Pennisetum purpureum cv. Cuba CT-115 used as biomass bank.Morphophysiological indicators
Pennisetum purpureum vc. Cuba CT-115 utilizado como banco de biomasa. Indicadores morfofisiológicos
Dayleni Fortes,I R.S. Herrera,I M. García,I Ana M. Cruz,I Aida Romero,I
IInstituto de Ciencia Animal. Aptdo. 24 San José de la Lajas, Mayabeque, Cuba.
]]>
ABSTRACT
This research was carried out with the objective of studying the performance of some morphophysiological indicators after grazing P. purpureum cv. Cuba CT-115 used as biomass bank. A completely randomized sampling was used and a total of 15 tufts were collected (experimental unit) immediately after grazing and every 15 days until to complete each growth cycle. The treatments consisted on the age of the grass regrowth from the moment the animals exit the paddock (zero time) and according to the biomass bank technology. During the first year and for each resting cycle the regrowth age influenced (P<0.001) on the plant height. For more of the cycles the length and width of leaves, and the leaf area increased (P<0.001) with the age and the higher values were in the cycles with more favorable climatic conditions for growth and leaf expansion, reaching the leaf area values of 246 cm2 for the fourth cycle of the first year and 204 cm2 for the fifth cycle of the second year. It was also determined the total yield performance with the regrowth age and growth indexes as Culture Growth Rate (CGR) and Net Assimilation Rate (NAR).The results showed that the growth cycles of the studied crop have an specific response pattern in function of the regrowth age and climatic characteristics. The CGR and the NAR have specific response for each cycle and are appropriate indicators for the analysis and characterization of grass growth. It was also proven that the increase of dry matter yield is done with low efficiency (NAR) from the high Culture Growth Rate is reached. It is recommended to incorporate the growth indexes studied in this research as novel tool for the management of other grasses and forage species.
Key words: P. purpureum cv. Cuba CT-115, morphophysiological indicators, biomass bank.
RESUMEN
Este trabajo se realizó con el objetivo de estudiar el comportamiento de algunos indicadores morfofisiológicos después del pastoreo de P. purpureum vc. Cuba CT-115 empleado como banco de biomasa. Se empleó un muestreo completamente aleatorizado y se tomaron 15 macollas (unidad experimental) inmediatamente después del pastoreo y cada 15 días hasta completar cada ciclo de crecimiento Los tratamientos consistieron en la edad de rebrote del pasto a partir del momento de salida de los animales del potrero (tiempo cero) y de acuerdo con la tecnología de banco de biomasa. Durante el primer año y para cada ciclo de reposo la edad de rebrote influyó (P<0.001) en la altura de la planta. Para la mayoría de los ciclos el largo y ancho de las hojas, y el área foliar se incrementaron (P<0.001) con la edad y los mayores valores se presentaron en los ciclos con condiciones climáticas más favorables para el crecimiento y la expansión foliar, alcanzando el área foliar valores de 246 cm2 para el ciclo cuatro del primer año y 204 cm2 para el ciclo cinco del segundo año. También se determinó el comportamiento del rendimiento total con la edad de rebrote y de índices de crecimiento como la Tasa de crecimiento del Cultivo (TCC) y la Tasa de Asimilación Neta (TAN). Los resultados mostraron que los ciclos de crecimiento del cultivo estudiado tienen un patrón de respuesta específico en función de la edad de rebrote y de las características climáticas. La TCC y la TAN tienen respuesta específica para cada ciclo y son indicadores adecuados para el análisis y caracterización del crecimiento del pasto. Además se comprobó que el incremento del rendimiento de materia seca se realiza con una menor eficiencia (TAN) a partir de que se alcanza la máxima Tasa de Crecimiento del Cultivo. Se recomienda incorporar los índices de crecimiento estudiados en este trabajo como herramienta novedosa para el manejo de otras especies de pastos y forrajes.
Palabras clave: P. purpureum vc. Cuba CT-115, indicadores morfofisiológicos, banco de biomasa.
INTRODUCTION
]]> The growth potential and grasses production is determined by the metabolic way used to conduct photosynthesis, as well as its relation with respiration (Herrera 2006).Moreover, it is stated that the biomass accumulation in grasslands is the result of complex interactions between genetic attributes, environmental, physiological process and morphological characteristics that determine the grassland productivity (Pedreira 2006).In this context, it is necessary to know the morphophysiological and structural characteristics of grasses to understand the dynamic of grassland production and their relations within the animal production system in grazing.
That is why, the objective of this research was to study the performance of some morphophysiological indicators after grazing Cuba CT-115 used as biomass bank.
MATERIALS AND METHODS
Location and soil. The studies were conducted in the B dairy unit from the Instituto de Ciencia Animal, San José de las Lajas, Mayabeque. The soil was classified as Brown gray springy soil (Hernández et al. 1999) with slightly acid pH and relatively low contents of N and K.
Treatment and design. The treatments consisted on the regrowth age or grass rest from the moment the animals exit the paddock (zero time) and according to the biomass bank technology (Martínez and Herrera 2006). The duration of the rest cycles or growth is defined according to the biomass bank technology.
Procedure. In a paddock of 0.68 ha a completely randomized sampling was used and a total of 15 tufts were collected (experimental unit) immediately after grazing and every 15 days up to complete each growth cycle according to Fortes et al. (2007).
Measures. The determined indicators were height, total dry matter yield, as well as the percentage of leaves, stems and dead matter according to Herrera (2006).
To determine the area of each leaf the expression used was (Fortes et al. 2007, unpublished data): ]]> AH= 11.41 + 0.89 (±0.09) L*A R2=95.06** SE= ±1.76, where: AH: foliar area, L: leaf length, cm (from the ligule to the apex) and A: leaf width (longitudinal middle point). The Culture Growth Rate (CGR) and the Net Assimilation Rate (NAR) were quantified according to Hunt (1990), starting from the dry mass, foliar area and time (regrowth age after grazing). The age corresponding to the highest CGR was established when making the first derivative from the regression equation equal zero (Ross 2009).
Statistical analysis. Variance analysis was made according to completely randomized design with SPSS version 5.0 (Visauta 2007) and Duncan (1955) test was used for means comparison in necessary cases. Linear models were proved (linear, quadratic and cubic) and non linear (logistic and Gompertz) to study the relation between the regrowth age and yields.
RESULTS AND DISCUSSION
In all growth cycles the tuft height decreased after grazing. During the first and second year and for each rest cycle the regrowth age influenced (P<0.001) on the plant height. In most of the cycles the reduction in the tuft height promoted by grazing, was not compensated by the growth during the grassland rest time. This is due in part, to the different duration of the rest cycles and another to the environmental variations imposed by the climate factors in each growth period. In this study the first three rest cycles which are in the dry season, were the most affected since, the maximum temperatures in these months not exceeded 25 ºC and the number of rainy days was always below five days. Nevertheless, it is important to highlight that in the rest cycles of more duration the plant had great possibilities to reach high height.
For most of the cycles the length and width of leaves, and the leaf area were increased (P<0.001) with the age and the highest values were in the cycles with more favorable climatic conditions for growth and leaf expansion, reaching the leaf area values of 246 cm2 for the fourth cycle of the first year and 204 cm2 for the fifth cycle of the second year. These results are correspond with those found in the same grass by Fernández (2009), who found significant differences between periods, with higher values for the rainy period. Gardner et al. (2000) reported that the increase of the leaf area with the regrowth age becomes constant from the moment it starts flowering and in turn, referred that when plants are efficient tend to invest most of their early growth in expanding its leaf area, resulting better use of solar radiation.
The previously mentioned could contribute to explain the performance of the leaf area in this research, since in most of the cycles, especially in those of more duration, this indicator remained constant to the highest regrowth ages.
The regrowth age influenced (P<0.05) on the performance of the content of leaf, stem and dead material (DM). The stem percentages were always higher than leaves and DM with 83.0 % as higher value in the third cycle of the first year. This pattern was in several rest cycles in which the stem values were high. The leaves content increased with the regrowth age up to 60 days and from there it was stabilized. The DM percentage was also high and decreased, generally, with the regrowth age.
Immediately after grazing (zero time) leaf values did not exceeded 10%, as it is the first fraction the animal intake, whenever sufficient availability exits for it (Gomide et al. 2001).This residual percentage of leaves after grazing has great importance for the plant restoration because it is one the most important elements for the regrowth as photosynthetic source required for the forage growth. From that moment, the leaves percentage with the rest age was increased, in an specific way for each cycle, without exceeding 40%, although the higher increases were recorded up to 60 days, due to, maybe, to the internal positive balance between photosynthesis and respiration achieved by the plant.
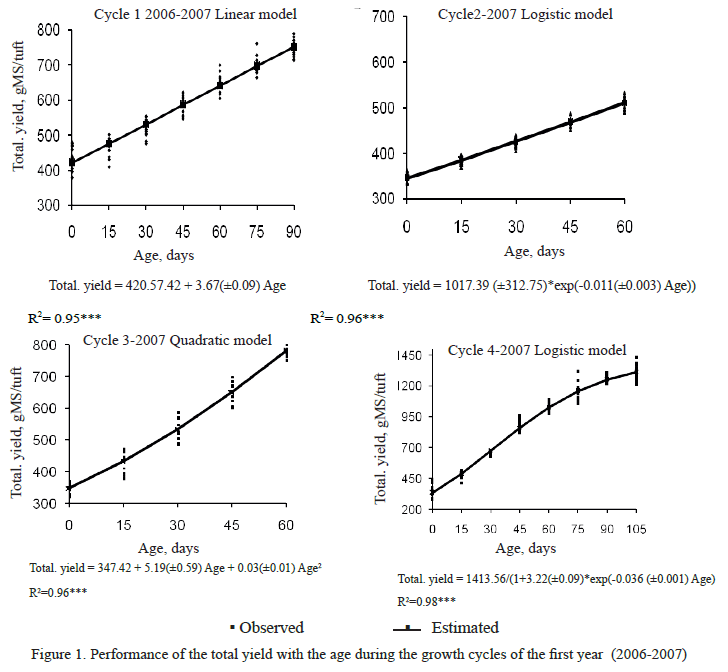
The total biomass production was increased with the regrowth age (figure 1) and in accordance with the established equations, only in the four cycle of the first year, this indicator was stabilized with the regrowth age, which could indicate, that from 90 regrowth days starts the third growth period named senescense or aging period where growth slows caused, mainly, by internal factors that limit it (Pérez and Martínez 1994).
]]> It is interesting to point out that the dry matter yields showed similar tendency with the regrowth age to the tuft height performance, that is to say that both indicators were increased with the age. Probably the most important characteristic that determines the ability of the plant to compete for light is the height, since, little differences in its height can generate effects on the superposition of some leaves in the others and consequently in the dry matter production.The Culture Growth Rate was characterized, in general, for their gradual increase from 15 days of age up to reaching the maxim growth rate, from that moment began to descend, this could indicate that from the referred ages the biomass accumulation is not biologically efficient, so it could be a negative balance between photosynthesis and respiration.
The highest CGR during the first year was reached at 71, 57 and 61 regrowth days for the cycles one, two and four, respectively; the higher grassland productivity was recorded in the rest cycles within the rainy season.
Several studies showed that the forage growth rate could be limited by photo-assimilates supply, by the plant reserves, or by the number, size and activity of the meristem (Perez et al. 2004). All these factors that influence on the CGR are determined, in turn, by climatic factors.
The linear equations obtained for the CGR in the cycles three of the first year, it could attributed to that cycles lasted 60 days and as Cuba CT-115 is a perennial plant of long growth cycle, in this age the plant is under completely conditions for its growth, especially that coincide with the rainy season in which climatic conditions are favorable for its growth, these cycles are characterized by the rapid dry matter accumulation from the first weeks until the 60 days end.
At the beginning of growth the plant leaf area is constituted, mainly, of young leaves with high photosynthetic capacity and high fixation efficiency of atmospheric CO2. As the plant is developing, the leaf senescence processes are intensified, the leaf photosynthetic efficiency is reduced and the plant respiratory loss increased, which compromises the Net Assimilation Rate and the Leaf Area Rate and, consequently, the CGR. This helps to explain the reductions that were in CGR in all the rest cycles from the specific characteristics of each of them.
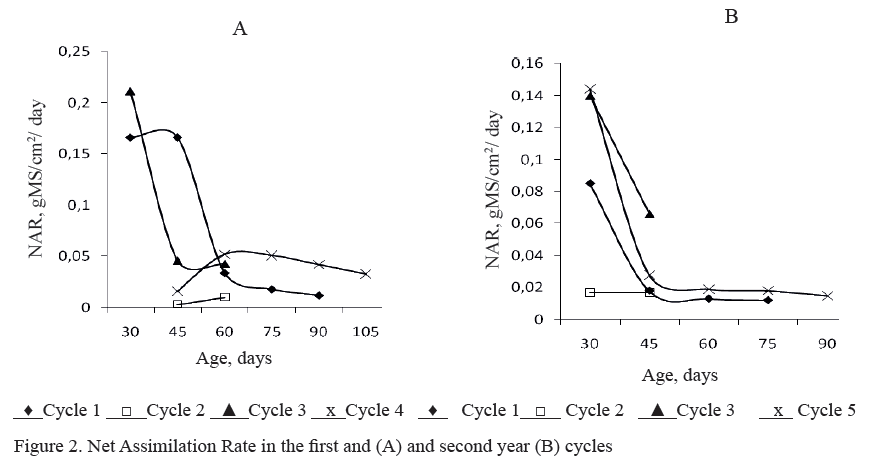
The Net Assimilation Rate (NAR) showed particular performance in each cycle, as well as the pattern that fallowed regarding the regrowth age. The NAR practically did not vary during the first year in cycle one, between 30 and 45 regrowth days but, from this moment it drastically decreased up to 60 days to continue its descend up to 90 days (figure 2 A). The same response pattern was presented in cycle three up to 60 days and it seems that the photo-assimilates production was not enough to replace the plant demands for growth and development. Nevertheless, in cycle two the NAR showed a slight increase which was interrupted by the short growth period. In cycle four, with the rapid growth that occurs between 45 and 60 days it was tried to replace the assimilates needs, that which was not achieved because from 60 days decreased. This performance occurred, maybe, at the expense of the leaf area increase with the regrowth age but with reductions of the Specific Leaf Area and the Leaf Area Duration.
During the second year there was an intense decrease between 45 and 60 days of regrowth during cycles one and five and from that moment on this indicator was stabilized with low values. In cycle three it also decreased while in cycle 2 it was constant (figure 2B). In general, during this year the lowest values compared to the first year were obtained.
In general, the NAR was reduced at the highest regrowth ages in all cycles, probably because the increase of the leaf age, joined to the self-shadowing of the plant inferior leafs reduces the photosynthetic efficiency. In that respect, Andrade et al. (2005) reported that the reduction of NAR with the development is owed to a decrease in the photosynthetic rates more than to the increase of the plant respiratory loss. In this research the CGR was reduced at high regrowth ages, however the plant yields are increased but this is carried out with a lower biological efficiency.
It was proven that the growth cycles of the studied crop have an specific response pattern in function of the regrowth age and climatic characteristics. The CGR and NAR have specific response for each cycle and are appropriate indicators for the analysis and characterization of the grass growth. The increase of the dry matter yield is done with less efficiency (NAR) from the high Culture Growth Rate is reached.
]]> It is recommended to incorporate the growth indexes studied in this research as novel tool for the management of others grasses and forages species, as well as in future researches to deepen in the aspects that integrate elements of morphology and plant physiology and their expressions in the growth and development of grasses and forages dedicated to animal feeding.
REFERENCES
Andrade, A. C., Fonseca, D. M., Lopes, R. S., Nascimento Júnior, D., Cecon, P. R., Queiroz, D. S., Pereira, D. H. & Reis, S. T. 2005. “Análise de crescimento do capim-elefante napier adubado e irrigado”. Ciência e agrotecnologia, 29 (2): 415–423.
Anon, A. 1999. Nueva versión de clasificación genética de los suelos de Cuba. La Habana, Cuba: AGROINFOR, 64 p.
Fernández, A. 2009. Evaluación de Pennisetum purpureum vc. Cuba CT-115 en el Valle del Cauto. M.Sc. Thesis, Universidad de Granma, Cuba.
Fortes, D., Herrera, R. S., Torres, V., García, M., Cruz, A. M., Romero, A., Noda, A. & González, S. 2007. “Determination of a sampling method for the morphophysiological study of grazing Pennisetum purpureum cv. Cuba CT-115”. Cuban Journal of Agricultural Science, 41: 381.
Gardner, F. P., Brent Pearce, R. & Mitchel, R. L. 2000. “Carbon fixation by crop canopies”. In: Physiology of Crop Plants, Iowa State University Press, p. 31.
]]> Gomide, J. A., Wendling, I. J. & Bras, S. P. 2001. “Consumo e produção de leite de vacas mestiças em pastagem de Brachiaria decumbens manejada sob duas ofertas diárias de forragem”. Revista Brasileira de Zootecnia, 30 (4): 1194–1199.Herrera, R. S. 2006. Ecofisiología. Su relación con la producción de pastos. Colombia: Universidad de Nariño, 134 p.
Hunt, R. 1990. Basic growth analysis: plant growth analysis for beginners. London: Unwin Hyman, 112 p.
Martínez, R. O. & Herrera, R. S. 2006. “Empleo del Cuba CT-115 para solucionar el déficit de alimentos durante la seca”. In: Velasco M. E., Hernández A., Perezgrovas R. A. & Sánchez B. (eds.), Producción y manejo de los recursos forrajeros tropicales, México: Univ. Autónoma de Chiapas, p. 75.
Pedreira, B. C. 2006. Interceptação de luz, arquitetura e assimilação de carbono em dosséis de capim xaraés (Brachiaria brizantha (A. Rich.) Stapf.cv. Xaraés) submetidos a estratégias de pastejo rotacionado. M.Sc. Thesis, ESALQ, Piracicaba, 86 p.
Pérez, A. J. A., García, M. E., Enríquez, Q. J. F., Quero, C. A. R., Pérez, P. J. & Hernández, G. A. 2004. “Análisis de crecimiento, área foliar específica y concentración de nitrógeno en hojas de pasto ‘mulato’(Brachiaria híbrido, cv.)”. Téc. Pecu. Méx., 42 (3): 447–458.
Pérez, F. & Martínez, J. B. 1994. Introducción a la Fisiología Vegetal. Madrid: Mundi-Prensa, 217 p.
Ross, D. 2009. “The Derivative”. In: Master Math: Calculus, 2nd ed., p. 344.
Visauta, B. 2007. Análisis estadístico con SPSS 14. Estadística básica. 3rd ed., España: McGrawHill/Interamericana, 358 p.
Received: November 24, 2015
Accepted: December 28, 2015
]]> Dayleni Fortes, Instituto de Ciencia Animal. Aptdo. 24 San José de la Lajas, Mayabeque, Cuba. Email: dfortes@ica.co.cu ]]>
{kind=link}
{kind=link}