31]. The mutant strains RecB- spontaneously segregate at a rate of (1.5 ± 0.9 × 10-2). The seg-dup assay supported the direct evaluation of multienzymatic complexes activities, as for RecFOR and RuvABC.HR mutants are grouped in two classes attending to their resistance to radiation
A dose response curve of base strain cells exposed to gamma rays is presented in Figure 2A. As can be observed, cell survival and the number of mutant colonies were clearly affected, and radiation induced a significant number of mutant colonies over the spontaneous value in a increasingly dose-dependent manner. Our data fulfill Boltzmann’s model, which is described by the equation: y = 10 + (1220) / (1 + e ((x -144) / 23)) with a χ2/DoF adjustment of 46.03644, r2 = 0.97216, so LD50 was established at 140 Gy. Since the survival of the base strain cells treated with 150 Gy was approximately 50 %, this dose and the 50 Gy were set as to assay the mutant strains.
The same experiments were conducted with a Rec– strain, but using a different dose range (0-70 Gy) due to its sensitivity to gamma rays (Figure 2 B). RecA-cells evidenced an extreme behavior facing gamma ray irradiation. In an analogous way, we proceeded for the calculation of LD50. In this case, the Boltzmann’s model equation was established as follows: y = – 98 + ((26565)/(1 + e ((x+69) / 25)) with an adjustment of χ2/ DoF = 37.01293, r2 = 0.98532 and LD50 of 16 Gy.
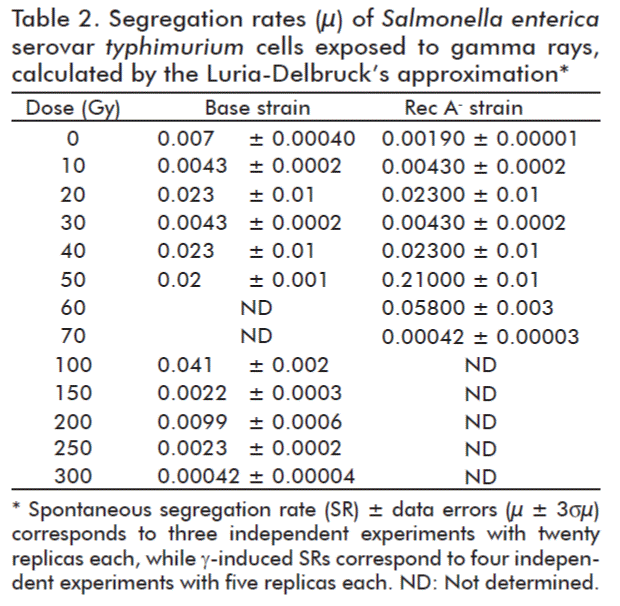
On the other hand, gamma induced segregation rate (μ) was calculated for both strains, by using the Luria-Delbruck’s approximation [23, 25] for each dose (Table 2). Base strain cells were treated with gamma rays, in four conditions with five replicas each irradiation dose (20 samples), and for spontaneous μ in three conditions with 20 replicas each (60 samples).
Our results confirmed the existence of a RecA-independent recombination pathway in Salmonella. RecA-independent recombination was studied by Dutra et al. [32], using different assays, all of them involving double strands ends as initiators of recombination and different mutants defective in RecA and exonucleases. In their experiments, spontaneous recombination rate was evaluated for smaller homology lengths (≤ 0.4 kbp) than ours (36.2 kpb). In a RecA– background, recombination rate was assayed in homology lengths below 0.1 kpb and appeared increased as compared to the base strain, but their results were similar to ours in the gene conversion assay when testing at larger lengths of homology (0.4 kbp), evidencing a recombination rate more than an order of magnitude lower due to RecA deficiency. RecA-independent homologous recombination systems have been regarded as contributing to generate integrating conjugative elements diversity. Such elements are associated with the spread of antibiotic resistance in bacteria and the systems are composed by SSB proteins and an exonuclease [33].
In our experiments it was found that different defects in HR pathways provide cells with variable gamma rays-susceptibility. These results were in agreement with those of Kim and Rose [34], who corroborated that the number of cells in the progeny of strains treated with ionizing radiation correlated with the number of mutant colonies in a dose dependent manner.
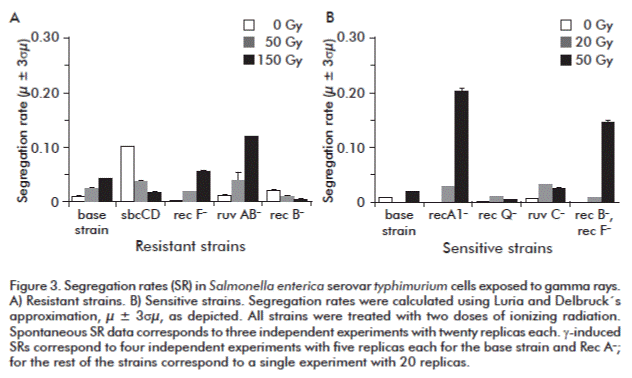
The gamma induced segregation rates of resistant strain (base strain, SbcCD–, RecF–, RecB– and RuvAB–) are shown in Figure 3A. Base strain cells as well as RecF– and RuvAB– cells showed a direct dose-dependent increase in recombination, while SbcCD– and RecB– cells behaved in the opposite way.
Dianov et al., [11] demonstrated that ionizing radiation induced clustered lesions, including abasic sites (AP) and oxidized bases (50 to 80 % of total damage). Theoretically, such lesions can arrive from multiple events or from a single impact [35]. The likelihood of occurrence of clusters increased with radiation density; what could happen in our experiments. These lesions are generally impossible to repair [36].
Gamma rays were very citotoxic to RuvAB- cells. Its population decreased in 75 % at 50 Gy and surviving cells exhibited a high ability to recombine, which increased up to 10-1 at the higher dose. E. coli Ruv– mutants are sensitive to UV light, ionizing radiation and chemical mutagens. This was evidenced by the formation of multinucleated filaments after exposure to low doses of injuring agents [9, 37, 38]. Simple mutants in ruv– were as efficient in recombination as the base strain cells [39, 40]. Our results disagree with those of other groups working with RuvAB– cells, which are less sensitive to UV and gamma rays than the base strain cells [41-43]. Nevertheless, determinations of mutation rates were focused in point reversion of hisG4 (Oc) or isolation of Rifr colony forming units (c.f.u.) by point mutations [41-43].
]]> In fact, our recombination results were mediated by long homology zones, in agreement with Donaldson et al. [44]. They used RuvG– and RuvAB– strains, and observed that the replication fork restoring kinetics in these helicases-deficient strains was similar to the base strain. So, radiation induces replication fork arrest among other mechanisms and the RuvAB activity is non-limiting, with fork restoration releasing recombinogenic substrates for segregation. Moreover, it was suggested by Harris et al. [45], that certain changes in ruvB give the cells with selective advantage.Opposite effects were obtained in RecF– and RecB– cells. RecF– cells increased recombination slightly over the base strain, somewhat expected since RecBCD is recognized as the major pathway to repair DSB, the lethal lesions induced by gamma rays [41]. RecB– cells had the lowest levels of recombination at every dose assayed, remarking the outstanding role of RecBCD pathway in processing DSBs induced by gamma rays. This coincided with reports by Chedin and Kowalczykowski [46], who evidenced that mutations in recB or recC caused a 2-fold decay in conju-gation, these cell types being extremely sensitive to DNA damaging agents [47].
Our findings demonstrate the deficiencies in SbcCD represent “a stimulus” for spontaneous recombination but conversely a disadvantage to respond to radiation induced injury. It has been evidenced that the inactivation of ExoI, the main E. coli exonuclease, increases recombinogenic tails longevity [48]. We obtained a similar effect for SbcCD null mutants, which classified as resistant. It was described that exonuclease activity not only diminished the length of the unwinding tail, but it also restored the SSB proteins exhausted pool, allowing the repetitive joining of free SSB proteins to unwinding DNA [48]. The former could be an argument to explain the coupling of vigorous nuclease activity with the fast and processive helicase activity of the RecBCD holoenzyme [49]. SbcCD alterations block the RecBCD pathway, such alterations probably explaining the decreased μ at high doses of gamma irradiation.
We also analyzed the segregation rate of sensitive strains (RecA–, RuvC–, RecQ– and RecB–/RecF–) after gamma treatment (Figure 3 B). RecQ– and RuvC– cells increase recombination after treatment with 20 Gy and decreased at 50 Gy due to the citotoxicity exerted by radiation in these backgrounds. At this dose, Rec A– and the double mutant seem to trigger highly mutagenic mechanisms to survive. Their segregation rates were an order above that of the base strain. Similar results were obtained with these mutants after treatment with ultraviolet light (10 J/cm2). It is also important to declare that white colonies obtained at this dose were true segregants and did not derive from point mutations in lacZ gene. We also corroborated this in previous studies [22].
It is notorious that in RecQ– cells, segregation rate decreased in one order of magnitude, probably due to the key role of this protein in the recovery of DNA replication. RecQ helicases also prevents excessive recombination by suppressing illegitimate recombination and by initiating HR via the RecF pathway [45]. Moreover, the concomitant action of the RecQ helicase and the RecJ exonuclease at damaged replication forks allows the controlled resection of the lagging strand, resulting in RecA loading by RecFOR and regression of the fork [50]. The regression of the fork allows the DNA lesion to be excised from duplex DNA by the NER, although a significant proportion of recombination events using the RecFOR pathway are RecQ-independent and can be initiated by RecJ nuclease alone or associated with another non identified helicase. Buljubašid et al. [51], demonstrated that RecQ could be partially substituted by UvrD helicase. Rec Q seems to be also important in SOS response. Hishida et al. [52], proposed that prokaryotic and eukaryotic RecQ helicases play a role in coordinating a cell cycle checkpoint response with recombination and replication. Their model predicts that RecQ processes gapped forks into DNA structures that serve to activate RecA and induce the SOS response, which leads to a delay in cell growth, prevention of replication and chromosome segregation, and to the expression of specific DNA repair genes [52].
It was recently demonstrated that radiation induced DNA lesions generate fragmentation and require replication events [53]. RuvABC proteins are also implicated in the resolution of Holliday junctions originating from stalled replication forks. The absence of RuvC could increase the lifetime of species susceptible to be transformed in recombinogenic substrates that might generate the segregation of the duplicated region, explaining the recombination events detected in the present work. Moreover, the lack of a strand sliding apparatus could explain the poor survival and the increased μ. Zahradka et al. [54], demonstrated that RuvABC is necessary for the repair of radiation-induced lesions particularly in recBC–/sbcBC– mutants, this background needed to observe the RecFOR system at work in previous assays [54].
In the case of the RecA– deficient strain and the double mutant RecB–/RecF–, they dramatically decreased c.f.u. counts after gamma irradiation and increased their segregation rate in three orders of magnitude (Figure 3 B). While the ability of the RecA-independent pathway to mediate changes in the bacterial chromosome, Swingle et al. [55], observed low but detectable recombination events between oligonucleotides and chromosomes of E. coli, S. typhimurium, Shigella flexneri and Pseudomonas syringae [55]. It seems that this mechanism is conserved in the gamma subdivision of proteobacteria. Besides, Bierne et al. [56] studied the recombination between consecutive repetitions in E. coli chromosome. They inserted 624-bp long tandem repeats into the lacZ gene and compared the efficiency of deletions of one repetition in RecA–, RecBC–, RecF–, RuvA– and RuvA/RecG– mutants, without detecting any relevant effect. They concluded that deleterious events apparently do not happen via the RecBCD or RecF pathways. They also found a new mutant able to recombine in a RecA-independent manner with a 15-fold increased mutation rate [56]. This hyper-recombinogenicity was associated with an Asp-to-Gly mutation in codon 133, of α subunit of polymerase III encoded by the dnaE gene. Another mutation in this gene, dnaE486, was assayed and shown to enhance RecA-independent recombination. They proposed a slippage mechanism to explain such findings. It is known that RecA-independent exchanges increase in the absence of Rep helicase, the replication delay facilitating the strand slippage proposed [56].
Analyzing the double mutant, there are several mechanisms to be considered in duplication segregation after irradiation. Given the lethality of DSB if they are not efficiently repaired, organisms have developed DNA repair systems such as homologous recombination, single strand annealing, illegitimate recombination and gap fulfilling by translesion synthesis polymerases (TLS) with affected proofreading. Overexpression of TLS by derepression of the DNA damage-inducible LexA regulon caused a 25-fold increase in deletion rate [57].
According to the repair-dependent model of cell radiation survival, extended to include ionizing radiation-induced transformations, the probability of transformation is assumed to scale up with the number of potentially lethal damages that are repaired in a surviving cell (of any kind) or the interactions of such damages. The theory predicts that at high doses, corresponding to low survival, the transformed to surviving cells ratio asymptotically approaches an upper limit. Regarding transformation as equivalent to segregation, since both are forms of homologous recombination that generates a mutant phenotype, then, we could explain the high segregation values obtained for RecA– and the double mutant strain on the basis of the abovementioned theory. Curiously, such a theory was also applied to neoplastic transformation in mammalian cells by Sutherland [58].
We have to remind that HR is a well conserved and regulated process, but when compromised, illegitimate or aberrant recombination may occur between regions of limited or no homology at all [59]. It has been proposed that some microorganisms use it as an adaptive response to long periods of stationary phase, conditioned by adverse environmental stimuli (e.g., nutritional deficiencies or aggressive environments), with aberrant recombination systems probably acquired by horizontal gene transfer in order to repair DNA when their genetic stability was at risk [60]. Besides, in response to radiation injury, SOS induction can act as a backup system for rescuing cell viability [18, 56]; although the repair route of choice varies between species and even among cell lines of the same species [61].
]]>CONCLUSIONS
In summary, our findings suggest that RecA-independent recombinational events could be induced by gamma irradiation. Such events seem to be partially dependent on RecB, SbcCD and RecQ proteins. They also suggested the existence of alternative pathways of recombination to RecFOR and RecBCD in Salmonella.
Since proteins that comprise repair systems seem to be designed according to a Domain Lego principle (by shuffling and recombining a limited repertoire of conserved domains), in the three super-kingdoms of life. The nature of such domains dictates the activities: DNA binding, cleavage, unwinding, ligation, polymerization and molecular adaptors. RecA/Rad recombinases appear to be vertically transmitted during evolution. The same happens with the SMC-ATPases SbcC protein and the Holliday junction resolvase RuvAB [21]. Generally, a bacterial protein has several structural and functional homologs in humans. Considering the above mentioned, we propose the seg-dup assay and the strain collection presented here as a simple, cheap and valuable alternative for studying the mechanisms of action of physical and chemical agents (novel drugs) that interacts with recombination machinery.
ACKNOWLEDGEMENTS
We appreciate the technical assistance of Biol. Sandra Hernández Ojeda and Q.F.B. Carlos Castellanos Barba. This work was partially supported by DGAPA (UNAM) IN204207; IN207513 and CONACyT41469.
CONFLICT OF INTEREST STATEMENT
The authors declare that there are no conflicts of interest.
]]>REFERENCES
1. Syeda AH, Hawkins M, McGlynn P. Recombination and replication. Cold Spring Harb Perspect Biol. 2014;6(11):a016550.
2. Ansari MA, Didelot X. Inference of the properties of the recombination process from whole bacterial genomes. Genetics. 2014;196(1):253-65.
3. Cox MM. Historical overview: searching for replication help in all of the rec places. Proc Natl Acad Sci USA. 2001;98(15):8173-80.
4. Michel B, Grompone G, Flores MJ, Bidnenko V. Multiple pathways process stalled replication forks. Proc Natl Acad Sci USA. 2004;101(35):12783-8.
5. Costes A, Lambert SA. Homologous recombination as a replication fork escort: fork-protection and recovery. Biomolecules. 2012;3(1):39-71.
6. Dillingham MS, Kowalczykowski SC. RecBCD enzyme and the repair of double-stranded DNA breaks. Microbiol Mol Biol Rev. 2008;72(4):642-71.
7. Pennington JM, Rosenberg SM. Spontaneous DNA breakage in single living Escherichia coli cells. Nat Genet. 2007;39(6):797-802.
8. West SC. Molecular views of recombination proteins and their control. Nat Rev Mol Cell Biol. 2003;4(6):435-45.
9. Otsuji N, Horiuchi T, Nakata A, Kawamata J. Strains of Escherichia coli hypersensitive to representative carcinostatic and carcinogenic agents. J Antibiot (Tokyo). 1978;31(8):794-6.
10. Kuipers GK, Poldervaart HA, Slotman BJ, Lafleur MV. The influence of formamidopyrimidine-DNA glycosylase on the spontaneous and gamma-radiation-induced mutation spectrum of the lacZ alpha gene. Mutat Res. 1999;435(2):141-50.
11. Dianov GL, O’Neill P, Goodhead DT. Securing genome stability by orchestrating DNA repair: removal of radiation-induced clustered lesions in DNA. Bioessays. 2001;23(8):745-9.
12. Volkert MR, Elliott NA, Housman DE. Functional genomics reveals a family of eukaryotic oxidation protection genes. Proc Natl Acad Sci USA. 2000;97(26):14530-5.
13. Blackwood JK, Rzechorzek NJ, Bray SM, Maman JD, Pellegrini L, Robinson NP. End-resection at DNA double-strand breaks in the three domains of life. Biochem Soc Trans. 2013;41(1):314-20.
14. Wigley DB. Bacterial DNA repair: recent insights into the mechanism of RecBCD, AddAB and AdnAB. Nat Rev Microbiol. 2013;11(1):9-13.
15. Hiom K. Homologous recombination: how RecA finds the perfect partner. Curr Biol. 2012;22(8):R275-8.
16. Danilowicz C, Peacock-Villada A, Vlassakis J, Facon A, Feinstein E, Kleckner N, et al. The differential extension in dsDNA bound to Rad51 filaments may play important roles in homology recognition and strand exchange. Nucleic Acids Res. 2014;42(1):526-33.
17. Ames BN, Durston WE, Yamasaki E, Lee FD. Carcinogens are mutagens: a simple test system combining liver homogenates for activation and bacteria for detection. Proc Natl Acad Sci USA. 1973;70(8):2281-5.
18. Zieg J, Kushner SR. Analysis of genetic recombination between two partially deleted lactose operons of Escherichia coli K-12. J Bacteriol. 1977;131(1):123-32.
19. Miesel L, Roth JR. Salmonella recD mutations increase recombination in a short sequence transduction assay. J Bacteriol. 1994;176(13):4092-103.
20. Fernandez-Lopez R, Machon C, Longshaw CM, Martin S, Molin S, Zechner EL, et al. Unsaturated fatty acids are inhibitors of bacterial conjugation. Microbiology. 2005;151(Pt 11):3517-26.
21. Galitski T, Roth JR. Pathways for homologous recombination between chromosomal direct repeats in Salmonella typhimurium. Genetics. 1997;146(3):751- 67.
]]>22. Cuétara EB, Sánchez-Lamar A, Hernández-Guadarrama BE, Espinosa-Aguirre JJ, Camacho-Carranza R. Collection of Salmonella typhimurium strains for genotoxicologic and anti-genotoxicologic evaluation. Biotecnol Apl. 2014;31(4):278-84.
23. Luria SE, Delbruck M. Mutations of bacteria from virus sensitivity to virus resistance. Genetics. 1943;28(6):491-511.
24. Espinosa-Aguirre J, Barajas-Lemus C, Hernandez-Ojeda S, Govezensky T, Rubio J, Camacho-Carranza R. RecBCD and RecFOR dependent induction of chromosomal deletions by sodium selenite in Salmonella. Mutat Res. 2009;665(1-2):14-9.
25. Lea DE, Coulson CA. The distribution of the numbers of mutants in bacterial populations. J Genet. 1949;49(3):264-85.
26. Hishida T, Han YW, Shibata T, Kubota Y, Ishino Y, Iwasaki H, et al. Role of the Escherichia coli RecQ DNA helicase in SOS signaling and genome stabilization at stalled replication forks. Genes Dev. 2004;18(15):1886-97.
]]>27. Kushner SR, Nagaishi H, Templin A, Clark AJ. Genetic recombination in Escherichia coli: the role of exonuclease I. Proc Natl Acad Sci USA. 1971;68(4):824-7.
28. Lloyd RG, Buckman C. Identification and genetic analysis of sbcC mutations in commonly used recBC sbcB strains of Escherichia coli K-12. J Bacteriol. 1985;164(2):836-44.
29. Ivancic-Bace I, Vlasic I, Salaj-Smic E, Brcic-Kostic K. Genetic evidence for the requirement of RecA loading activity in SOS induction after UV irradiation in Escherichia coli. J Bacteriol. 2006;188(14):5024-32.
30. McCool JD, Long E, Petrosino JF, Sandler HA, Rosenberg SM, Sandler SJ. Measurement of SOS expression in individual Escherichia coli K-12 cells using fluorescence microscopy. Mol Microbiol. 2004;53(5):1343-57.
31. Jockovich ME, Myers RS. Nuclease activity is essential for RecBCD recombination in Escherichia coli. Mol Microbiol. 2001;41(4):949-62.
]]>32. Dutra BE, Sutera VA Jr., Lovett ST. RecA-independent recombination is efficient but limited by exonucleases. Proc Natl Acad Sci USA. 2007;104(1):216-21.
33. Garriss G, Poulin-Laprade D, Burrus V. DNA-damaging agents induce the RecA-independent homologous recombination functions of integrating conjugative elements of the SXT/R391 family. J Bacteriol. 2013;195(9):1991-2003.
34. Kim JS, Rose AM. The effect of gamma radiation on recombination frequency in Caenorhabditis elegans. Genome. 1987;29(3):457-62.
35. Sutherland BM, Bennett PV, Sidorkina O, Laval J. Clustered damages and total lesions induced in DNA by ionizing radiation: oxidized bases and strand breaks. Biochemistry. 2000;39(27):8026-31.
36. Trampuz A, Piper KE, Steckelberg JM, Patel R. Effect of gamma irradiation on viability and DNA of Staphylococcus epidermidis and Escherichia coli. J Med Microbiol. 2006;55(Pt 9):1271-5.
37. Otsuji N, Iyehara H, Hideshima Y. Isolation and characterization of an Escherichia coli ruv mutant which forms nonseptate filaments after low doses of ultraviolet light irradiation. J Bacteriol. 1974;117(2):337-44.
38. Otsuji N, Iyehara-Ogawa H. Thermoresistant revertants of an Escherichia coli strain carrying tif-1 and ruv mutations: non-suppressibility of ruv by sfi. J Bacteriol. 1979;138(1):1-6.
39. Lloyd RG, Benson FE, Shurvinton CE. Effect of ruv mutations on recombination and DNA repair in Escherichia coli K12. Mol Gen Genet. 1984;194(1-2):303-9.
40. Lloyd RG, Buckman C, Benson FE. Genetic analysis of conjugational recombination in Escherichia coli K12 strains deficient in RecBCD enzyme. J Gen Microbiol. 1987;133(9):2531-8.
41. Sargentini NJ, Smith KC. Involvement of RecB-mediated (but not RecF-mediated) repair of DNA double-strand breaks in the gamma-radiation production of long deletions in Escherichia coli. Mutat Res. 1992;265(1):83-101.
42. Sargentini NJ, Smith KC. Role of ruvAB genes in UV- and gamma-radiation and chemical mutagenesis in Escherichia coli. Mutat Res. 1989;215(1):115-29.
43. Shiba T, Iwasaki H, Nakata A, Shinagawa H. SOS-inducible DNA repair proteins, RuvA and RuvB, of Escherichia coli: functional interactions between RuvA and RuvB for ATP hydrolysis and renaturation of the cruciform structure in supercoiled DNA. Proc Natl Acad Sci USA. 1991;88(19):8445-9.
44. Donaldson JR, Courcelle CT, Courcelle J. RuvAB and RecG are not essential for the recovery of DNA synthesis following UV-induced DNA damage in Escherichia coli. Genetics. 2004;166(4):1631-40.
45. Harris DR, Pollock SV, Wood EA, Goiffon RJ, Klingele AJ, Cabot EL, et al. Directed evolution of ionizing radiation resistance in Escherichia coli. J Bacteriol. 2009;191(16):5240-52.
46. Chedin F, Kowalczykowski SC. A novel family of regulated helicases/nucleases from Gram-positive bacteria: insights into the initiation of DNA recombination. Mol Microbiol. 2002;43(4):823-34.
47. Bibikova M, Carroll D, Segal DJ, Trautman JK, Smith J, Kim YG, et al. Stimulation of homologous recombination through targeted cleavage by chimeric nucleases. Mol Cell Biol. 2001;21(1):289-97.
48. Lehman IR, Nussbaum AL. The deoxyribonucleases of Escherichia coli. V. On the specificity of Exonuclease I (phosphodiesterase). J Biol Chem. 1964;239:2628-36.
49. Ivankovic S, Dermic D. DNA end resection controls the balance between homologous and illegitimate recombination in Escherichia coli. PLoS One. 2012;7(6):e39030.
50. Bennett RJ, Keck JL. Structure and function of RecQ DNA helicases. Crit Rev Biochem Mol Biol. 2004;39(2):79-97.
51. Buljubasic M, Repar J, Zahradka K, Dermic D, Zahradka D. RecF recombination pathway in Escherichia coli cells lacking RecQ, UvrD and HelD helicases. DNA Repair (Amst). 2012;11(4):419-30.
52. Hishida T, Han YW, Shibata T, Kubota Y, Ishino Y, Iwasaki H, et al. Role of the Escherichia coli RecQ DNA helicase in SOS signaling and genome stabilization at stalled replication forks. Genes Dev. 2004;18(15):1886-97.
]]>53. Khan SR, Kuzminov A. Replication forks stalled at ultraviolet lesions are rescued via RecA and RuvABC protein-catalyzed disintegration in Escherichia coli. J Biol Chem. 2012;287(9):6250-65.
54. Zahradka D, Zahradka K, Petranovic M, Dermic D, Brcic-Kostic K. The RuvABC resolvase is indispensable for recombinational repair in sbcB15 mutants of Escherichia coli. J Bacteriol. 2002;184(15):4141-7.
55. Swingle B, Markel E, Costantino N, Bubunenko MG, Cartinhour S, Court DL. Oligonucleotide recombination in Gram-negative bacteria. Mol Microbiol. 2010;75(1):138-48.
56. Bierne H, Vilette D, Ehrlich SD, Michel B. Isolation of a dnaE mutation which enhances RecA-independent homologous recombination in the Escherichia coli chromosome. Mol Microbiol. 1997;24(6):1225-34.
57. Koskiniemi S, Andersson DI. Translesion DNA polymerases are required for spontaneous deletion formation in Salmonella typhimurium. Proc Natl Acad Sci USA. 2009;106(25):10248-53.
]]>58. Sutherland JC. Repair-dependent cell radiation survival and transformation: an integrated theory. Phys Med Biol. 2014; 59(17):5073-90.
59. Ikeda H, Shiraishi K, Ogata Y. Illegitimate recombination mediated by double-strand break and end-joining in Escherichia coli. Adv Biophys. 2004;38:3-20.
60. Bowater R, Doherty AJ. Making ends meet: repairing breaks in bacterial DNA by non-homologous end-joining. PLoS Genet. 2006;2(2):e8.
61. Brena-Valle M, Serment-Guerrero J. SOS induction by gamma-radiation in Escherichia coli strains defective in repair and/or recombination mechanisms. Mutagenesis. 1998;13(6):637-41.
Received in March, 2016. ]]> Acepted in May, 2016.
Rafael Camacho-Carranza. Departamento de Medicina Genómica y Toxicología Ambiental, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, UNAM. Apartado Postal 70228, D.F. 04510, México. E-mail: rcamacho@biomedicas.unam.mx
]]>{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}