










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Dentro de los vertebrados, los reptiles se encuentran entre los grupos taxonómicos más sensibles a disturbios, como la fragmentación y pérdida de hábitat y los efectos locales y globales del cambio climático, especialmente en los grupos insulares (Allison et al., 2002). Los reptiles terrestres cubanos están representados por 160 especies de ellas 64 pertenecen al género Anolis (Losos 2009 y Rodríguez Schettino et al., 2013).
Los reptiles ocupan una gran variedad de hábitats y de tipos de coberturas, desde áreas abiertas y bosques transformados, hasta bosques conservados (Duellman 1979); este uso diferencial de los hábitats o de las coberturas originales da lugar a patrones de distribución horizontal y vertical diferente de los ensamblajes, con la consiguiente diferenciación en el uso de los recursos, entre estos el alimentario (Pianka 1989; Duellman 1979; Vitt y Pianka 2005; Mesquita et al., 2006).
La composición de la fauna de lagartos está profundamente influenciada por la estructura de la vegetación. En general los requerimientos de hábitat de casi todas las especies de reptiles de Las Antillas se encuentran definidos por adaptaciones morfológicas, fisiológicas y conductuales (Schwartz y Henderson 1991).
Cuando ocurren cambios en el paisaje, por acción antropogénica como la fragmentación y pérdida de coberturas vegetales, se afecta la disponibilidad, abundancia y riqueza del recurso disponible (Carvajal y Urbina-Cardona 2008). Esta situación repercute en la respuesta y amoldamiento de las especies a las nuevas condiciones de uso de los recursos que queden disponibles.
Partiendo de estas bases teóricas, esta investigación va dirigida a evaluar la incidencia de las modificaciones del bosque semideciduo, en la dinámica de recuperación posterior a las intervenciones del aprovechamiento forestal, sobre el uso de ciertos recursos, en las comunidades de lagartos y su relación con las variables de vegetación.
MATERIALES Y MÉTODOS
La investigación se llevó a cabo en el bosque semideciduo en dos sectores del Parque Nacional Guanahacabibes: Cabo San Antonio y Cabo Corrientes, que se ubica en el extremo más occidental de la isla de Cuba (Figura 1).

Fig. 1 - Esquema de ubicación de las localidades de estudio, Cabo de San Antonio y Cabo Corrientes en la Península de Guanahacabibes
Para evaluar los impactos que provocan las modificaciones del bosque aprovechado desde el punto de vista forestal sobre las comunidades de lagartos. Se evaluaron en cada sector dos tratamientos con diferentes tiempos de recuperación post aprovechamiento forestal durante tres años (2013, 2014 y 2015):
Cabo de San Antonio (CSA) y Cabo Corrientes (CC).
Tratamiento 1-5 años de recuperación post aprovechamiento forestal.
Tratamiento 2 e 30 años o más de recuperación post aprovechamiento forestal.
En cada tratamiento se realizaron 10 transectos con una longitud de 100 m y 5 m de ancho, a una distancia de 100 m uno de otro siguiendo el método de Rand (1964). Se recorrieron a paso lento en los horarios de 9:00 a 13:00 horas en dos periodos del año lluvioso y poco lluvioso y se tomaron los siguientes datos:
Sexo y estadio (Hembras y machos adultos o juveniles).
Tipo de sustrato donde se encuentra el individuo al ser observado, tronco de los árboles o arbustos, roca, rama o suelo.
Cuando el sustrato es rama o tronco se mide la altura del suelo con cinta métrica graduada en 0,5 cm y el diámetro de la parte que lo sostiene.
Iluminación a la que se expone el animal, clasificándola en sol filtrado (F), sol directo (D) y sombra (S).
Análisis de datos
Se realizó un estudio de la fortaleza de la relación entre las variables potencialmente predictoras y la variable dependiente, utilizándose el coeficiente de correlación de rangos de Spearman. Entre múltiples modelos probados se decidió seleccionar el modelo que incluye como variables intrasujeto, localidad, estación, tratamiento y año.
Para las asociaciones entre las variables (altura de percha, iluminación y sustrato frecuentado) se realizó una regresión logística binaria.
Se aplicó el estadístico de Mantel-Haenszel, en el análisis multivariado para estimar la ventaja común controlando por los estratos de altura, iluminación y sustrato. Se determinó, además el estadístico Chi-Cuadrado (c2) para verificar mediante las pruebas de Breslow-Day y De Tarone la significación de la homogeneidad de razón de ventajas entre los estratos controlados.
Mediante el análisis de correspondencia canónica, empleando el programa PcOrd versión 4.10, se analizan de un modo directo las asociaciones que se establecen entre las densidades de los lagartos y las variables medidas de la estructura del bosque.
Se realiza la prueba de Mantel que es muy empleada en el análisis de datos ecológicos multivariados. Es una regresión en la cual las matrices de las variables son de distancia o de similitud, que resume las similitudes en muchas variables entre pares de localidades. Este estadístico, sin embargo, no tiene una distribución teórica fija, sino que se compara con la distribución nula que se obtiene por las permutaciones de filas y columnas de la matriz de datos (modelo nulo).
RESULTADOS Y DISCUSIÓN
Efecto de la recuperación post aprovechamiento forestal sobre el uso de los recursos
Altura de percha
La Tabla 1 recoge las asociaciones entre las variables categóricas ponderadas por las frecuencias de las variables que se declaran en la primera columna. Se presentan las variables con las que se calcularon las asociaciones con la variable dependiente, mediante regresión logística binaria. La razón de ventajas y su intervalo de confianza se presentan también y en la última columna de la tabla se señalan las asociaciones que resultaron significativas, así como la significación bilateral.
Tabla 1 - Grado de asociación entre variables categóricas. Total de lagartos
| Variable de ponderación | Asociaciones | Razón de ventajas | Límite inferior | Límite Superior | P |
| Frecuencia por Altura | Localización CC/CSA | ||||
| Alt_1.99/Alt_0.99 | 0,836 | 0,714 | 0,979 | 0,026* | |
| Alt_2.00/ Alt_0.99 | 0,714 | 0,979 | 0,714 | 0,027* | |
| Año 2007/2006 | 0,770 | 0,645 | 0,919 | 0,004* | |
| Año 2008/2006 | 0,764 | 0,643 | 0,908 | 0,002* | |
| tratamientos | |||||
| Alt_1.99/Alt_0.99 | 1,155 | 0,987 | 1,352 | ns | |
| Alt_2.00/ Alt_0.99 | 1,101 | 0,848 | 1,431 | ns | |
| Año 2007/2006 | 0,946 | 0,792 | 1,129 | ns | |
| Año 2008/2006 | 1,024 | 0,862 | 1,218 | ns | |
| Estación | |||||
| Alt_1.99/Alt_0.99 | 1,003 | 0,857 | 1,173 | ns | |
| Alt_2.00/ Alt_0.99 | 0,724 | 0,555 | 0,943 | 0,017* | |
| Año 2007/2006 | 0,770 | 0,645 | 0,919 | 0,004* | |
| Año 2008/2006 | 0,764 | 0,643 | 0,908 | 0,002* | |
| Localización*trat. | 1,118 | 0,971 | 1,286 | ns | |
| Trat.*Estación | 0,934 | 0,812 | 1,075 | ns | |
| Trat. *Años (2007) | 0,946 | 0,792 | 1,129 | ns | |
| Trat. *Años (2008) | 1,024 | 0,862 | 1,218 | ns | |
| Estación*Años (2007) | 0,953 | 0,798 | 1,137 | ns | |
| Estación*Años (2008) | 0,956 | 0,804 | 1,136 | ns | |
| Frecuencia por Iluminación | Localización CC/CSA | ||||
| Filtrado/sol* | 1,447 | 1,222 | 1,715 | <0,001* | |
| Sombra/sol | 1,080 | 0,894 | 1,304 | ns | |
| Año 2007/2006 | 0,735 | 0,616 | 0,878 | 0,001* | |
| Año 2008/2006 | 0,742 | 0,624 | 0,883 | 0,001* | |
| tratamiento | |||||
| Filtrado/sol | 0,890 | 0,752 | 1,053 | ns | |
| Sombra/sol | 1,026 | 0,850 | 1,237 | ns | |
| Año 2007/2006 | 0,735 | 0,616 | 0,878 | ns | |
| Año 2008/2006 | 0,742 | 0,624 | 0,883 | ns | |
| Estación | |||||
| Filtrado/sol* | 1,203 | 1,016 | 1,424 | 0,032* | |
| Sombra/sol* | 1,232 | 1,021 | 1,487 | 0,029* | |
| Año 2007/2006 | 0,963 | 0,807 | 1,149 | ns | |
| Año 2008/2006 | 0,982 | 0,826 | 1,167 | ns | |
| Localización*trat. | 0,902* | 0,783 | 1,038 | ns | |
| Trat.*Estación | 1,057 | 0,919 | 1,216 | ns | |
| Trat. *Años (2007) | 0,926 | 0,776 | 1,105 | ns | |
| Trat. *Años (2008) | 1,005 | 0,846 | 1,195 | ns | |
| Estación*Años (2007) | 0,963 | 0,807 | 1,149 | ns | |
| Estación*Años (2008) | 0,982 | 0,826 | 1,167 | ns | |
| Frecuencia por sustrato | Localización CC/CSA | ||||
| Roca/suelo | 1,193 | 0,930 | 1,530 | ns | |
| Tronco/suelo | 0,863 | 0,711 | 1,047 | ns | |
| Rama/suelo | 0,000 | 0,000 | - | ns | |
| Año 2007/2006 | 0,898 | 0,747 | 1,078 | ns | |
| Año 2008/2006 | 0,866 | 0,725 | 1,035 | ns | |
| tratamiento | |||||
| Roca/suelo | 1,036 | 0,808 | 1,327 | ns | |
| Tronco/suelo | 1,070 | 0,882 | 1,299 | ns | |
| Rama/suelo | 6,982 | 2,689 | 18,129 | <,001* | |
| Año 2007/2006 | 0,873 | 0,727 | 1,049 | ns | |
| Año 2008/2006 | 1,045 | 0,874 | 1,249 | ns | |
| Estación | |||||
| Roca/suelo | 0,769 | 0,599 | 0,986 | 0,038* | |
| Tronco/suelo | 0,775 | 0,639 | 0,941 | 0,010* | |
| Rama/suelo | 0,469 | 0,238 | 0,922 | 0,028* | |
| Año 2007/2006 | 1,036 | 0,863 | 1,245 | ns | |
| Año 2008/2006 | 0,963 | 0,806 | 1,152 | ns | |
| Localización* trat. | 0,910 | 0,786 | 1,053 | ns | |
| Trat. *Estación | 1,123 | 0,970 | 1,300 | ns | |
| Trat. *Años (2007) | 0,873 | 0,727 | 1,049 | ns | |
| Trat. *Años (2008) | 1,045 | 0,874 | 1,249 | ns | |
| Estación*Años (2007) | 1,036 | 0,863 | 1,245 | ns | |
| Estación*Años (2008) | 0,963 | 0,806 | 1,152 | ns |
Fuente: Elaboración propia a partir de las tablas de salida de SPSS.
El análisis multivariado (Tabla 2) mostró diferencias significativas en las combinaciones "Localidad* tratamiento" controlado por los estratos de altura con una razón de ventajas común de 0,9 y una significación en la prueba de homogeneidad de ventajas de 0,018 lo que demuestra que el comportamiento no es el mismo en los diferentes estratos. Otro tanto sucede con "Localización *estación" con razón de ventajas de 1,085 que es la cantidad de veces que es más probable de encontrar los lagartos en una estación que en otra cuando se controla por la altura y una significación en la prueba de homogeneidad de ventajas de 0,033.
Tabla 2 Resultados de asociación en el análisis multivariado
| Asociaciones | Estadístico de Mantel-Haenszel | Significación de la homgeneidad de razón de ventajas | ||||
| Razón de ventajas común | Significación asintótica bilateral | Estadístico(2 | Significación asintótica bilateral | |||
| Frecuencia por altura | Localización*Tratamiento*Estación | 1,094 | 0,213 | 0,043 | 0,836 | |
| Localización*Tratamiento*Años | 0,894 | 0,119 | 1,964 | 0,375 | ||
| Localización*Estación*Año | 1,089 | 0,237 | 0,514 | 0,773 | ||
| Estación* Tratamiento*Años | 1,070 | 0,363 | 1,316 | 0,518 | ||
| Localización*Tratamiento*Altura | 0,900 | 0,143 | 8,013 | 0,018* | ||
| Localización*Estación*Altura | 1,085 | 0,254 | 6,80 | 0,033* | ||
| Estación* Tratamiento*Altura | 1,072 | 0,334 | 0,274 | 0,872 | ||
| Frecuencia por Iluminación | Localización*Tratamiento*Estación | 0,900 | 0,144 | 0,221 | 0,638 | |
| Localización*Tratamiento*Años | 0,899 | 0,139 | 2,083 | 0,149 | ||
| Localización*Estación*Años | 1,087 | 0,248 | 1,251 | 0,263 | ||
| Estación* Tratamiento*Años | 1,057 | 0,443 | 1,662 | 0,436 | ||
| Localización*Tratamiento*Iluminación | 0,910 | 0,192 | 9,837 | 0,007* | ||
| Localización*Estación*Iluminación | 1,079 | 0,291 | 1,345 | 0,510 | ||
| Estación* Tratamiento*Iluminación | 1,059 | 0,422 | 0,079 | 0,961 | ||
| Frecuencia por sustrato | Localización*Tratamiento*Estación | 0,908 | 0,196 | 0,390 | 0,532 | |
| Localización*Tratamiento*Años | 0,910 | 0,206 | 4,040 | 0,133 | ||
| Localización*Estación*Año | 1,075 | 0,335 | 2,917 | 0,233 | ||
| Estación* Tratamiento*Año | 1,126 | ,114 | 1,066 | 0,114 | ||
| Localización*Tratamiento*Sustrato | 0,947 | 0,467 | 0,206 | 0,902 | ||
| Localización*Estación* Sustrato | 1,058 | 0,453 | 21,670 | 0,001* | ||
| Estación* Tratamiento* Sustrato | 1,139 | 0,084 | 2,890 | 0,409 | ||
*diferencias significativas al 5 % >Fuente: Elaboración propia a partir de las tablas de salida de SPSS.
Altura
Ponderación: frecuencia por alturas
La categoría de altura de mayor uso en todas las combinaciones de los tres efectos considerados (año, estación y tratamientos) para ambas localidades fue de 0-0,9 m (Figura 2). Con la localidad resultaron significativos las asociaciones en las categorías de altura y años, en el primer caso la cantidad de veces que es probable encontrar los lagartos a la altura de 1,99 respecto a 0,99 m es de 0,836. No indicaron asociaciones significativas entre los tratamientos para ninguna de las combinaciones año*estación. La estación mostró asociación significativa con la frecuencia de lagartos en alturas iguales o superiores a 2,00 m y con los años. El resto de las asociaciones cuando se pondera por la frecuencia según la altura no resultaron significativas.
En ambas localidades, hubo una tendencia a la disminución del uso de la categoría 0-0,9 m. Para el año 2013, a partir del período lluvioso comenzó a disminuir levemente el uso de esta categoría establecida en el tratamiento de cinco años de recuperación, lo que continuó de forma más marcada en el 2014. Esto estuvo influenciado por el aumento de mayor frecuencia de lagartos por el uso de la altura intermedia (1,00-1,99 m). Este mismo patrón comenzó en el año 2014 en el período lluvioso para el tratamiento de más de 30 años sin aprovechamiento forestal. Para el año 2015, en ambas estaciones, las diferencias entre usos de la altura 0-0,9 m fueron menores que en años anteriores.
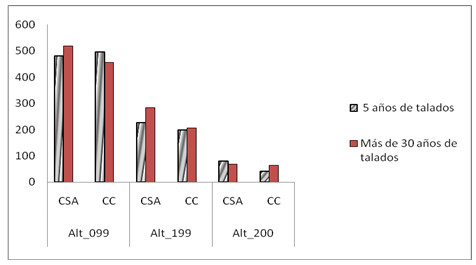
Fig. 2 - Frecuencia de lagartos. Localización Cabo de San Antonio (CSA) y Cabo Corrientes (CC) por tratamiento por las tres categorías establecidas de altura de percha
En ambas localidades la altura preferida por los lagartos fue de 0-0,9 m, con una tendencia a la disminución del uso de esta categoría en los dos tratamientos para los dos últimos años, aumentando la cantidad de individuos en la segunda categoría establecida (1,00-1,9 m). En la localidad de Cabo Corrientes, las diferencias significativas aparecen a partir del 2013 en la época de lluvia y continúan durante todo el 2014 en ambas estaciones y en la localidad de Cabo de San Antonio en la época de sequía en el último año de muestreo. Esta respuesta se debe a que los lagartos se observan ocupando individuos propios de la regeneración natural del bosque, menor de 2 m de altura y menor de 2 cm de diámetro, establecida por Delgado y Ferro (2000), que coincide con la preferida por los lagartos de la comunidad en estudio.
Esta regeneración natural varía en dependencia del tratamiento que se le hace al bosque, en lugares más perturbados hay una variación más significativa en cuanto a la densidad en el período lluvioso y el período poco lluvioso, en los tratamientos de menos tiempo de aprovechamiento forestal se define por el banco de semillas y en los tratamientos más conservadas la estrategia de la regeneración es por plántulas.
En los casos que las perturbaciones tanto antrópicas como naturales provocan claros en el bosque por la disminución de la cobertura vegetal como en el caso del tratamiento de menos tiempo de recuperación postaprovechamiento forestal, el comportamiento de la regeneración es totalmente diferente. Los manejos forestales provocan disminución en la diversidad de especies forestales en el bosque y no activan el incremento de ellas en la regeneración natural (Delgado y Ferro, 2000).
Desde el punto de vista forestal, a mayor altura de los estratos es notable un incremento de la copa de los árboles, trayendo aparejado una mayor densidad del dosel y con ello una disminución de la penetración de la luz solar (Delgado y Ferro, 2006). Por tratarse de especies poiquilotérmicas que precisan de la incidencia de la luz solar para regular su temperatura interna, los lagartos frecuentan alturas menores en busca de los claros que pueden aparecer debido al aprovechamiento forestal.
Iluminación
Ponderación: frecuencia por iluminación
El mayor uso de este recurso lo tuvo la categoría establecida como sol filtrado por la vegetación (Figura 3), cuya frecuencia fue estable a lo largo de todos los años de estudio para los dos tratamientos. En el tratamiento de cinco años postaprovechamiento forestal, los valores de uso del sol filtrado tuvieron mayor variabilidad.
Con el efecto de la localidad resultaron significativas las categorías de Iluminación de sol filtrado por la vegetación y sol y el año. En cuanto a los tratamientos no se mostraron asociaciones significativas, la estación indicó asociaciones significativas en cuanto a las tres categorías establecidas de iluminación, no así con los años. Las demás asociaciones en el análisis univariado no resultaron significativas para un 5 %. En el análisis multivariado, solo resultó significativa la combinación "Localización*tratamiento" cuando se controla por los estratos de iluminación con una razón de ventajas de 0,910 y una significación de homogeneidad de razón de ventajas de 0,007.
En ambas localidades y para el año intermedio hubo una disminución apreciable del uso de la categoría soleado y un aumento de la categoría sombra. En el Cabo de San Antonio no se observó un patrón definido de estos cambios entre los tratamientos, pero sí en Cabo Corrientes, donde el uso de la categoría soleado fue mayor en el tratamiento 1, mientras que la categoría sombra fue mayor en el tratamiento 2.
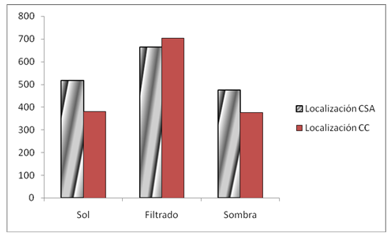
Fig. 3 - Frecuencia de lagartos. Localización Cabo de San Antonio (CSA) y Cabo Corrientes (CC) por las tres categorías establecidas de Iluminación
El comportamiento del recurso iluminación fue estable en todos los años de estudio, prefiriendo siempre sitios con sol filtrado. Como se ha comentado en previos análisis, tratamos un grupo vertebrado que no regula su temperatura interna y períodos prolongados de exposición al sol directo, así como de sombra total, pueden afectar el funcionamiento propio del organismo y la realización de sus funciones vitales (Rodríguez Shettino 1999).
El costo de la termorregulación depende, en gran parte, del hábitat: aumentar la temperatura corporal es más difícil en bosques cerrados que en áreas más abiertas, por lo que las especies de ambientes costosos para la termorregulación son, generalmente, pasivas ante los cambios de la temperatura ambiental, es decir, son termoconformes (Huey y Slatkin 1976).
En ambas localidades, en el 2013, hubo una disminución notable del uso de la categoría sol directo y un aumento de la categoría sombra, las diferencias no arrojan significación, pero asumimos que las condiciones climáticas de este año se mostraron diferentes al resto de los años , el período lluvioso fue por encima de la norma histórica, como nos referimos anteriormente y el período poco lluvioso fue más corto por lo que el período de caducidad de los árboles disminuyó; este tipo de bosque se identifica por tener entre el 40 y 70 % de los árboles del estrato arbóreo superior, pierden las hojas en el periodo poco lluvioso (Del Risco 1999) y al ser este período más corto disminuye este efecto, lo que conllevan a que estos cambios hayan influido de manera diferencial en el uso de los recursos aquí discutidos por la comunidad de lagartos.
En Cabo Corrientes, el uso de la categoría sol directo fue mayor en el tratamiento 1. La estructura de la vegetación condiciona la temperatura ambiente de cada tipo de hábitat, pero en ella subyacen también diferentes posibilidades de ocupar verticalmente el espacio, el tratamiento de cinco años de recuperación post aprovechamiento forestal es la de menor cobertura vegetal y consecuentemente de mayor incidencia solar, mientras que las de más de 30 años sin intervenir tienen mayor incidencia de sombra.
Sustrato
Ponderación: frecuencia por sustrato
En el análisis de uso de sustratos, en ambas localidades, el más utilizado para todas las combinaciones (años, tratamientos y estaciones) fueron los troncos de los árboles (Figura 4) con tendencia al aumento de su uso en el tratamiento de más de 30 años post aprovechamiento forestal.
En este caso, resultó significativa la asociación tratamiento*sustrato en la categoría rama/suelo la cantidad de veces que es más probable de encontrar lagartos en las ramas que en el suelo es de 6,98. La asociación estación*sustrato mostró una diferencia significativa en el uso de los sustratos. El análisis multivariado dio un valor de Chi cuadrado muy alto que indica falta de homogeneidad en la razón de ventajas entre los sustratos en la combinación "Localización* Estación.
A lo largo de los tres años de estudio, no aparecieron cambios estadísticamente significativos para el uso del recurso tronco, pero si una tendencia al aumento de su uso en el tratamiento de más de 30 años post aprovechamiento forestal, con un rango de valores entre 40 y 70 %. El uso de este recurso se incrementó en los dos últimos años del estudio por igual en ambos tratamientos.

Fig. 4 - Frecuencia de lagartos por localidad Cabo de San Antonio (CSA) y Cabo Corrientes (CC) por las cuatro categorias establecidas de sustrato
En todas las localidades y períodos, se observó una preferencia por los troncos como sustrato. Resultados similares se han encontrado por Rodríguez Schettino (1999), coincidiendo en la selección del grupo. La especie más abundante en la comunidad son especies simpátricas del ecomorfo tronco-suelo.
El aprovechamiento forestal, se caracteriza por la extracción de individuos con diferentes fines, y al extraer volúmenes altos de madera en una formación vegetal, se modifica la estructura de la misma y trae aparejado cambios como incremento de temperatura, disminución de la humedad, cambios de luz y de velocidad del viento (Pinard y Putz, 1996), pero en los años consecutivos la respuesta natural del bosque lleva a una recolonización de estos espacios por especies invasoras, oportunistas y austeras (Delgado et al., 2005) que incrementa la densidad vegetal, y que con el paso del tiempo, por la competencia entre todos los nuevos individuos y los ya restantes, se llega a la eliminación de los menos adaptados, y por ende a la disminución del número de individuos por hectárea.
También se debe considerar que los bosques más conservados tienen entre sus características una mayor rugosidad de los troncos por las especies que lo caracterizan (Ferro, 2004), y esto favorece la presencia de gran variedad de microhábitats para los invertebrados y su acumulación, lo que sirve de alimento al grupo en análisis.
Las particularidades aparecen en la localidad Cabo de San Antonio donde aumentó el uso de las ramas en el período lluvioso, este período fue el más lluvioso de los tres años muestreados con valores por encima de la media histórica (1480.mm) y aumenta la humedad relativa, desplazándose los individuos a mayores alturas.
La explicación a estos resultados es que la supervivencia y reproducción de los lagartos está influenciada por el uso del suelo, de perchas o nicho estructural y la disponibilidad de alimentos o nicho trófico (Shoener, 1968), son elementos que pueden ser alterados a tal modo por el manejo forestal, que favorezca a algunas especies en detrimentos de otras.
Al realizar una correlación entre las densidades de los estratos de la vegetación y la preferencia de las cuatro categorías de sustratos establecidas para este estudio, obtenemos resultados significativos negativos entre las preferencias de las rocas y las ramas con el estrato arbustivo. Podemos asumir que esto tiene lugar debido a que cuando disminuye la rocosidad aumenta la densidad del estrato arbustivo y al ser la densidad de este estrato mayor la ramificación es menor, ya que esta aumenta en los lugares menos densos.
Relación con las variables de vegetación. Estudio de la fortaleza de la relación entre las variables potencialmente predictoras y la variable dependiente
La frecuencia total de lagartos se correlaciona significativamente con la riqueza del estrato arbustivo (-0,294), altura del estrato arbóreo superior (0,208), longitud media de los troncos caídos (0,182) y la regeneración natural (-0,261). La frecuencia de machos sólo mostró relación significativa con la riqueza del estrato arbustivo (-0,206) y la regeneración natural (-0,189) al igual que los adultos (-0,230 y -0, 189 respectivamente) la frecuencia de hembras y de juveniles no mostraron relación significativa con ninguna de las variables de vegetación.
En las Figuras 5 y 6, se presentan los resultados de los análisis de correspondencias canónicas, llevados a cabo entre las variables de vegetación de cada tratamiento de estudio y las frecuencias de especies de lagartos en cada una de ellas. Se comprueba que el tiempo de recuperación de los bosques influye en las diferencias entre las asociaciones de los lagartos con estas variables. Anolis quadriocellifer es la especie que no se afecta con las variables de vegetación medidas en los dos tratamientos de estudios en el sector Cabo de San Antonio.
En análisis del período lluvioso, el primer cuadrante de la Figura 5, muestra las especies que preferentemente utilizan el suelo y las rocas como sustrato, Anolis loysiana y Leiocephalus macropus marcan asociación con el Área basal de la formación vegetal, pues su densidad fue máxima en las parcelas que más elevada cifra presentaron.
Leiocephalus carinatus frecuentó sitio de mayores alturas del estrato arbustivo, mientras que Ameiva auberi fue más detectada en áreas de altos volúmenes. En el segundo cuadrante solamente observamos dos especies de lagartos, Anolis porcatus y A. homolechis asociadas ellas a la riqueza de los árboles y a los sitios de mayores densidades respectivamente. Por otra parte, Anolis angusticeps y A. alutaceu no mostraron asociación con ninguna de las variables de vegetación medidas. La primera especie siempre fue más detectada en espacios abiertos con penetración del sol y en tratamientos de menos tiempo de aprovechamiento forestal, se encontró en el sustrato rama y hoja en igual proporción, debido a que los movimientos de este pequeño lagarto son lentos y tiene el hábito de mantenerse muy unido a las ramas (Böhm et al., 2013) sin embargo de Anolis alutaceu se observaron pocos individuos, mayormente en el suelo y a la sombra coincidiendo con lo planteado por Schwartz y Henderson (1991).
En el período poco lluvioso, el comportamiento de las especies difiere de la anterior, excepto en caso del endémico regional que es la única especie que ocupa los cuatro tipos de sustrato en ambas áreas con frecuencia alta por el tronco de los árboles del 70 % a l80 % (Pérez et al., 2014). En el primer cuadrante encontramos las variables de vegetación volumen y área basal en asociación con Anolis homolechis, A. porcatus, A. luteogularis, Leiocepahaulus macropus, L. carinatus y Ameiva auberi, se registraron en parcelas con mayores riquezas específicas de vegetación, donde se evidencia una estrategia de segregación para disminuir la competencia por los recursos, coincidiendo estos resultados con los de González (2017), mientras que Anolis alutaceus y Anolis loysiana no mostraron asociación alguna con ninguna variable medida.
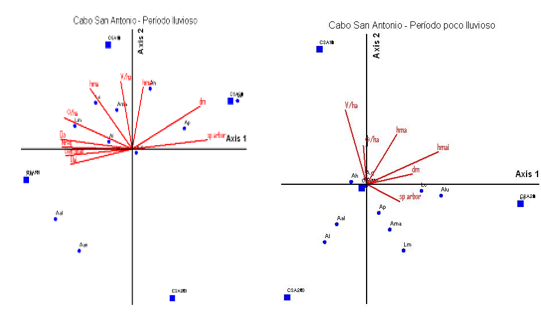 Nota: Las variables de vegetación se muestran como flechas. Las unidades de censo se muestran como círculos y los tratamientos de trabajo como cuadrados. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Área Basal, hma, Altura media del estrato arbustivo, hmai, Altura media del estrato arbóreo inferior, V ha-1, Volumen, Da, Densidad del estrato arbustivo, Dai, Densidad del estrato arbóreo inferior, Sp arbr, Riqueza de especies, Nreg, Regeneración natural.
Nota: Las variables de vegetación se muestran como flechas. Las unidades de censo se muestran como círculos y los tratamientos de trabajo como cuadrados. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Área Basal, hma, Altura media del estrato arbustivo, hmai, Altura media del estrato arbóreo inferior, V ha-1, Volumen, Da, Densidad del estrato arbustivo, Dai, Densidad del estrato arbóreo inferior, Sp arbr, Riqueza de especies, Nreg, Regeneración natural.Fig. 5 - Diagrama de ordenación de análisis de correspondencia realizado con la densidad especies de lagartos y las variables de vegetación en el sector Cabo de San Antonio
En la localidad Cabo Corrientes, la especie que manifiesta menos modificación con las variables de vegetación resultó ser Anolis sagrei, la cual no comparte su nicho ecológico en ninguno de los dos períodos con Anolis quadriocellifer. En análisis del período lluvioso, en el segundo cuadrante se muestra asociación de Anolis homolechis y Ameiva auberi con las variables de vegetación, volumen y densidad del estrato arbóreo inferior, mientras que Anolis loysina y Anolis allogus lo hacen con los mayores valores de área basal y diámetro de los árboles, en los tratamientos de más de 30 años de recuperación post aprovechamiento forestal.
Leiocephalus macropus en este período se encuentra alejado del resto de las especies mientras que en el período poco lluvioso prefirió los hábitats donde existe una mayor regeneración natural, que coincide con tratamientos de menor tiempo de recuperación post aprovechamiento forestal (Uvero Quemado).
Anolis porcatus se asocia a parcelas con mayores alturas y Anolis luteogularis a mayores densidades. Anolis homolechis tuvo un comportamiento diferente en este período respecto al anterior ya que en este caso seleccionó los hábitats de mayor riqueza de especies.
Las especies que resultaron alejadas de las demás fueron Ameiva auberi y A. alutaceus, estas dos especies tienen diferentes usos del recurso, la primera se observó ocupando el sustrato del suelo y las rocas, expuestas al sol directo o filtrado por la vegetación, la otra especie prefirió las bajas alturas y la sombra.
Anolis allogus se solapa con Anolis sagrei prefiriendo los lugares soleados del bosque y mayores alturas.
En el período poco lluvioso de esta localidad, Anolis allogus y Leiocephalus carinatus se asocian a parcelas de mayores volúmenes de los árboles, área basal y alturas, Ameiva auberi a los sitios más densos, Anolis luteogularis y A. alutaceus no presentan asociación con ninguna variable medida y Anolis angusticeps con las mayores alturas.
Anolis loysina y Anolis allogus se asocian con los mayores valores de área basal y diámetro de los árboles, en los tratamientos de más de 30 años de recuperación post aprovechamiento forestal, pero es este tiempo recuperativo el que lleva a que la formación vegetal se caracterice por el incremento de estos indicadores, pues después de haberse establecido la competencia y eliminación de los individuos menos adaptados, los restantes cuentan con mayores espacios, más homogéneamente distribuidos y así llegan a desarrollar mayores diámetros y áreas basales (Delgado y Ferro 2006).
Anolis porcatus y Anolis angusticeps se asocian a parcelas con mayores alturas y Anolis luteogularis a mayores densidades, todas son especies que frecuentan los sustratos más elevados en la formación vegetal (troncos y ramas).
En el período poco lluvioso, Anolis allogus y Leiocephalus carinatus se asocian a parcelas de mayores volúmenes de los árboles, áreas basales y alturas, mientras que Ameiva auberi frecuentó los sitios más densos y con las mayores alturas.
Los lagartos podrían mantener viable las poblaciones en los fragmentos aislados para más largo plazo que los pájaros y los mamíferos Hinsley et al., 1996; (Henle et al., 2004), garantizando su persistencia en los paisajes fragmentados a pesar de su habilidad de la colonización reducida.
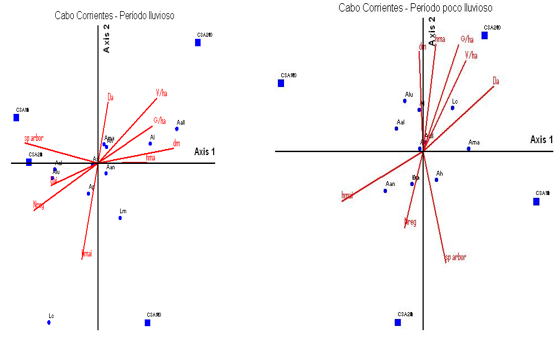 Nota: Las variables vegetación se muestran como flechas, las especies se muestran como círculos y los tratamientos de trabajo como cuadrados. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Área Basal, hma, Altura media del estrato arbustivo, hmai, Altura media del estrato arbóreo inferior, V ha-1, Volumen, Da, Densidad del estrato arbustivo, Dai, Densidad del estrato arbóreo inferior, Sp arbr, Riqueza de especies, Nreg, Regeneración natural.
Nota: Las variables vegetación se muestran como flechas, las especies se muestran como círculos y los tratamientos de trabajo como cuadrados. Aq, Anolis quadriocellifer*, Ah, Anolis homolechis*, Al, Anolis loysiana*, Aan, Anolis angusticeps, Ap, Anolis porcatus*, As, Anolis sagrei, Lc, Leiocephalus carinatus, Lm, Leiocephalus macropus*, Ama, Ameiva auberi, Aall, Anolis allogus*, Aal, Anolis alutaceus*, Alut, Anolis luteogularis*, Ls, Leiocephalus stictigaster*, G ha-1, Área Basal, hma, Altura media del estrato arbustivo, hmai, Altura media del estrato arbóreo inferior, V ha-1, Volumen, Da, Densidad del estrato arbustivo, Dai, Densidad del estrato arbóreo inferior, Sp arbr, Riqueza de especies, Nreg, Regeneración natural.Fig. 6 - Diagrama de ordenación de análisis de correspondencia realizado con la densidad de especies de lagartos y las variables de vegetación en el sector de Cabo de San Antonio. Cabo Corrientes
Al aplicar la prueba de Mantel se detecta como resultado que existe correlación entre las variables de vegetación en las dos localidades estudiadas (r= 0,86; P < 0,05) y las densidades de lagartos. Según los resultados expuestos, se puede establecer que existen grandes variaciones en las relaciones de las comunidades de lagartos con la estructura de la vegetación entre los períodos lluvioso y poco lluvioso.
El resultado de los análisis de correspondencias canónicas se debe a la selección de microhábitat como se muestra en las Figura 5 y Figura 6 podría ser un mecanismo explicando la persistencia de poblaciones de los lagartos en relación a las variables de la vegetación asociada y el tiempo de recuperación post aprovechamiento forestal de los bosques en estudio, ya que la selección del microhábitat puede variar en el espacio y tiempo respecto a las fuentes de alimentación, la reproducción, los depredadores y otros componentes de calidad del hábitat (Martín y López 2002). Coincidiendo con Caro (2010), en su revisión sobre los grupos de animales que pueden ser indicadores de varios disturbios antrópicos y naturales, señala que los reptiles podrían servir como indicadores del manejo forestal.
Con este estudio se ha podido puntualizar la relación de cada especie de lagarto con los diferentes elementos de vegetación y el tiempo recuperativo post aprovechamiento forestal para cada localidad de estudio, por lo que quedan definidos los indicativos para cada rasgo estructural y período de restablecimiento.
Las especies de este género, con un amplio nicho ecológico, son más probables a realizar desplazamientos ecológicos para alguna de las dimensiones del hábitat, en busca de refugio, alimento y mínimas condiciones de sobrevivencia. El estudio de los lagartos, en los últimos tiempos se ha enfocado a determinar los efectos que puedan causar los manejos forestales sobre el desplazamiento por alteraciones de la vegetación (Arias y Martínez 2014). Con ello, se intenta demostrar que los lagartos podrían ser indicadores de disturbios del manejo forestal y de otros impactos antrópicos (Hager 1998).
La riqueza de especies de lagartos y la riqueza de la vegetación asociados con los límites de rangos de la distribución han sido estudiados por diferentes autores (Schall y Pianka 1977, Bock et al., 1981 y Uetz 2000). Sin embargo, estos análisis no revelan las últimas causas de modelos de distribución de los mecanismos ecológico (Rojas et al., 2016).
Otros trabajos se han enfocado en las preferencias del hábitat de especies particulares o comunidades, a veces considerando, las relaciones entre los rasgos del hábitat y la actuación diferencial de individuos o poblaciones, tasa de crecimiento y supervivencia (Medina y Cardena 2015).
CONCLUSIONES
El uso de los recursos por las especies detectadas varió significativamente con el tiempo de recuperación post aprovechamiento forestal en las localidades de estudio para la altura y la iluminación. Además, tuvo una influencia en este indicador el período del año, mostrando el período poco lluvioso menor variación en la preferencia de las categorías establecidas en el uso del recurso.
Las variables de vegetación que mayor asociación mostraron con las diferentes especies de lagartos fueron la riqueza de especies, densidad del estrato arbóreo inferior y altura de los tres estratos establecidos.
Las asociaciones que se establecen entre los cambios de estructura del bosque y las comunidades de lagartos se manifiestan significativamente controlado por los estratos de la altura y la iluminación en ambas localidades y las variables de vegetación que tienen una mayor correspondencia son riqueza del estrato arbustivo, altura del estrato arbóreo superior y la regeneración natural.

