










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Protección Vegetal
On-line version ISSN 2224-4697
Rev. Protección Veg. vol.31 no.3 La Habana Sept.Dec. 2016
ARTÍCULO ORIGINAL
Distribución espacial y temporal de Empoasca spp. (Typhlocybinae) en un campo de frijol (Phaseolus vulgaris L.)
Spatial and temporal distribution of Empoasca spp. (Typhlocybinae) in a bean field (Phaseolus vulgaris L.)
Adayakni Sánchez-Castro, Ileana Miranda Cabrera, Basilia Miriam Fernández Argudín
Dirección de Sanidad Vegetal, Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Provincia Mayabeque, Cuba
RESUMEN
Fue objetivo del presente trabajo determinar la distribución espacio–temporal de Empoasca spp. y su relación con las variables meteorológicas de interés en un campo de producción de frijol de 26 ha (variedad Cuba Cueto 25-11), en Güines, Mayabeque, Cuba. Para ello se muestrearon, cada siete días, 90 plantas en el periodo enero a marzo de 2015. Se calculó la distribución espacial a partir de la Ley de Potencia de Taylor. Se determinaron el número de muestras y las curvas poblacionales de ninfas y adultos mediante el registro de la densidad poblacional por muestreo. Para determinar la influencia de las variables meteorológicas registradas sobre la población, se llevó a cabo un análisis de componentes principales. La distribución espacial de la población de los saltahojas tendió a la agregación. Las curvas poblacionales de ninfas y total mantuvieron un comportamiento similar, con un pico poblacional en el segundo muestreo. El número de plantas a muestrear fue de 20 plantas y no de 30, lo cual reduce el costo del muestreo. Se comprobó que la humedad, la temperatura y la velocidad del viento fueron las variables con mayor repercusión en el comportamiento de la población.
Palabras clave: curva poblacional, distribución espacial, Empoasca spp., Phaseolus vulgaris.
ABSTRACT
The objective of the present work was the determination of the spatial and temporal pattern distribution of Empoasca spp. and its relationship with weather variables of interest in a 26 ha black bean field (variety Cuba Cueto 25-11) in the municipality of Güines, Mayabeque, Cuba. In the first trimester of 2015, 90 plants were sampled each seven days. The spatial pattern distribution was calculated using Taylor´s power law (LPT): s2 = amb. The population curves were determined from the population density per sampling, and a principal component analysis was used to determine the relationship of the populations with the climate factor. The spatial distribution of the leafhopper population tended to the aggregation, and the nymph and adult population curves had a similar behaviour with a population peak in the second sampling. The sampling of twenty plants instead of thirty was determined to be enough. Humidity, temperature, and wind velocity were the variables with the greatest impact on the behavior of the leafhopper population.
Key words: population curve, spatial distribution, Empoasca spp., Phaseolus vulgaris.
INTRODUCCIÓN Entre las leguminosas, el frijol común (Phaseolus vulgaris L.) es una de las de mayor consumo en el mundo (1) por ser fuente de proteína vegetal en la dieta humana (2). En Cuba es un alimento de gran demanda (1), pero no existen niveles de producción que cubran las necesidades de la población, por lo que resulta de notable interés estudiar los fitófagos que habitan el cultivo, muchos de los cuales pueden causar daño.
Entre los insectos que inciden en las plantas del grano se encuentran los saltahojas, que son capaces de producir severas lesiones (3) a través de su alimentación directa (4, 5) y por ser vectores de enfermedades (6, 7, 8). En un estudio precedente, se determinó que los saltahojas recolectados e identificados en el frijol en zonas de Mayabaque, Cuba, conformaron un complejo de especies del género Empoasca (Typhlocybinae: Empoascini) identificados como Empoasca kraemeri Ross y Moore, Empoasca fabae Harris y Empoasca papayae Oman (9). Estos insectos son difíciles de separar por especies para determinar su comportamiento poblacional, debido a su pequeño tamaño y porque presentan características morfológicas externas afines entre sí (10). Sin embargo, coexisten en las plantas con un comportamiento similar, por lo que para determinar sus parámetros poblacionales se debe considerar el complejo Empoasca spp.
Los sistemas de vida de las poblaciones tienen un entramado complejo que requieren del conocimiento de los atributos esenciales que les son inherentes. Estos atributos, como son la disposición que presentan en el espacio, las fluctuaciones poblacionales en el tiempo y su relación con las variables meteorológicas, entre otras, son herramientas que permiten establecer medidas precisas para la elaboración de programas de control y manejo (11).
El conocimiento de cómo una población se distribuye en el espacio representa una primera aproximación hacia el logro de reducir el número de unidades muestreales, con miras a minimizar el costo y al acercamiento del plan de muestreo que represente la población (12, 13, 14). Además, permite definir la diversidad de las especies en las regiones y subregiones geográficas de las localidades en estudio (15).
La teoría de la dinámica de poblaciones utiliza una serie de modelos matemáticos para describir el comportamiento de las poblaciones en el tiempo; el modelo más simple describe el crecimiento poblacional como una función exponencial, dependiente de una población inicial y de una tasa de crecimiento (11, 16). La dinámica poblacional de los insectos está gobernada por un complejo proceso de interacciones exógenas (denso-independientes) y endógenas (denso-dependientes), las cuales repercuten en la regulación poblacional (17, 18).
Si bien estos instrumentos se emplean en el análisis poblacional de la especies fitófagas mundialmente, los estudios de dinámica poblacional realizados en Cuba (19, 20, 21) se apartan un poco de estas útiles herramientas, que también sirven de base para definir el patrón de dispersión de la enfermedad que transmiten (22).
A partir de estas premisas el presente trabajo tuvo como objetivos determinar la distribución espacio-temporal que exhibe el complejo Empoasca spp. y conocer la influencia que ejercen las variables meteorológicas (temperatura, humedad, lluvia y velocidad del viento) en su comportamiento poblacional en un campo de frijol.
MATERIALES Y MÉTODOS
Se realizaron muestreos, con frecuencia de siete días, en un campo de frijol de la variedad Cuba Cueto 25-11 de 26 ha, con marco de plantación de 7 cm x 0,90 cm, en la finca de un productor privado, en el periodo enero a marzo de 2015, en Güines, Mayabeque, Cuba.
Se contabilizaron los ejemplares de Empoasca spp. en 30 plantas en cada parcela, de las tres en que se dividió el campo para el área de muestreo (N =90). La densidad poblacional se estimó como la media del número de insectos (ninfas, adultos y total) por planta para conocer la tendencia de la distribución espacial y confeccionar la curva poblacional por estructura de edades (ninfas y adultos) y población total que refleja las fluctuaciones de la población en el tiempo.
Con las medias de población (m) generadas en cada evaluación y sus respectivas varianzas (s2), transformadas a logaritmo, se elaboró un análisis de regresión lineal y se ajustó a la función de la Ley de Poder de Taylor (LPT): s2 = a mb, equivalente a la función log s2 = log a + b log m; donde a es un factor, dependiente del tamaño de la unidad de muestreo, mientras que b (pendiente) es el coeficiente del tipo de distribución (23).
Se calculó el número de muestra mediante la fórmula: n =E-2amb-2 (E-error) y se usó la ecuación m = antilog [log (n.E2.a-1) (b-2)-1] para estimar el valor de las medias; donde a y b fueron, respectivamente, el intercepto y el coeficiente de agregación derivados de la LPT (24, 25).
Se realizó un análisis de componentes principales con los datos registrados de las temperaturas máxima, media y mínima; de las humedades relativas máxima, media y mínima; lluvias, velocidad del viento y la población. Se empleó el programa informático InfoStat 2.0 en el análisis estadístico (26).
RESULTADOS Y DISCUSIÓN
Distribución espacial de Empoasca spp.
La distribución espacial de la población de Empoasca spp. tendió a la agregación con valores de: a ═ 0,27; b ═ 1,58; R2═ 0,84. Este tipo de distribución sugiere que para el monitoreo debe realizarse un diseño experimental del tipo aleatorio estratificado sistemático.
Estos resultados conciden con lo informado por Segnini (27) quien, en sus estudios desarrollados en Venezuela, encontró que E. kraemeri, si bien puede presentar variaciones en la distribución espacial en relación con la época del cultivo y en la fase de desarrollo del insecto, los adultos, los huevos y las ninfas presentaron una distribución agregada durante el mayor periodo de estudio en el frijol. Las variaciones en el tipo de distribución resultan del cambio temporal en la densidad donde se dan elementos de ganancia (natalidad e inmigración) o pérdida (mortalidad y emigración) (28).
En Irán, Naseri et al. (29) informaron una distribución espacial agregada de Empoasca dicipiens (Paoli) en frijol, mientras que en otras especies de leguminosas indicaron una distribución aleatoria a causa de la densidad poblacional en las distintas especies. Estos autores hicieron referencia a que E. kraemeri puede tener distribución agregada, o al azar, en dependencia del hábitat. Kogan y Herzog (30), por su parte, encontraron que las ninfas de E. fabae se distribuyeron al azar en dependencia de la pubescencia de las hojas de la soya. Como puede observarse, el tipo de distribución, aunque es una característica específica para la especie, puede modificarse en dependencia de las condiciones del ambiente donde se desarrollen y de su potencial biótico. De ahí la necesidad de conocer la tendencia de la dispersión cuando se trate de estudios poblacionales, porque a través de ese conocimiento se deben encaminar los estudios sobre la dinámica poblacional.
Número de muestra
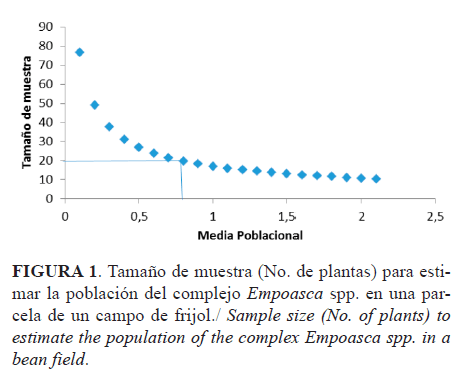
A partir de los datos obtenidos en la distribución espacial, se arribó a la conclusión de que 20 plantas eran suficientes para hacer los monitoreos y no 30 (Figura 1); lo que evidencia que se logra el mismo objetivo con menor número de plantas y así se reduce el costo del muestreo, por concepto de tiempo.
Distribución temporal de Empoasca spp.
Las curvas poblacionales de ninfas, adultos y total (ninfas + adultos) describieron similar comportamiento (Figura 2). Cuando se implanta el cultivo hay un flujo migratorio de adultos hacia las plantas, fundamentalmente de hembras (27), que implica un crecimiento poblacional alto; esto justifica el pico poblacional de ninfas y adultos en el segundo muestreo. A partir de ahí se produjo una desaceleración, ya que los adultos migran durante el desarrollo del cultivo hacia hojas y plantas sanas para garantizar las siguientes generaciones (10). A partir del tercer muestreo se observó, además, una tendencia al crecimiento, pero este fue menor, paulatino y lento, y se observó el envejecimiento del cultivo.
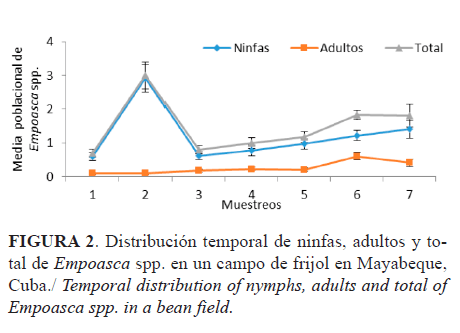
Otro elemento a tener en cuenta para explicar la tendencia al crecimiento desde el inicio se debe al corto ciclo de desarrollo de las especies involucradas (31). Es de significar que el efecto de grupo (competencia por el espacio) ejercido por la alta densidad de la población, a causa del crecimiento explosivo desde el segundo muestreo, pudo provocar una caída significativa que, unida a los otros factores mencionados, estuvieron incidiendo, como son la edad del cultivo y la emigración de los adultos.
Murguido (19), en experimentos realizados en la región occidental de Cuba, observó que la incidencia de E. kraemeri sobre plantas de frijol comenzó desde la fase fenológica de hoja primaria (V1); señaló, además, que los datos de los conteos demostraron que una vez establecidos los primeros ejemplares, la población aumentó progresivamente (periodo de crecimiento abierto) hasta alcanzar el valor máximo (vértice o pico de la población) y en ese momento el número de saltahojas disminuyó, lo que concuerda con los resultados observados en este trabajo.
Relación de las variables meteorológicas con la densidad poblacional de Empoasca spp.
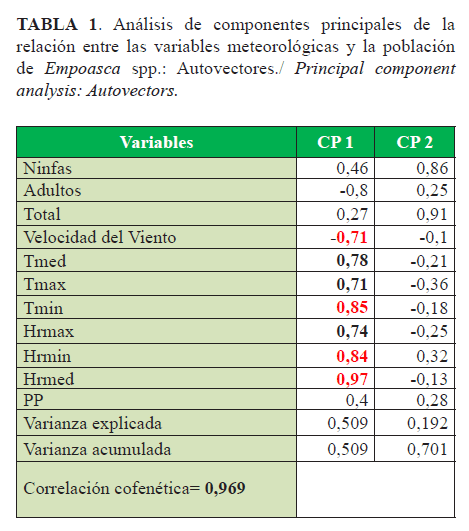
Los resultados alcanzados evidenciaron que con las dos primeras componentes es posible explicar más del 70% de la variación total con alta correlación. Como se muestra, la humedad media y mínima tuvieron mayor ponderación positiva sobre los adultos y las ninfas, seguido de las temperaturas, fundamentalmente la mínima (Tabla 1, Figura 3).
La velocidad del viento, que también tuvo peso entre las variables, influyó sobre los adultos, de donde puede inferirse que favoreció el desplazamiento a las plantas cercanas para iniciar una nueva generación.
La lluvia mostró poca influencia (Figura 3); algunos autores (32, 33) refirieron que las abundantes lluvias disminuyen sus poblaciones, pero rápidamente se recuperan. Estudios similares revelan que el ataque de E. kraemeri es más severo en épocas secas y cálidas y la situación se agrava cuando la humedad del suelo es insuficiente (27).
Los saltahojas se caracterizan por presentar unas cubiertas denominadas brocosomas, estructuras superhidrofóbicas que repelan el agua y sus propios exudados (34). Este componente, producido a partir de sus túbulos de Malpighi, los protege del agua, por lo que al margen de que puedan ser barridos por excesivas lluvias gran parte queda protegido y puede recuperar rápidamente sus poblaciones. Otro elemento de importancia es que su nicho fundamental es el envés, que les proporciona protección.
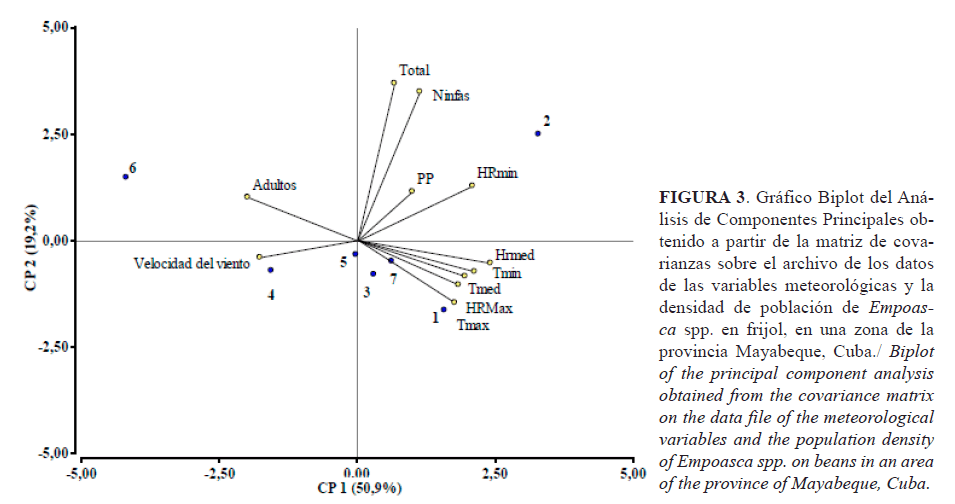
El gráfico de dispersión Biplot (Figura 3) muestra lo explicado anteriormente. Los puntos azules reflejan las fechas de los muestreos realizados. Se observa que en los muestreos 1, 2, 3 y 7 las temperaturas y las humedades tuvieron mayor repercusión; en los muestreos 4, 5 y 6, la velocidad del viento tuvo mayor responsabilidad en la variabilidad.
Estos estudios constituyen los primeros que se desarrollan en Cuba con Empoasca spp. Si bien se han abordado los estudios de las fluctuaciones en el tiempo, donde se consideraron aspectos como la fenología y las condiciones del tiempo, han adolecido de un elemento esencial, como es el conocimiento del tipo de distribución que tienen en el espacio: elemento clave para conocer la dinámica temporal del grupo. Estos elementos permiten, además, precisar los momentos óptimos para aplicar alguna medida de control. No obstante, resulta conveniente continuar los estudios de dinámica de población de saltahojas-enfermedades para ampliar estos resultados.
REFERENCIAS
1. Talukder Z, Anderson E, Miklas P, Blair M, Osorno J, et al. Genetic diversity and selection of genotypes to enhance Zn and Fe content in common bean. Can J Plant Sci. 2010;90:49-60.
2. Batista K, Prudencio S, Fernandes K. Changes in the functional properties and antinutritional factors of extruded Hard-to-Cook common beans (Phaseolus vulgaris, L.). Journal of Food Science. 2010;75:286-290.
3. Sardinha de Souza B, Gabriel de Oliveira J, Oliveira T, Ribeiro Campos A, Boiça A Jr. Infestação de Empoasca kraemeri Ross & Moore e Caliothrips phaseoli (Hood) em cultivares de feijoeiro, na época da seca, em Selvíria-MS, e os efeitos na produtividade. Entomo Brasilis. 2012;5(3):204-210.
4. Baje L, Stewart AJA, Novotny V. Mesophyll cell-sucking herbivores (Cicadellidae: Typhlocybinae) on rainforest trees in New Guinea: local and regional diversity of a taxonomically unexplored guild. Ecological Entomology. 2014;39(3): 325-333. ISSN 0307-6946.
5. Backus EA, Serrano MS, Ranger CM. Mechanisms of hopperburn: an overview of insect taxonomy, behavior, and physiology. Annu Rev Entomol. 2005;50:125-151.
6. Acosta K, Piñol B, Arocha Y, Wilson M, Boa E, Lucas J. Transmission of Phytoplasma associated with Bunchy Top symptom of papaya by Empoasca papayae Oman. J Phytopathol. 2010;158:194-195.
7. Bosco D, D´Amelio R. Transmission specificity and competition of multiple phytoplasmas in the insect vector. In: Weintraub PG, Jones P (Eds) Phytoplasmas: genomes, plant hosts and vectors. CAB International, Wallingford, UK. 2013; 293-308.
8. Ishii Y, Matsuura Y, Kakizawa Sh, Nikoh N, Fukatsua T. Diversity of Bacterial Endosymbionts associated with Macrosteles leafhoppers vectoring phytopathogenic phytoplasmas. Applied and Environmental Microbiology. 2013;79(16):5013-5022.
9. Sánchez A, Miranda I, Quiñones M, Piñol B, Fernández M. Saltahojas (Typhlocybinae) y su relación con los síntomas de enfermedades en un campo de frijol (Phaseolus vulgaris L.). Rev Protección Veg. 2016; 31(3):153-158.
10. Sánchez A, Quiñones M, Piñol Pérez BE, Fernández BM. Primer informe de Typhlocybinae como vectores potenciales de fitoplasmas en Cnidoscolus chayamansa (Miller) I.M. Johnst. (chaya) en Cuba. Rev Protección Veg. 2015;30(2):148-157.
11. Sharov A. Quantitative population ecology. Department on Entomology. Virgina Tech, Blacksburg, VA. 1996; 119 p.
12. Vivas L, Notz A. Plan de muestreo secuencial de Oebalus insularis Stal (Hemiptera: Pentatomidae) en el cultivo de arroz en Calabozo estado Guárico, Venezuela. Revista Científica UDO Agrícola, 2009;9(4):857-872.
13. Vivas LE, Notz A. Distribución espacial en poblaciones de Oebalus insularis Stal (Hemiptera: Pentatomidae) en el cultivo de arroz en Calabozo, estado Guárico, Venezuela. Revista Científica UDO Agrícola. 2011;11(1):109-125.
14. Vecco D, Pinedo R, Fernández M. Métodos de muestreo para Pseudophilothrips sp. (Tubulifera: Phlaeothripidae) y sus enemigos naturales: I. Muestreo secuencial enumerativo. Rev Protección Veg. 2015;30(1): 6-13.
15. Wang X, Huang M, Zhang Y. Cluster analysis of Typhlocybinae (Hemiptera: Cicadellidae) distributional pattern in China. AGRIS. 2013;40(4):86-94.
16. Abramson G. La matemática de los sistemas biológicos. Instituto Balseiro/Universidad Nacional de Cuyo, Argentina. 2013; 279 p. url http//física.cab.enea.gov.ar/estadistica/abramson/notes/biológicos.pdf.
17. Wilby A, Thomas MB. Diversidad y manejo en los ecosistemas. EnJarvis DI; Padoch C; Cooper HD (eds). El manejo de la Biodiversidad en los ecosistemas agrícolas. Roma. 2011; 283-307 pp.
18. Paleólogos MF, Flores CC. Principios para el manejo ecológico de plagas. EnSarandón SJ; Flores CC (eds). Agroecología, bases teóricas para el diseño y manejo de agroecosistemas sustentables. 1ª. ed. Universidad Nacional de La Plata, Argentina. c.10. 2014; 260-285 pp.
19. Murguido CA. Biología, Ecología y lucha contra el saltahojas Empoasca kraemeri Ross y Moore (Homoptera: Cicadellidae) en el frijol (Phaseolus vulgaris L.). [Tesis presentada en opción del grado científico de Doctor en Ciencias Agrícolas]. Instituto de Investigaciones de Sanidad Vegetal. Ciudad de La Habana. Cuba. 1995; 98 p.
20. Pérez L, Borges M, Luis M. Aspectos sobre biología, ecología y manejo de Empoasca papayae Oman (Homoptera: Cicadellidae). Boletín Informativo NotiCitriFrut. 2008;10:1-4.
21. Hidalgo-Gato M, Rodríguez R, Ricardo N, Hermen E. Dinámica poblacional de cicadélidos (Homoptera: Cicadellidae) en un agroecosistema cañero de Cuba. Rev Biol Trop. 1999;47(3):2511-5550.
22. Orenstein S, Zahavi T, Nestel D, Sharon R, Barkalifa M, Weintraub P. Spatial dispersion patterns of potential leafhopper and planthopper (Homoptera) vectors of phytoplasma in wine vineyards. Ann Appl Biol. 2003;142:341-348.
23. Taylor LR. Aggregation, variance and the mean. Nature. 1961;189:732-735.
24. Vivas Carmona LE, Notz A. Número de muestras requeridas para estimar la población de Oebalus insularis Stal en el cultivo de arroz (Oriza sativa L.) en Calabozo, estado Guárico, Venezuela. Revista Científica UDO 56 Agrícola. 2013;13(1):56-65.
25. Vivas LE, Notz A. Distribución espacial en poblaciones de Oebalus insularis Stal (Hemiptera: Pentatomidae) en el cultivo de arroz en Calabozo, estado Guárico, Venezuela. Revista Científica UDO Agrícola 2011;11(1):109-125.
26. Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW. InfoStat versión 2016. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Disponible en: http://www.infostat.com.ar.
27. Segnini S, Montagne A. Biologia y ecología poblacional de Empoasca kraemeri Ross y Moore (Homoptera: Cicadellidae) en caraota (Phaseolus vulgaris). Fluctuación poblacional de E. kraemeri en campos cultivados con caraota. Agronomía Tropical. 1986;36(4-6):29-45.
28. Huffaker CB, Van der Vries M, McMurtry JA. Ecology and natural enemies. Hilgardia. 1999;27(14) 343-383.
29. Naseri B, Fathipour Y, Talebi A. Population Density and Spatial Distribution pattern of Empoasca decipiens (Hemiptera: Cicadellidae) on different bean species. J Agric Sci Technol. 2009;11:239-248.
30. Kogan M, Herzog DC. Sampling Methods in Soybean Entomology. Springer Verlag, New York, 1980; 587 pp.
31. Maya-Hernández V, Vera-Graziano J, Garza-García R. Parámetros poblacionales de Empoasca kraemeri Ross & Moore (Homoptera: Cicadellidae) en genotipos de frijol. Agrociencia. 2000;34:603-610.
32. Edelstein JD, Bartó CA, Trumper EV. ARTROPOB: Sistema de Simulación de Dinámica Poblacional y Manejo de Artrópodos Plaga. 39JAIIO - ISSN: 1852-4850 - CAI 2010; 763p.
33. Carvalho DP, Oliveira HN. Population fluctuation of Empoasca sp. (Hemiptera: Cicadellidae) in a physic nut crop in MatoGrosso do Sul. Acta Biol Colomb. 2016;21(1):81-85.
34. Rakitov R, Gorb SN. Brochosomal coats turn leafhopper (Insecta, Hemiptera, Cicadellidae) integument to superhydrophobic state. Proc R Soc B 2013;280:2012-2391. http://dx.doi.org/10.1098/rspb.2012.2391.
Recibido: 2/03/16
Aceptado: 18/07/2016
Basilia Miriam Fernández Argudín, Dirección de Sanidad Vegetal, Centro Nacional de Sanidad Agropecuaria (CENSA). Email: miriam@censa.edu.cu


{kind=link}
{kind=link}