










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCultivos Tropicales
Print version ISSN 0258-5936
cultrop vol.33 no.1 La Habana Jan.-Mar. 2012
Influencia de tres fechas de siembra sobre el crecimiento y la relación fuente- demanda del cultivo del maíz (Zea mays L.).
Influence of three planting dates on growth and source-sink relationship of maize (Zea mays L.).
Naivy HernándezI y Dr.C. F. SotoII
IReserva Científica del Instituto nacional de CienciasAgrícolas, gaveta postal 1, San José de las Lajas, Mayabeque, Cuba,CP 32 700.
IIInvestigadorTitular del departamento de Fitotecnia, Instituto nacional de CienciasAgrícolas, gaveta postal 1, San José de las Lajas, Mayabeque, Cuba,CP 32 700.
RESUMEN
Con el objetivo de evaluar la influencia de tres fechas de siembra sobre el crecimiento y la relación fuente demanda en el cultivo del maíz (Zea mays), se desarrolló el presente trabajo en el Instituto Nacional de Ciencias Agrícolas; se estudiaron 3 fechas de siembra: noviembre de 2008, junio de 2009 y julio de 2009, utilizándose una densidad de 50000 plantas.ha-1. Se realizaron muestreos destructivos cada 15 días de emergidas las plantas hasta la cosecha, determinándoseles la masa seca total de la parte aérea y el Índice de Área Foliar (IAF), ajustándose los datos a una función exponencial polinómica de segundo grado, “x” fue los días después de la emergencia y “y” la variable en cuestión, a partir de las ecuaciones de regresión se calcularon la Tasa de asimilación neta (TAN) y la Relación de área foliar (RAF), también se calculó la relación entre la potencia de la fuente y la potencia de la demanda, encontrándose que una mayor eficiencia en el uso de la radiación conlleva a valores máximos más elevados en la tasa de asimilación neta (TAN) y en los potenciales entre los órganos fuente y demanda y por tanto a una mayor acumulación de biomasa en momentos próximos a la madurez fisiológica, destacándose en este caso en estudio la fecha de junio de 2009 la cual mostró una mayor eficiencia respecto a los índices evaluados alcanzándose en esta los mayores rendimientos.
Palabras clave: maíz, fechas de siembra, índices de crecimiento, relación fuente - demanda.
ABSTRACT
In order to evaluate the influence of three planting dates on growth and the demand supply ratio in the cultivation of maize (Zea mays), this work was developed at the National Institute of Agricultural Sciences, were studied 3 planting dates November 2008, June 2009 and July 2009, using a density of 50000 plants.ha-1. Destructive samplings were conducted every 15 days the plants emerged until harvest determined total dry mass of shoot and Leaf Area Index (LAI), adjusting the data to an exponential function of second-degree polynomial, "x" was days after emergence and y the variable in question, from the regression equations were calculated net assimilation rate (NAR) and leaf area ratio (LAR), also were calculated the ratio between the power Supply and demand power, finding that greater efficiency in the use of radiation leads to higher peaks in the net assimilation rate (NAR) and in potential between the supply and demand organs and therefore greater accumulation of biomass at a time close to physiological maturity, standing in this case study, dated June 2009 which showed a higher efficiency compared to rates reaching evaluated in this the greatest returns.
Key words: maize, sowing date, growth rates, source – sink relationship.
INTRODUCCIÓN
El crecimiento se define como un incremento constante en el tamaño de un organismo determinado por procesos de morfogénesis y diferenciación; el primero es el desarrollo de la forma o modelo de la célula u órgano, mientras que el segundo es el proceso por el cual las células cambian estructural y bioquímicamente para formar o adquirir funciones especializadas (1).
Según se plantea el crecimiento y la capacidad productiva de un cultivo es el resultado del genotipo, del ambiente que lo rodea y de su interacción. El genotipo es relativamente constante si se compara con la variabilidad del ambiente; sin embargo, la expresión fenotípica es ampliamente influenciada por los cambios ambientales y cualquier variable que produzca efectos sobre el medio va a verse reflejada en el crecimiento y productividad del cultivo (2).
La asimilación de materia seca y su distribución dentro de la planta, son procesos importantes que determinan la productividad del cultivo. El estudio de los patrones de asignación de materia seca hacia las diferentes partes de la planta, la variabilidad de estos patrones entre cultivares y el efecto de las condiciones ambientales en el proceso, pueden ayudar a maximizar la productividad y a seleccionar cultivares para un propósito particular (3).
Los asimilados producidos por la fotosíntesis en los órganos “fuente” (principalmente las hojas), pueden ser almacenados o distribuidos vía floema entre los diferentes órganos “demanda” de una planta. Para lograr un rápido crecimiento inicial de las plantas jóvenes, es importante un incremento substancial de la superficie foliar en la fase vegetativa, debido a que gran parte de la radiación solar incidente no es interceptada. Por lo tanto, en esta fase, una gran parte de los asimilados deben ser destinados a la formación de las hojas. (4).
El cultivo del maíz, es un importante cereal en la alimentación humana y animal. Además, por el uso que hace del agua, en diferentes condiciones de disponibilidad o déficit hídrico, tiene particularidades importantes en el manejo tecnológico (5). Al mismo tiempo es una de las especies con un elevado potencial de rendimiento asociado con altos niveles de fotosíntesis (2).
Sin embargo los factores climáticos pueden influir en la distribución a corto plazo de asimilados, como consecuencia de la respuesta de la potencia de demanda de los órganos individuales a los cambios de las condiciones externas, y también, a largo plazo, a través del efecto que ejercen sobre el número de órganos sumideros que crecen en la planta (4). Adicionalmente, el peso seco de la parte aérea de la planta de maíz depende de la cantidad de radiación fotosintéticamente activa (RFA) interceptada por el cultivo, la cual también puede variar con la temperatura (6).
Por lo que las variaciones en la fecha de siembra afectan de manera importante el crecimiento y desarrollo del cultivo, ya que colocan a las distintas etapas de ge¬neración del rendimiento bajo diferentes condiciones de radiación, temperatura y precipitaciones (7). De acuerdo con ello el presente trabajo tuvo como objetivo evaluar la influencia de tres fechas de siembra sobre el crecimiento y la relación fuente demanda del cultivo del maíz (Zea mays L.).
MATERIALES Y MÉTODOS
El trabajo se desarrolló en el área experimental del Instituto Nacional de Ciencias Agrícolas (INCA), ubicado en San José de las Lajas, Mayabeque, situado a 138 m snm. Se establecieron tres fechas de siembra: 18 de noviembre de 2008, 12 de junio de 2009 y 31 de julio de 2009, sobre un suelo ferralítico rojo (8), siguiendo las recomendaciones para la producción del cultivo (9). Los experimentos se extendieron hasta los 104, 76 y 90 días respectivamente para cada fecha de siembra hasta alcanzar la madurez fisiológica. El maíz var. P79-28 se estableció a 0.90 x 0.30 m, obteniéndose una densidad de siembra de 50000 plantas.ha-1. Semanalmente y a partir de los 15 días después de la emergencia (DDE) se determinó mediante muestreos destructivos, los siguientes indicadores.
1. La masa seca de la parte aérea, para la cual se separaron los diferentes órganos de la planta y se secaron en una estufa de circulación forzada a 800C, hasta peso constante.
2. Índice de Área Foliar (IAF), para ello se estimó el área foliar por el método del disco en base a masa seca.
3. Rendimiento en grano seco.
Los datos de los indicadores de crecimiento se ajustaron a una función exponencial polinómica de segundo grado, calculándose a partir de esto, la Tasa de Asimilación Neta (TAN) de la masa seca y la Relación de Área Foliar (RAF).
Adicionalmente, se determinó la fuerza o potencia de fuente y de demanda de cada fecha de siembra a lo largo del ciclo del cultivo, utilizando las fórmulas (10).
Potencia de la fuente = Tamaño de la fuente (Área Foliar)*Actividad de la fuente (TAN).
Potencia de la demanda = Tamaño de la demanda (Materia Seca Mazorca)*Actividad de la demanda (TRC Mazorca).
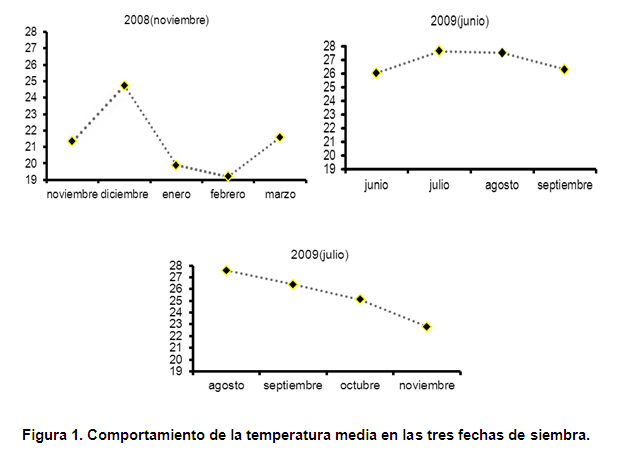
Los datos de temperatura media del aire se tomaron de la estación meteorológica de Tapaste, próxima al sitio experimental, no teniéndose en cuenta las precipitaciones ya que al mismo se le aplicó riego siempre que fue requerido. (Figura 1)
RESULTADOS Y DISCUSIÓN
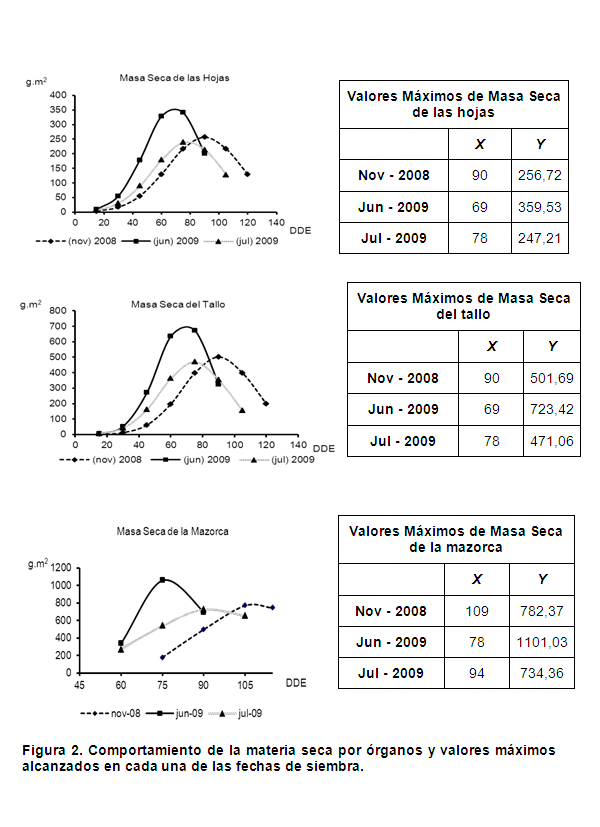
La figura 2 muestra el comportamiento de la masa seca por órganos de la planta (hoja, tallo y mazorca) en cada una de las fechas de siembra evaluadas, como se puede observar el mayor acumulado de masa seca de las hojas se presentó en la fecha de siembra de junio de 2009, con 359,53 g.m-2 a los 69 días después de la emergencia (estado masoso del grano), seguida de las fechas de siembra de noviembre de 2008 y julio de 2009 las cuales mostraron los mayores acumulados de masa seca de las hojas a los 90 y 78 días después de la emergencia respectivamente, correspondiendo a la fase fenológica de estado pastoso del grano. Resulta interesante evidenciar como la siembra de junio de 2009 alcanzó un mayor desarrollo del follaje o aparato fotosintético, en un menor período de tiempo y en una fase fenológica más próxima a la cosecha que el resto de las siembras.
Por otro lado, al observar el acumulado de materia seca de los tallos a lo largo del ciclo del cultivo, se puede observar un comportamiento similar al de la masa seca de las hojas pues los valores máximos se obtuvieron en la fecha correspondiente a jun-2009 con 723,53 g.m-2 a los 69 días después de la emergencia, seguida de las fechas de siembra de nov-2008 y jul-2009, como se muestra en la tabla de valores máximos. Se plantea que el peso seco de la parte aérea de la planta de maíz depende de la cantidad de radiación fotosintéticamente activa interceptada por el cultivo, la cual también puede variar con la temperatura (6).
Las curvas de acumulado de masa seca de las mazorcas muestran que a los 78 días los valores máximos alcanzados correspondieron a la fecha de siembra de junio de 2009 con 1101,53 g.m-2, seguida de las fechas de siembra de noviembre de 2008 y julio de 2009. Este comportamiento podría explicarse debido a que las mayores temperaturas se presentaron en la fecha de siembra de jun-2009 donde se obtuvieron los mayores acumulados de masa seca y en un menor período de tiempo, mientras que el maíz sembrado en nov-2008 estuvo expuesto a temperaturas más bajas durante todo el ciclo del cultivo, no siendo así en la fecha de jul-2009, donde durante los primeros estadios las plantas estuvieron sometidas a altas temperaturas, disminuyendo progresivamente durante el resto del ciclo del cultivo (figura 1).
Por otro lado estudios realizados al cultivo indican que el maíz por ser una especie de metabolismo C4, tiende a expresar su máxima productividad cuando la etapa de máxima área foliar coincide con una mayor disponibilidad de radiación solar, siempre que no haya déficit hídrico (11).
Diferentes autores señalan que las mayores temperaturas aceleran la velocidad de aparición de las hojas. El aumento en la velocidad de emergencia y despliegue de las hojas determina el rápido establecimiento de un canopeo eficiente en capturar la luz solar. Este efecto, sumado a los niveles crecientes de radiación solar incidente hacia el verano, le permite a las siembras tardías acumular una cantidad de radiación interceptada hasta la floración semejante a la de las siembras tempranas, a pesar del menor número de días transcurridos. Las mayores temperaturas durante la fase vegetativa que experimentan las siembras tardías favorecen una alta eficiencia de conversión de luz en crecimiento, acumulando generalmente más biomasa al momento de floración que las siembras tempranas (12).
También se plantea que para la mayoría de los cultivos, el rendimiento está positivamente relacionado con la duración del período del llenado del grano; los factores genéticos, climáticos y de manejo que permiten una mayor duración y un mantenimiento de la fotosíntesis durante este período son fundamentales en su aporte a un incremento del rendimiento final. Se puede deducir que una planta que alcance una mayor acumulación de biomasa en el período de floración dará un mayor rendimiento (13).
Por otro lado se plantea que el mayor rendimiento agrícola está asociado a un crecimiento más precoz y de mayor magnitud, atendiendo a la masa seca de la parte aérea de las plantas por unidad de superficie (14).
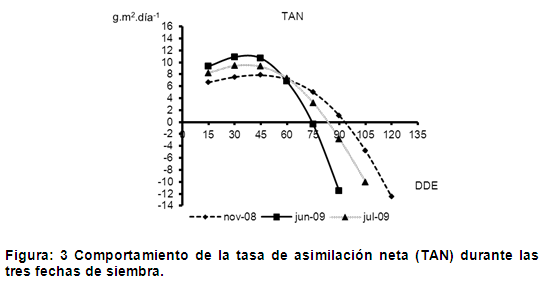
La tasa de acumulación de materia seca por unidad de área foliar y tiempo se define como la tasa de asimilación neta (TAN), que es una medida de la eficiencia media fotosintética de las hojas (15 y 16). En la figura 3 se presenta el comportamiento de este indicador de eficiencia fotosintética promedio, el cual presentó los mayores valores al comienzo del ciclo de cultivo (hasta 45 días después de la emergencia) en las tres fechas de siembra, debido a que las plantas se encontraban en el inicio de la etapa fenológica de desarrollo de hoja.
Después de los 45 días la TAN disminuyó progresivamente en las tres fechas de siembras, sin embargo como se muestra en dicha figura, los valores mínimos se alcanzaron primeramente en las fechas de junio y julio de 2009 a los 75 y 90 días después de la emergencia respectivamente y los 105 días se alcanzaron estos valores para la siembra de noviembre de 2008. Este comportamiento pudo deberse a que durante la fecha de junio de 2009 las plantas estuvieron sometidas a mayores temperaturas por lo que el crecimiento fue más acelerado, completando su ciclo en un menor periodo de tiempo respecto al resto de las siembras, no siendo así para julio de 2009 donde las plantas estuvieron primeramente sometidas a temperaturas más elevadas y luego disminuyeron progresivamente durante el ciclo del cultivo, mientras que para noviembre de 2008 las temperaturas se mantuvieron bajas durante todo su ciclo, ocurriendo un crecimiento más prolongado.
La disminución de la TAN después de alcanzar el máximo valor, indica que la capacidad fotosintética del dosel tiende a disminuir continuamente con el crecimiento y expansión del área foliar y puede cesar en el momento en que las plantas alcanzan su máxima área foliar, debido principalmente, al sombreo que ocurre entre las hojas de los diferentes estratos del dosel, lo cual ocasiona que la respiración tienda a ser mayor que la fotosíntesis (17).
Por otro lado, se observa que la fecha de siembra de junio de 2009 presentó una mayor eficiencia fotosintética, mostrando un incremento mayor respecto al resto de las fechas de siembra, correspondiendo esta alta eficiencia fotosintética a la fase fenológica de formación de la mazorca, seguida de la siembra de julio de 2009 donde su máxima tasa de asimilación neta se alcanzó en la formación de la espiga y en el último extremo, con una TAN menor, la fecha de noviembre de 2008, donde su máxima eficiencia fotosintética correspondió a la fase fenológica de formación de las 10 hojas. Estas diferencias en los valores máximos de la TAN y el momento de ocurrencia podrían estar dados por la diferencia de temperatura y la radiación interceptada, la cual alcanza, en Cuba, los mayores valores en los meses de junio y julio oscilando entre los 19 y 20 MJ.m-2 (18), además de la duración del cultivo durante cada fecha de siembra.
Según estudios realizados se plantea (19) que cada especie responde de manera particular a los estímulos de radiación que recibe del ambiente, siendo diferentes estas respuestas incluso dentro de una misma especie en diversas etapas fisiológicas del desarrollo. Resultados de este tipo fueron reportados, encontrando en la variedad ‘Río Grande’ de tomate una TAN mayor en los primeros estados de desarrollo de plantas expuestas a radiación total, condición que se hizo contraria en los estados más avanzados de crecimiento (20).
Los valores máximos de TAN implican mayor producción de materia seca por unidad de área foliar y por unidad de tiempo, lo que se traduce en una mayor eficiencia fotosintética. En estudios realizados a dos genotipos de Stevia rebaudiana se registró que la diferencia mayor de área foliar fue más marcada cuando estuvo creciendo bajo niveles altos de radiación; por lo tanto, se podría decir que la mayor cantidad de biomasa que envió este genotipo al órgano de interés, se explica por un desarrollo mayor de su lámina fotosintética, sumado a su mayor eficiencia en la síntesis de fotoasimilados por unidad de área (19).
Por otra parte en trabajos realizados con maíz bajo condiciones óptimas de crecimiento, atribuyen principalmente las diferencias en la acumulación de biomasa observadas entre los años de crecimiento a diferencias en el uso eficiente de la radiación, que varía debido a las diferencias en el desarrollo del área foliar (21). Por otra parte, investigaciones realizadas en lechuga (Lactuca sativa L.) mostraron que la tasa de asimilación neta ponderada a través de todo el ciclo del cultivo se redujo al disminuir el nivel de radiación incidente (22).
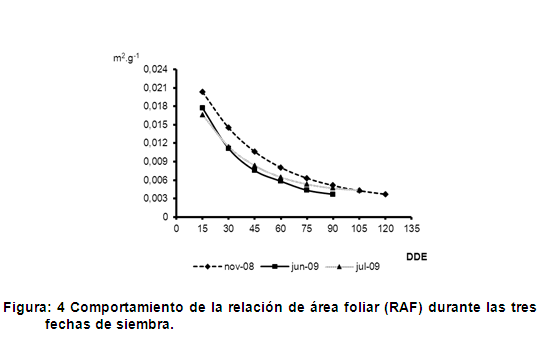
El criterio planteado con anterioridad coincide con lo que se observa en la figura 4 donde se presenta la relación de área foliar (RAF) en las tres fechas de siembras, la cual es una medida del balance entre la capacidad fotosintética potencial y el costo respiratorio potencial. En esta figura se muestra que las plantas expuestas a temperaturas más bajas y por tanto menor radiación incidente presentaron los valores más altos de RAF, correspondiendo con la siembra de noviembre de 2008, en tanto las siembras expuestas a temperaturas más elevadas durante el ciclo del cultivo y por tanto mayores radiaciones incidentes mostraron los más bajos valores de RAF.
Los anteriores resultados indican que las bajas radiaciones estimularon un incremento de la relación de área foliar en las plantas. Esto demuestra que estas plantas hacen una mayor inversión en tejido foliar, posiblemente para lograr una mayor captación de luz. Por otra parte, las plantas expuestas a bajas radiaciones presentaron resultados bajos de área foliar y materia seca de hojas evidenciando el lento crecimiento y la ineficiencia fotosintética bajo estas condiciones, lo que representa una limitante para expresar su máximo rendimiento.
La tendencia encontrada coincide con resultados obtenidos sobre aclimatación de Panicum maximum a diferentes regímenes lumínicos (10%, 30% y 100%), en los que la RAF se incrementó a medida que la radiación ambiental disminuyó, con incrementos de 83% y 121% cuando las plantas se expusieron a 30% y 10% de radiación ambiental, respectivamente (23).
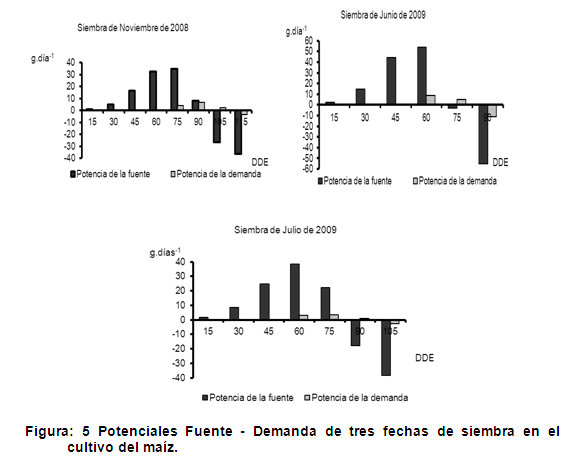
La figura 5 corresponde a la determinación de los potenciales de fuente y demanda, de las hojas y las mazorcas respectivamente, como se puede observar la siembra de junio de 2009 presentó una mayor potencia de la fuente, donde a los 60 días después de la emergencia expresó su máximo potencial con 53,76 g.día-1, sin embargo las fechas de siembra de noviembre de 2008 y julio de 2009 no mostraron diferencias entre sí, ya que los máximos potenciales de la demanda se obtuvieron a los 75 y 60 días con valores de 34,68 g·día-1 y 38.32 g·día-1 respectivamente. Estos resultados de potencia de fuente se pueden interpretar como el momento en el cual la maquinaria fotosintética fue más eficiente en este proceso metabólico y las redes vasculares fueron más eficientes en la carga floemática de azúcares (24).
Por otro lado, la potencia de la demanda como se muestra en dicha figura, la fecha de siembra de junio de 2009 mostró los mayores valores máximos a los 60 días con 8.63 g.día-1, mientras que las siembras de nov-2008 y jul-2009 presentaron los valores máximos a los 90 y 75 días con 6.65 y 3.59 g.día-1 respectivamente. Como se evidencia en los resultados obtenidos, entre las fechas de siembra existe un comportamiento diferente en cuanto a la potencia de los órganos demanda, lo cual pudiera deberse a que la habilidad de los órganos demanda para obtener asimilados puede estar relacionada con la rápida división y expansión celular, que se refleja en una fase lineal de acumulación de materia seca de estos órganos de la planta. Este proceso implica una mayor descarga floemática de fotoasimilados en este tejido vertedero y por ende una mayor fuerza como tejidos “demandantes” de carbohidratos y otras sustancias orgánicas (25).
Varios autores sugieren que las altas tasas de translocación son un importante factor en el mantenimiento de altas tasas fotosintéticas en muchas plantas (24), de igual forma una inadecuada utilización de fotoasimilados y bajas tasas de translocación pueden resultar en una reducción de la fotosíntesis, manifestando una estrecha relación entre la potencia de fuente y la potencia de demanda.
Por lo anterior, es de esperar que las fechas de siembra evaluadas presenten periodos en los cuales los valores de potencia de fuente y potencia de demanda sean muy similares, indicando habilidades equivalentes para producir asimilados en las hojas y para obtenerlos en las mazorcas. En el caso de la fecha de siembra de junio de 2009 este se presentó a los 60 días donde se obtuvieron los mayores potenciales de fuente y demanda, mientras que en las fechas de nov-2008 y jul-2009 ocurrió en el periodo correspondiente a los 75-90 días y 60-75 días después de la emergencia, según se plantea este fenómeno tiene implicaciones a nivel agronómico si se tiene en cuenta que una disminución en el área foliar ocasionada por problemas fitosanitarios, puede resultar en baja potencia de fuente y así disminuir el llenado de las mazorcas lo que pueden conducir a bajos rendimientos (25).
Como se muestra en dicha figura, en la fecha de siembra de jun-2009, después de los 75 días, se observó que la potencia de la fuente en este intervalo de tiempo es más baja en comparación con la potencia de la demanda, incluso el follaje se encuentra en etapa de senescencia en este periodo del ciclo de cultivo, por lo que se presume la posibilidad de un almacenamiento temporal en los tallos antes de la acumulación final de asimilados en las mazorcas, en la etapa final del ciclo de cultivo. Además una característica de varias plantas que se ha planteado con anterioridad (24), es la habilidad para almacenar temporalmente metabolitos fotosintéticos y nutrientes en tejidos a lo largo de la vía de transporte entre la fuente y la demanda, quedando disponible para un uso posterior en la planta ya sea porque la demanda se incrementa o porque la disponibilidad de fotoasimilados se reduce en un momento dado.
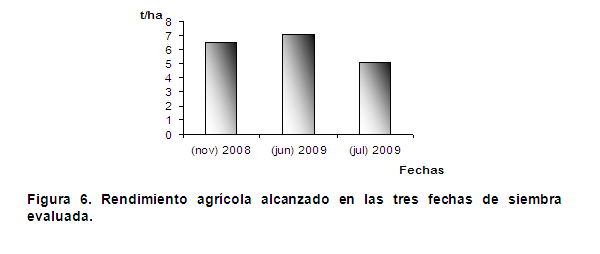
Al observar la figura 6 se muestran los rendimientos obtenidos durante las tres fechas de siembra, evidenciándose que los mayores rendimientos corresponden a la siembra de jun-2009, seguida de las siembras de nov-2008 y jul-2009. Demostrando que una mayor eficiencia en el uso de la radiación conlleva a valores máximos más elevados en la tasa de asimilación neta (TAN) y en los potenciales entre los órganos fuente y sumidero y por tanto a una mayor acumulación de biomasa en momentos próximos a la madurez fisiológica alcanzándose mayores rendimientos. Resultados similares fueron obtenidos en estudios realizados a la influencia de la radiación solar en la producción de semillas de papa, donde a menor radiación se disminuyó la eficiencia fotosintética de la planta, lo cual repercutió directamente en la biomasa del vástago y tubérculo; así como en los índices de eficiencia medidos (26).
REFERENCIAS
1. Taiz, L. y E. Zeiger. 2006. Plant physiology. Sinauer Associates Inc., Sunderland, MA.
2. Rincón, A., Ligarreto, G. y Sanjuanelo, D. Crecimiento del maíz y los pastos (Brachiaria sp.) establecidos en monocultivo y asociados en suelos ácidos del piedemonte llanero colombiano. Agron. Colomb, 2007, vol. 25, no. 2, pp. 264-272.
3. Ñústez, C. E., Santos, Marcela y Segura, Mariela. Dry matter allocation and partitioning of four potato varieties (Solanum tuberosum L.) in zipaquirá, cundinamarca (Colombia). Rev. Fac. Nal. Agr. Medellín, 2009, vol. 62, no. 1, pp. 4823-4834.
4. Piel, Roberta. M. y Galvez, J. L. Dry-matter partitioning as a determinant of greenhouse fruit Vegetable crops production. R. bras. Agrociência, 2005, vol.11, no. 1, pp. 05-11.
5. Balbi, Celsa. N., García, P. A. y Ferrero, A. R. 2005. Intercepción de la y radiación y materia seca acumulada en maíz y sorgo en la provincia de Corrientes. Universidad del Nordeste. Comunicaciones científicas y Tecnológicas. [Consultado: feb.2010]. Disponible en:< http://www.unne.edu.ar/Web/cyt/com2005/5-Agrarias/A-023.pdf >
6. García, P., Cabrera, S. y Sánchez, J. Rendimiento del maíz y las épocas de siembra en los LLanos Occidentales de Venezuela. Agronomía Tropica, 2009, vol. 59, no. 2, pp. 161-172.
7. Castellarín, P.H., Ferraguti, J.M., y Rosso, O. 2009. Fechas de siembra y rendimientos de maíz en Oliveros (Santa Fe), campaña 2008/09. Manejo de Cultivos. EEA Oliveros INTA. [Consultado: jun.2011]. Disponible en:< http://www.inta.gov.ar/oliveros/info/revistas/Ma%C3%ADz%202009/3%20Fechas%20de%20siembra%20y%20rendimientos%20de%20ma%C3%ADz%20en%20Oliveros.pdf >
8. Hernández, A., Morell, F., Ascanio, M. O., Borges, Y., Morales, M. y Yong, A. Cambios globales de los suelos ferralíticos rojos lixiviados (Nitisoles Ródicos Eútricos) de la provincia La Habana. Cultivos Tropicales, 2006, vol. 27, no. 2, pp. 41-50.
9. IIHLD. Recomendaciones técnicas para el cultivo del maíz. 1997, 27p.
10. Castellanos, Marcela., Segura, Mariela. y Ñústez, C. E. Growth Analysis and Source-Sink Relationship of Four Potato Cultivars (Solanum tuberosum L.) in the Zipaquira Town (Cundinamarca, Colombia). Rev.Fac.Nal.Agr. Medellín, 2010, vol. 63, no. 1, pp. 5253-5266.
11. Bergamaschi, H. / et al. /. Distribuicao hidrica no periodo critico do milho e producao de graos. Pesq. Agropec. Bras., 2004, vol. 39, no. 9, pp. 831-839.
12. Cirilo, A. Fecha de siembra y rendimiento en maíz. INTA. Pergaminos. Buenos Aires. [Consultado: nov.2010]. Disponible en: http://www.inta.gov.ar/ ediciones/idia/cereales/maiz07.pdf
13. Soto, F., Plana, R. y Hernández, N. Influencia de la temperatura en la duración de las fases fenológicas del trigo harinero (triticum aestivum ssp. Aestivum) y triticale (x triticum Secale wittmack) y su relación con el rendimiento. Cultivos Tropicales, 2009, vol. 30, no. 1, pp. 57-60.
14. Maqueira, L. A., Torres, W. y Miranda, A. Crecimiento y rendimiento de dos variedades de arroz de ciclo corto en época poco lluviosa. Cultivos Tropicales, 2009, vol. 30, no. 3, pp. 28-31.
15. Shipley, P. Net assimilation rate, specific leaf area and leaf mass ratio: which is most closely correlated with relative growth rate? A meta-analysis. Fun. Ecol., 2006, no. 20, pp. 565-574.
16. Carranza, C., Lanchero, O., Miranda, D. y Chaves, B. Growth analysis of ‘Batavia’ lettuce (Lactuca sativa L.) cultivated in a saline soil of the Bogota Plateau. Agronomía Colombiana, 2009, vol. 27, no. 1, pp. 41-48.
17. Mora, R. / et al. /. Índices de eficiencia de genotipos de papa establecidos en Condiciones de secano. Revista Chapingo, 2006, vol. 12, no. 001, pp. 85-94.
18. Cabrera, A., Terán, Z., Soto, F. y Bertolí, M. Época de plantación de la Jamaica en Cuba. En: (Gómez, J.F.; Coordinador General): 2009. Avances científicos y tecnológicos de la cadena productiva del cultivo de la Jamaica. Primera edición. México 224 pp.
19. Jarma, A., Rengifo, Teresita. and Aramendiz-tatis, H. Physiology of stevia (Stevia rebaudiana) regarding radiation near the Colombian Caribbean coast.: II. Growth analysis. Agron. colomb., 2006, vol. 24, no. 1, pp. 38-47.
20. Páez, A., Paz, V. y López, J. Crecimiento y respuestas fisiológicas de plantas de tomate cv. 'Río Grande'. Rev. Fac. Agron. (LUZ), 2000, vol. 17, no. 2, pp. 173-184.
21. Lindquist, J., Arkebauer, T., Walters, D., Cassman, K. y Dobermann, A. Maize radiation use efficiency under optimal growth conditions. Agron. J., 2005, vol. 97, pp. 72-78.
22. De Grazia, J., Tittonell, P. y Chiesa, A. Efecto de la época de siembra, radiación y nutrición nitrogenada sobre el patrón de crecimiento y el rendimiento del cultivo de lechuga (Lactuca sativa L.). Invest. Agr. Prod. Prot. Veg, 2001, vol. 16, no. 3, pp. 356-365.
23. Páez, A., González, M. y Villasmil, J. Acclimation of Panicum maximum to different light regimes: effect of subsequent defoliation. Rev. Fac. Agron. (LUZ), 1997, vol. 14, no. 6, pp. 625-639.
24. Wardlaw, I. F. The control of carbon partitioning in plants. New Phytologist, 1990, vol. 116, no. 3, pp. 341-381.
25. Santos, Marcela., Segura, Mariela. and Ñústez, C. E. Growth Analysis and Source-Sink Relationship of Four Potato Cultivars (Solanum tuberosum L.) in the Zipaquira Town (Cundinamarca, Colombia). Rev.Fac.Nal.Agr.Medellín, 2010, vol. 63, no. 1, pp. 5253-5266.
26. Flores, R. / et al. /. Effect of solar radiation in the production of seed potato in soiless cultivation. Revista Chapingo Serie Horticultura, 2009, vol. 15, no. 1, pp. 25-30.
Recibido 21/07/2010, aceptado 6/09/2011.
Naivy Hernández, Reserva Científica del Instituto nacional de CienciasAgrícolas, gaveta postal 1, San José de las Lajas, Mayabeque, Cuba,CP 32 700. Email: naivy@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}