










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkVaccimonitor
On-line version ISSN 1025-0298
Vaccimonitor vol.18 no.2 Ciudad de la Habana May-Aug. 2009
ARTICULOS ORIGINALES
Nueva generación de vacunas antimeningococica basadas en vesículas de membrana externa.
New generation of outer-membrane vesicles-based meningococcal vaccines.
Jan Poolman, Philippe Denoël, Nathalie Devos, Christiane Feron, Karine Goraj, Patricia Momin, Christine Tans, Vincent Weynants
GlaxoSmithKline Biologicals, Rixensart, B-1330, Belgium.
email: gmoe@chori.org
Abstract
With the development of conjugate vaccines, Neisseria meningitidis serogroup B (MenB) will remain the major cause of meningitis, as it is not covered by vaccination. In order to develop a vaccine able to confer a broad cross-protection against invasive MenB strains, we have developed a new generation of outer-membrane vesicles (OMVs) based on the over-expression of well conserved minor outer membrane proteins (OMPs) and the presence of high percentage of detoxified LOS.
Keywords: Neisseria meningitidis,OMV, meningococcal vaccines
Introduction
With the availability of H. influenzae type B vaccine, meningococcal serogroup A, C, W-135 and Y vaccines, and pneumococcal conjugate vaccine, serogroup B meningococci will remain the major cause of bacterial meningitis. The use of serogroup B capsular polysaccharide as vaccine antigen has been hampered by its poor immunogenicity and by potential concern of inducing auto-antibodies.
Our strategy to develop a safe, immunogenic and cross-protective MenB vaccine is based on the selection of different surface antigens, well conserved, and involved in different stages of the infection process. In order to present these vaccine antigens in their natural environment, an alternative expression system was developed in N. meningitidis, taking into account its capacity to produce large amounts of outer-membrane vesicles (OMVs). The OMVs are enriched in vaccine antigens because they are produced from strains genetically modified to over-express the selected antigens by using either the gene delivery (GD) or promoter replacement (PR) strategies (1).
After literature survey and genome mining, approximately 50 potential antigens were selected for subsequent evaluation. In addition to these proteins, the lipo-oligosaccharide (LOS) of invasive MenB strains was also evaluated as vaccine antigen because antibodies to LOS are known to be bactericidal.
In addition, due to its adhesion and toxic properties, the LOS can be considered as a major virulence factor. Although at least 11 different LOS immunotypes are described, L3,7 represents up to 70% of disease cases while L2 appears to be the second most prevalent immunotype (2).
By using a preclinical mouse model that mimics the PorA-specific bactericidal response observed in infants with wild-type OMVs, we have developed a new generation of OMV- based meningococcal vaccines (3). Some key results are described to illustrate this development.
Synergy of antibodies against minor MenB OMPs
Because of the dominant immune response mediated by PorA, OMVs derived from MenB wild-type strains induce only limited cross-protection against circulating heterologous strains. This partial cross-protection appears to be induced by minor and well-conserved outer-membrane proteins (OMPs) and LOS. We have genetically modified strain H44/76 to prevent the expression of the major variable protein PorA and to up-regulate the expression of some minor OMPs that have potential as vaccine antigens. These over-expressions were done in a galE LOS background and OMVs were prepared by detergent extraction using 0.5% Na deoxycholate (DOC).
The bactericidal response measured in sera from mice immunized with OMVs over-expressing only one minor OMP showed no or only limited increase in bactericidal antibodies even though the induction of specific anti-OMP was demonstrated by ELISA. However, by mixing the sera of animals immunized with OMVs over-expressing TbpA and OMVs over-expressing Hsf, we observed a positive interaction between anti-TbpA and anti-Hsf antibodies in the ability to induce complement-mediated killing of bacterial strains. This finding was confirmed by the immunization of mice with OMVs co-over-expressing both TbpA and Hsf (Figure 1). Comparable synergistic effects were shown between other minor OMPs, such as Nspa and OMP85.
Anti-LOS antibodies for protection against N. meningitidis strains
Since LOS is the main target of bactericidal antibodies in convalescent subjects (4), we have evaluated the use of D porA OMVs as a vehicle to present LOS. In a first series of preclinical experiments, OMVs expressing a full L3,7 LOS or a galE LOS were tested for their potential to induce a bactericidal antibody response. The DOC extraction process to prepare OMVs was evaluated by comparison of 0.1% DOC (leaving high LOS levels within OMV) and 0.5% DOC (classical OMV process). It was observed that only OMVs containing a high percentage of L3,7 LOS were able to mediate a protective response against most of the invasive L3,7 MenB strains tested.
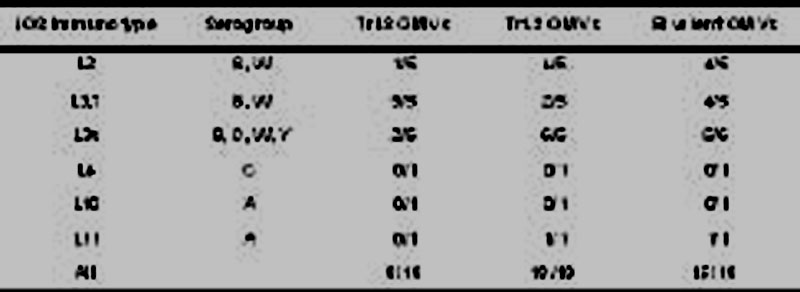
Two genetic modifications were used to alter LOS: one modification (lgtB mutation) removes the terminal galactose of LOS (TrL3), the other modification leads to a penta-acylated, less toxic lipidA (msbB mutation). The 0.1% DOC OMVs produced from this modified H44/76 strain (TrL3 OMVs) were still able to induce cross-protective bactericidal responses against L3,7 MenB strains and demonstrated a reduced and acceptable reactogenicity profile in different LOS toxicity models (pyrogenicity in rabbit, production of pro-inflammatory cytokines by human monocytic cells, degree of inflammation at the injection site in mice and rabbits). However, 0.1% DOC OMVs based on the L3,7 immunotype were not able to induce a bactericidal response against strains expressing either the L2 immunotype or the recently described L3v immunotype, both differing from L3,7 by the composition of the LOS inner core. Therefore, a bivalent OMV vaccine based on detoxified L3,7 and L2 LOS (lgtB and msbB mutations) was evaluated in mice. This bivalent vaccine induced the production of bactericidal antibodies able to mediate complement killing of most of the meningococcal strains tested, irrespective of the serogroup (Table 1).
Discussion
Over-expression of minor OMPs in MenB vaccine strains followed by purification of OMVs resulted in antigen preparations that have demonstrated their potential against Men B, by raising specific anti-OMP serum antibodies in mice. This effect was due to the correct presentation (native folding) of the proteins, as opposed to recombinant integral OMPs that are known as weak inducers of specific bactericidal antibodies. However, to efficiently kill wild-type bacteria, it was also observed that several minor OMPs need to be targeted in order to reach the threshold level of bactericidal antibodies for the activation of the classical complement pathway. We further provided evidence for LOS being a potent vaccine antigen, and again, a threshold level of LOS in OMVs had to be reached to induce anti-LOS antibodies in mice.
Figura 1 . Over-expression of Hsf and/or TbpA. (A) Impact on the protein content in 0.5% DOC OMVs. (B) Impact on the induction of antibodies against minor OMPs in mice immunized with purified proteins, and impact on the induction of complement-mediated killing by bactericidal antibodies (adapted from [3]).
Tabla 1. Induction of bactericidal antibodies in mice immunized with either monovalent or bivalent OMV vaccine. Serum antibodies were tested in rabbit SBA against a panel of 19 N. meningitidis strains. Results are expressed as the number of strains killed (titre ³ 1/128) per number of strains tested (adapted from [5]).
Therefore, the use of detoxified mutated LOS was needed to combine a high level of LOS and an acceptable safety/toxicity profile.
References
1. Poolman JT, Feron C, Desquesne G, et al. Outer membrane vesicles and other options for a meningococcal B vaccine. In: Ferreirós C, Criado MT, Vázquez J, editors. Emerging strategies in the fight against meningitis: molecular and cellular aspects.Wymondham, Norfolk, UK, Horizon Scientific Press, 2002: p. 135-49.
2. Scholten RJ, Kuipers B, Valkenburg HA, Dankert J, Zollinger WD, Poolman JT. Lipo-oligosaccharide immunotyping of Neisseria meningitidis by a whole-cell ELISA with monoclonal antibodies. J Med Microbiol 1994;41:236-43.
3. Weynants VE, Feron CM, Goraj KK, et al. Additive and synergistic bactericidal activity of antibodies directed against minor outer membrane proteins of Neisseria meningitidis. Infect Immun 2007;75:5434-42.
4. Zollinger W, Moran E, Schmiel D, Brandt B. Specificity of cross-reactive bactericidal antibodies in normal and convalescent human sera (P8.3.19). 15th Int Pathogenic Neisseria Conference, September 10-15, 2006, Cairns, Australia 2006.
5. Weynants V, Denoël P, Devos N, Janssens D, Feron C, Goraj K, Momin P, Monnom D, Tans C, Vandercammen A, Wauters F, Poolman JT. Genetically modified L3,7 and L2 lipooligosaccharides from Neisseria meningitidis serogroup B confer broad cross-bactericidal response. Infect. Immun. 2009 (in press).


{kind=link}
{kind=link}