










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La diversidad de los moluscos terrestres del archipiélago cubano, se manifiestan tal vez como en ningún otro grupo de la fauna cubana, debido a los frecuentes procesos de especiación que ocurren en la mayoría de las islas. Evidencia de esto son las 476 especies terrestres de Neritimorpha y Caenogastropoda descritas para Cuba, con un endemismo del 99,1 % (Hernández et al., 2018).
De las seis familias de moluscos terrestres cubanos, actualmente ordenadas en Neritimorpha y Caenogastropoda, Annulariidae es la más extensa, con 352 especies agrupadas en 37 géneros (Hernández et al., 2018). Según el catálogo de Espinosa y Ortea (1999) esta familia en Granma está representada por 19 especies incluidas en cinco géneros (tres endémicos).
Los inventarios de malacofauna terrestre cubana disponibles hasta la fecha indican que Parachondria (Parachondria neglectus) (Pfeiffer 1858) presumiblemente constituye el único representante de esta familia en Manzanillo (Sariego 2010).
En esta zona geográfica, sin incluir las aguas interiores, la proporción cubierta de bosques con relación a la superficie terrestre total ha sido estimada en 27,1 % (ONEI 2018). Asimismo, la extensión de presencia para esta especie no excede 1 091 km² (Sariego 2006). Lo anterior ratifica la importancia de su estudio ecológico con fines conservacionistas.
Los moluscos terrestres, debido a la microlocalización y requerimientos ecológicos estrictos de muchas de sus especies, resultan ideales para evaluaciones sobre biodiversidad y estudios acerca de la historia de las comunidades de organismos en el espacio y el tiempo (Fontenla et al., 2013).
El nicho ecológico es un término de amplio uso en la teoría ecológica y evolucionaria. Es una concepción que intenta representar la función de los organismos en la naturaleza y sus interacciones con el ambiente (Fontenla 2001). Su estudio es de gran importancia para conocer las interacciones y las relaciones competitivas de las especies. Además, aporta elementos importantes para el manejo y conservación de la fauna. Según Silva y Berovides (1982) la determinación del nicho ecológico debe ser la unidad central de los estudios poblacionales, incluyendo los moluscos terrestres.
Para poder plantear proyectos de restauración o realizar tomas de decisiones en la conservación, resulta esencial disponer de información de base, así como de datos de referencia, para elaborar análisis temporales y predicciones con mayor solidez estadística. Las observaciones a largo plazo son necesarias para responder a cuestiones teóricas de la ecología, tales como reducir las incertidumbres sobre la irreversibilidad de los cambios estructurales y garantizar el éxito de los programas de restauración y conservación (Díaz-Delgado, 2016).
Por lo anteriormente expuesto, en la presente investigación se caracterizó la dinámica temporal del subnicho estructural de una población de P. neglectus en el municipio de Manzanillo perteneciente a la provincia de Granma, Cuba.
MATERIALES Y MÉTODOS
El municipio de Manzanillo ubicado en la provincia de Granma posee una extensión superficial de 498,95 km2 (ONEI 2017) y se encuentra en la llanura del Cauto-Guacanayabo. La ciudad está bordeada por alturas calizas de hasta 80 msnm y el extremo sur del territorio municipal se adentra en la precordillera de la Sierra Maestra (Sariego 2010). La localidad estudiada está situada entre 20°20’29.77’’N -77°05’00.37’’ W, pertenece al poblado Blanquizal y presenta un área aproximada de 20 500 m2. Su caracterización florística fue realizada en el herbario del Centro Oriental de Ecosistemas y Biodiversidad (BIOECO) y forma parte de los resultados del presente estudio. Para la ordenación y nomenclatura de las especies y familias botánicas se siguieron los criterios de Greuter y Rankin (2016).
Subnicho estructural de P. neglectus
La terminología utilizada para el estudio del subnicho estructural es la sugerida por Silva y Berovides (1982), teniendo en cuenta las subdivisiones propuestas por Berovides et al. (1988). La presencia del peristoma replegado permitió la identificación de los adultos. Para caracterizar la dinámica temporal del subnicho estructural de la población de P. neglectus se efectuaron dos muestreos mensuales (en parcelas de 3 x 3 m) durante el período comprendido entre diciembre de 2013 y febrero de 2015.
Sustratos de reposo diurno
Fueron cuantificados los ejemplares observados en el suelo y en el tronco de árboles y arbustos de la localidad para conocer el sustrato preferencial de la población, así como su frecuencia de utilización mensual.
Distancia al tronco
En todos los ejemplares que utilizaron el suelo como sustrato, se determinó la distancia al tronco (en cm) con un flexómetro Thorsen profesional de tres metros, la cual fue medida desde la región de la concha más cercana al árbol hasta la base del mismo.
Altura sobre el suelo y estratos
En los moluscos que se observaron en el tronco de árboles y arbustos, se determinó la altura sobre el suelo en centímetros. Los datos individuales de las alturas sobre el suelo fueron agrupados en las siguientes clases o estratos: I- (0,1 a 50 cm), II- (50,1 a 100 cm), III- (100,1 a 150 cm), IV- (150,1 a 200 cm), V- (200,1 a 250 cm), VI- (250,1 a 300 cm), VII- (300,1 a 350 cm) y VIII- (350,1 a 400 cm).
Relación planta-animal
El estudio se efectuó a través de la observación directa de los individuos y de las especies de plantas que servían de sustrato. Se realizaron líneas de recorrido para determinar las plantas utilizadas por P. neglectus. Para el estudio sobre la dinámica estacional de la disponibilidad y el uso de las especies vegetales fue seguida la metodología utilizada por Reyes-Tur (2004). Durante cada muestreo se seleccionaron en toda el área de trabajo 50 puntos al azar. En cada punto, se situó un listón de 3 m de largo y 5 cm de ancho. Se anotaron los contactos de dicho listón con las diferentes especies vegetales disponibles empleadas como substrato de reposo. Adicionalmente, para determinar la cantidad de veces que fue empleada cada planta, se consideraron observaciones realizadas dentro del área de trabajo, pero fuera de los puntos seleccionados. A partir de los datos recogidos se calculó el índice C (Pearre 1982) para las especies vegetales utilizadas como substrato por P. neglectus. Este índice fue calculado según la fórmula (Ecuación 1).
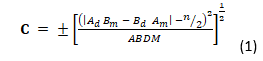
Donde Ad es la cantidad de individuos de la especie A empleada como substrato de reposo; Am es la cantidad de individuos de la especie A disponible; Bd es la cantidad de individuos del resto de las especies empleadas como substrato de reposo; Bm es la cantidad de individuos del resto de las especies disponibles; A es igual a Ad + Am; B es igual a Bd + Bm; D es igual a Ad + Bd; M es igual a Am + Bm y n es igual a Ad + Bd+ Am+ Bm.
El índice C, basado en χ 2 permite establecer la significación del grado de preferencia o rechazo para cualquier tamaño de muestra. Su valor oscila entre -1 y +1, los valores positivos indican preferencia, los valores negativos indican rechazo y C = 0 indica uso al azar. El criterio de significación para esta prueba fue P < 0,05. Se utilizaron los valores tabulados de χ2 con un grado de libertad según las sugerencias de Pearre (1982), Pearre (2011).
Los valores de este índice se determinaron teniendo en cuenta períodos lluviosos y poco lluviosos. Para establecer ambos períodos se analizaron los valores promedio mensuales de los milímetros de lluvia reportados por la Estación Meteorológica de Manzanillo y publicados por ONEI (2017). El estudio abarcó los siguientes períodos: Período poco lluvioso (diciembre de 2013 a abril de 2014), período lluvioso (mayo a octubre de 2014), período poco lluvioso (noviembre de 2014 a febrero de 2015).
Todos los cálculos estadísticos se realizaron con el programa STATISTICA 13.0 (Statsoft 2015). A través del cálculo de la media, la desviación estándar y el coeficiente de variación fueron caracterizadas estadísticamente las distancias al tronco y las alturas sobre el suelo de los moluscos. Se efectuó una prueba de Independencia con la realización de una tabla de contingencia 2 x 2 con corrección de Yates para conocer si en la población estudiada la frecuencia de utilización de un sustrato u otro depende de la ocurrencia o no de las precipitaciones. La prueba de Contraste de Kolmogorov-Smirnov se empleó para el análisis de las distribuciones por sustratos tanto en los individuos juveniles como en los adultos. La prueba U de Mann-Whitney permitió comparar mensualmente las distancias al tronco entre los individuos juveniles y adultos, así como las distancias al tronco y las alturas sobre el suelo alcanzadas por esta especie en los períodos lluviosos y poco lluviosos.
RESULTADOS Y DISCUSIÓN
Sustratos de reposo diurno
En general, el 83,9 % de los individuos se detectaron en el suelo, alrededor de la base de árboles y arbustos, el resto empleó los troncos como sustrato de reposo diurno. La frecuencia mensual de esta especie en ambos sustratos se muestra en la Figura 1. Aunque la utilización del suelo fue superior, en octubre de 2014 se registró la proporción más elevada de individuos en este sustrato (Figura 1).
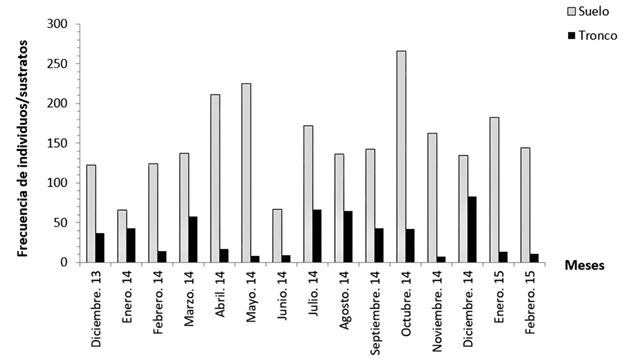
Fig. 1. - Frecuencia mensual por sustratos de individuos de la población estudiada de P. neglectus en Manzanillo
La prueba de Contraste de Kolmogorov-Smirnov indicó la existencia de diferencias estadísticas significativas entre las distribuciones de los juveniles en suelo y tronco (DN = 0,87; P = 0,0003). En los adultos, también existieron diferencias estadísticas significativas durante el análisis de distribuciones por sustratos (DN = 0,82; P = 0,0001). Sin embargo, no se encontraron diferencias estadísticas en la utilización que le daban al suelo tanto los juveniles como los adultos (DN = 0,45; P = 0,18). Tampoco existieron diferencias estadísticas significativas en la utilización del tronco por ambos grupos etarios (DN = 0,37; P = 0,24).
La prueba de independencia realizada mediante una tabla de contingencia 2×2 con corrección de Yates demostró que en la población la frecuencia de utilización de un sustrato u otro depende de la ocurrencia o no de las precipitaciones (χ2 = 214,88; P = 0,0001).
Durante el trabajo de campo nunca fueron observados moluscos de Parachondria neglectus en las piedras, como los encontró Gundlach en este mismo municipio, según Arango (1878) refiriéndose al sinónimo Chondropoma revinctum (Poey in Pfeiffer 1858). El género Chondropoma ya fue reportado por Mesa (1986) dentro de la clasificación de los moluscos arborícolas de Cuba. En el libro de moluscos terrestres de Cuba de Espinosa y Ortea (2009), posterior a la revisión más actualizada de moluscos terrestres de esta familia en el Caribe (Watters 2006) se menciona esta especie con el nombre de Chondropoma neglectum (Gundlach in Pfeiffer 1856). No obstante, estos autores reportan que muchas especies de moluscos terrestres son capaces de adaptarse a la ocupación temporal del espacio según las necesidades vitales de su ciclo biológico, la tierra, las piedras y la vegetación pueden formar parte de su ciclo de vida.
Sustrato suelo
Distancia al tronco
La media de la distancia al tronco fue 13 ± 6,42 cm. El 22,7 % de los moluscos se observaron alrededor y al pie de la base de árboles y arbustos. La variación temporal de las distancias al tronco de juveniles y adultos (Tabla 1).
Tabla 1. - Variación temporal de las distancias al tronco (cm) de individuos juveniles y adultos de la población estudiada de P. neglectus en Manzanillo
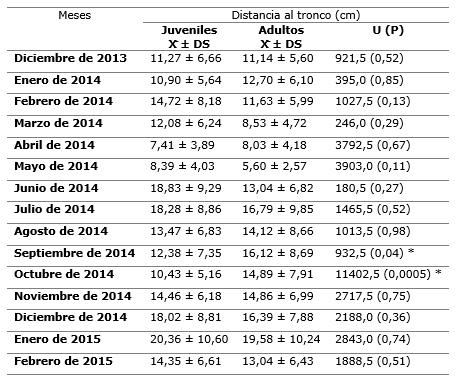
X: Media; DS: Desviación estándar; U: Prueba U de Mann-Whitney con la probabilidad asociada. *: Si P< 0,05 se considera significativa la prueba.
Solo en los meses de septiembre y octubre de 2014 las distancias al tronco entre ambos grupos etarios fueron estadísticamente diferentes. Los resultados de la Prueba U de Mann-Whitney indicaron que, en los períodos lluviosos y poco lluviosos, las distancias al tronco para la población no fueron estadísticamente significativas (U = 472; P = 0,11).
Según Sariego et al. (2018) para esta especie el tronco de los arbustos y árboles es el sustrato preferencial para su reproducción. Estos autores informaron que en los meses de septiembre y octubre ocurrió el período de reclutamiento en la población. De lo anterior, es posible inferir que las diferencias estadísticas significativas entre las distancias al tronco de los juveniles y los adultos. Estos dos meses pudieran indicar que la eclosión de los huevos ocurre con mayor éxito en condiciones favorables en la base de árboles y arbustos.
Sustrato tronco
Altura sobre el suelo y estratos
La altura sobre el suelo presentó un valor medio de 153,5 ± 71,3 cm y un máximo de 394 cm. Durante los días lluviosos aumentó la frecuencia de moluscos de P. neglectus en el tronco. Se observó también un mayor número de individuos activos en este sustrato (Figura 2).

Fig. 2. - Imagen ilustrativa de la morfología externa de P. neglectus durante la actividad diurna en el tronco de Swietenia mahagoni favorecida por las lluvias en la localidad
Los juveniles mostraron preferencia por el estrato más bajo (55,6 %) y ocuparon cinco de los ocho estratos establecidos. El 25 % de los adultos se concentró en el estrato IV. De manera general, los adultos se distribuyeron en el tronco ocupando todos los estratos (Figura 3). Las alturas sobre el suelo alcanzadas por esta especie en los días lluviosos y no lluviosos mostraron diferencias estadísticamente significativas (U = 21519,5; P < 0,001) (Figura 3).
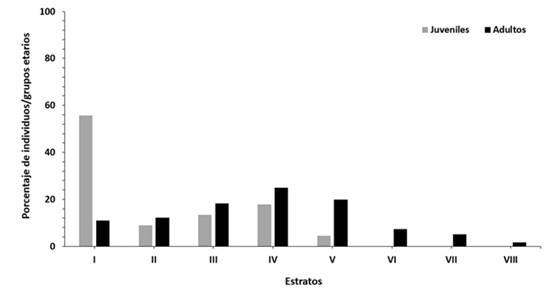
Fig. 3. - Porcentaje de individuos juveniles y adultos por estratos en el tronco de árboles y arbustos utilizados por P. neglectus en la localidad estudiada
Durante el reposo diurno se observaron individuos que se adhirieron al tronco con una banda corta de mucus seco (no mayor de dos milímetros) con visibles pliegues longitudinales prolongados desde el sustrato por la superficie interior de la concha hasta el opérculo. Según Watters (2014), la propensión de moluscos de esta familia a suspenderse de hilos mucosos se ha reportado para especies de Jamaica, Haití, República Dominicana, las Antillas Holandesas y Cuba. El autor planteó, además, que este comportamiento pudiera indicar un rasgo muy antiguo como estrategia antidepredadora o una estrategia para disipar el calor.
Desde el punto de vista energético, la locomoción de los moluscos es lenta y compleja debido a la secreción de mucus. Teniendo en cuenta este criterio, es lógico interpretar que ante condiciones favorables de temperatura y humedad se incremente su actividad y ascenso por el tronco. Se ha reportado además que en algunas especies de moluscos terrestres los ciclos de actividad diarios y anuales están sincronizados e interactúan con diversos factores ambientales (Reyes-Tur et al., 2018).
Los períodos climáticos estudiados y establecidos según reportes de ONEI (2017) para Manzanillo coincidieron con los dos períodos estacionales bien definidos en todo el territorio cubano, uno lluvioso que abarca de mayo a octubre y otro poco lluvioso que se extiende de noviembre a abril (Durán 2016).
Relación planta-animal
El inventario florístico de la localidad estudiada permitió la identificación de 35 especies (19 familias) de plantas superiores (16 herbáceas, cinco lianas, una epífita, ocho arbustos y cinco árboles). Sin embargo, P. neglectus fue observada en 8 especies vegetales (cinco árboles y tres arbustos) como sustrato de reposo diurno.
Los valores del índice C mostraron preferencia significativa por Swietenia mahagoni durante todos los períodos climáticos muestreados. Para Cordia gerascanthus, Ehretia tinifolia y Bursera simaruba existieron preferencias estadísticamente significativas en el período lluvioso. En el período poco lluvioso para estas tres especies, así como para Gastrococos crispa, Bourreria virgata, B. taylorii y Pisonia aculeata (en todos los períodos climáticos) los valores del índice C no resultaron significativos (C, P > 0,05) (Tabla 2).
Los gastrópodos terrestres disponen de numerosas especies vegetales que se encuentran en su entorno, de las cuales pueden emplear el tronco como sustrato de reposo y/o alimentario, por lo cual son considerados habitualmente como herbívoros generalistas (Reyes-Tur, 2004).
Independientemente de este comportamiento generalista en relación con el uso de las plantas como sustratos de reposo, existen especies capaces de desarrollar estrategias de vida en cuanto a la preferencia y rechazo de plantas. Las características de dichas estrategias están relacionadas principalmente con la disponibilidad de especies vegetales y sus características físicas (morfo-anatómicas) (Reyes-Tur 2004).
En la localidad estudiada P. negletus mostró preferencia por tres especies arbóreas y un arbusto de porte grande. No obstante, la preferencia encontrada por plantas de gran talla no puede considerarse una peculiaridad de la especie. La misma ha sido observada en otras localidades en Manzanillo asociada a poáceas erguidas, así como a las siguientes especies de plantas: Amphilophium gnaphalanthum (liana), Turbina corymbosa (trepadora), Leucaena leucocephala (arbusto), Sideroxylon foetidissimum (árbol) y Cocos nucifera (árbol) (Sariego 2010) (Tabla 2).
Tabla 2. - Listado florístico de familias y especies con los valores del índice C (Pearre, 1982) para taxa vegetales utilizados por P. neglectus en la localidad estudiada
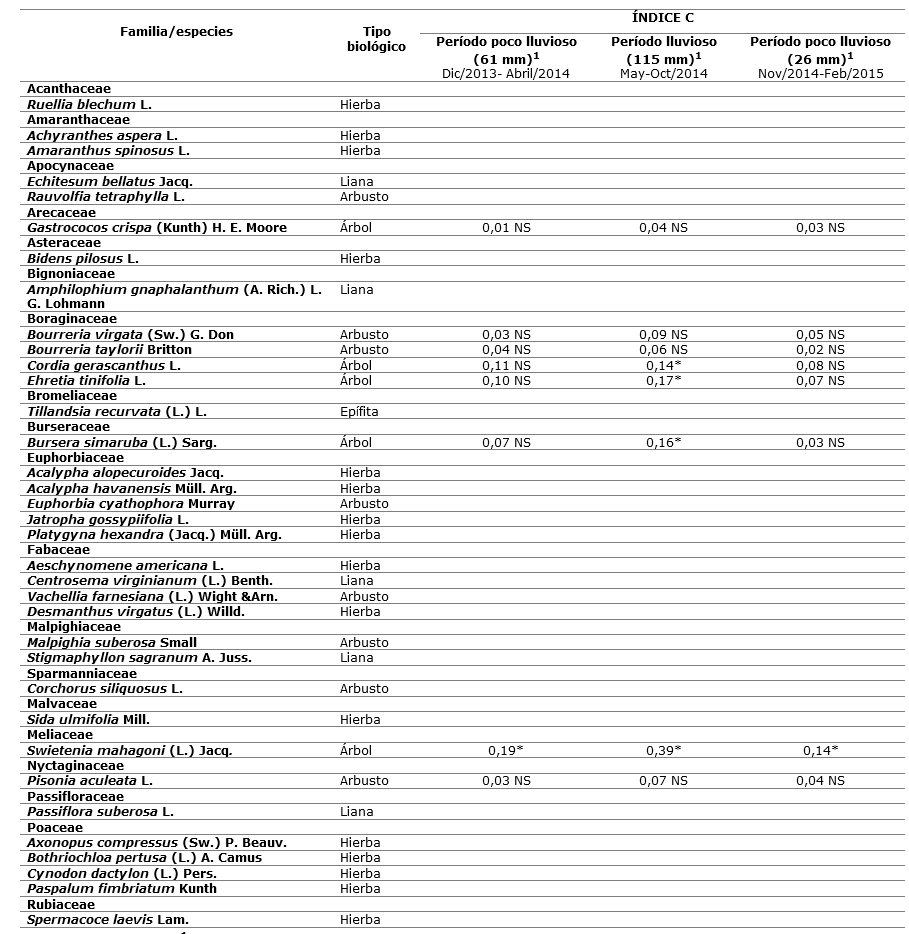
NS: no significación estadística; *: P < 0,05; 1: Promedio mensual de los mm de lluvia acumulados en Manzanillo en los períodos climáticos estudiados obtenidos de ONEI (2017).
CONCLUSIONES
De acuerdo a las observaciones realizadas del uso de sustratos en la localidad estudiada del municipio de Manzanillo, Granma; es posible inferir que P. neglectus es un molusco terrícola asociado a la vegetación.
En la población estudiada de P. neglectus, la frecuencia de utilización de un sustrato u otro dependió de la ocurrencia o no de las precipitaciones; ante condiciones favorables de temperatura y humedad estos moluscos incrementaron su actividad y ascenso por el tronco.
Los valores del índice C mostraron preferencia significativa de P. neglectus por Swietenia mahagoni (caoba) durante todos los períodos climáticos muestreados. Para Cordia gerascanthus, Ehretia tinifolia y Bursera simaruba existieron preferencias estadísticamente significativas en el período lluvioso.

