










 Custom services
Custom services Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las emisiones de gases de efecto invernadero (GEI) son una de las causas principales del calentamiento global de la Tierra en la actualidad y como producto de estas el cambio climático es una de las mayores amenazas sobre nuestro planeta. Estas emisiones proceden principalmente de la quema de combustibles fósiles tales como petróleo, gas y carbón (Ibárcena y Scheelje, 2003). Actualmente muchos proyectos están encaminados al estudio y desarrollo de diferentes alternativas novedosas para disminuir, eliminar o tratar dichas emisiones, con el fin de mitigar los efectos catastróficos que estas producen en nuestro planeta (Botello y col., 2005).
En este sentido, las algas marinas son un grupo grande y heterogéneo de organismos vegetales de unas 50 000 especies, entre las que se cuentan especies unicelulares (microalgas) y plantas que pueden medir hasta 5 metros (macroalgas) y se caracterizan por ser fotótrofas.
Las algas utilizan el CO2 como fuente de carbono (Chung et al., 2010). En ausencia de este compuesto el crecimiento se considera nulo, al igual que un suministro insuficiente a menudo es el factor limitante de la productividad. Según la composición química media de la biomasa de las algas, se necesitan 1,8 toneladas de CO2 para producir una tonelada de biomasa de microalgas (Vicente, 2010), (Rodríguez, 2012). La disolución natural de CO2 del aire en el agua no es suficiente. Se podría mejorar añadiendo aire en el agua, pero teniendo en cuenta que el aire contiene un 0,0383 % de CO2, se necesitaría todo el CO2 disponible en 37 000 m3 de aire para conseguir una tonelada de algas secas (Vicente, 2010), mientras que para la producción de 1t de macroalgas seca se necesitan 1,5 t de CO2 (Nikolaisen y col., 2011). Otra opción es el uso de CO2 puro, pero el costo de éste es elevado. Una alternativa novedosa tanto para el cultivo de las algas como para la disminución del efecto de las emisiones de los gases de combustión es utilizar una fuente de residuos de este gas, como pueden ser los gases de combustión de una caldera (Carlsson y col., 2007).
En Duarte y col., (2017) se reporta la acuicultura de algas marinas como un componente importante dentro de las estrategias de adaptación al cambio climático con una propuesta de cuatro parámetros socio-ecológicos a tener en cuenta: la productividad biológica para mejorar la captura de carbono (cuyos límites los determinan las características biológicas de las especies a cultivar), las limitaciones ambientales a la expansión de la acuicultura de algas marinas, los instrumentos normativos que permiten las diferentes prácticas de acuicultura, y la gestión de las preferencias de la sociedad. También afirman que el precio de mercado puede mantenerse para alentar a los cultivadores de algas marinas a aplicar mejoras de diseño que permitan aumentar los beneficios ambientales, pero esto requiere una diversificación de estos para aumentar la demanda de productos de algas marinas en el mercado.
Uno de los beneficios ambientales del cultivo de macroalgas es el consumo de gases de combustión, cuyo índice por hectárea varía según la especie de alga utilizada y a lo largo del día con la intensidad luminosa, por lo que ha de optimizarse para esta aplicación (Mooney-McAuley y col., 2016); (Zollmann y col., 2021).
En correspondencia con todo lo anterior, en Cuba se desarrollan proyectos e investigaciones para la disminución de la contaminación provocada por los gases de combustión usando propuestas novedosas, como el cultivo intensivo de algas, lo cual contribuye a mitigar los efectos negativos que producen estos gases y genera biomasa algal, un producto valioso que contribuye a la sostenibilidad del proceso.
Considerando lo anteriormente expuesto, el objetivo de este trabajo es estimar el potencial fotosintético de micro- y macroalgas en reactores biológicos expuestos a la intemperie. Con este objetivo se pretende hacer más sostenible y con menor impacto ambiental la generación de energía eléctrica en el país. En este sentido, ya se dan pasos para aplicar esta investigación en la Empresa Termoeléctrica “Carlos Manual de Céspedes” de Cienfuegos. La novedad de la investigación es el tratamiento detallado de la propagación de la radiación fotosintéticamente activa y la ultravioleta (UV) dentro del biorreactor, acoplado a un modelo de fotosíntesis.
MATERIALES Y MÉTODOS
La metodología consta de los siguientes pasos:
Cálculo de las irradiancias espectrales incidentes en la superficie del biorreactor.
Cálculo de las irradiancias espectrales que atraviesan la interfase aire-agua.
Propagación del espectro obtenido en la columna de agua.
Cálculo del potencial fotosintético de las algas.
Para calcular las irradiancias espectrales E(λ,0+)-), (W/m2), que llegan a la superficie del biorreactor se propone utilizar el programa de propagación de radiaciones (Tropospheric Ultraviolet and Visible, 2020) (TUV), software libre en FORTRAN elaborado en el Centro Nacional de Investigaciones Atmosféricas de los Estados Unidos (https://www2.acom.ucar.edu/modeling/tropospheric-ultraviolet-and-visible-tuv-radiation-model). Se calculan las irradiancias espectrales para la banda de la radiación o luz fotosintéticamente activa (LFA) en el intervalo de longitudes de onda 400-700 nm. La radiación ultravioleta (UV) tiene un efecto inhibitorio sobre la fotosíntesis, por lo que se también se calculan las irradiancias espectrales en el intervalo 280-399 nm (por debajo de 280 nm la atmósfera bloquea casi totalmente el UV). Se propone modelar la variabilidad diaria y estacional de las irradiancias teniendo en cuenta diferentes ángulos solares zenitales y condiciones atmosféricas tales como la cobertura nubosa.
Una vez obtenido el espectro en la superficie acuática del biorreactor, es necesario propagarlo en la columna de agua. Se plantea modelar esto utilizando la clasificación óptica de las aguas costeras del oceanógrafo N. Jerlov (Shifrin, 1988), la cual provee los coeficientes de atenuación de la luz para los cinco tipos de aguas costeras, denotadas como C1, C3, C5, C7 y C9, acorde a su turbidez. En este trabajo se muestran resultados para las aguas más claras (C1) y las más turbias (C9), abarcando así el más amplio intervalo de posibles respuestas. Las irradiancias espectrales E(λ,0-), (W/m2), que penetran la interfase aire-agua en la superficie del biorreactor se calculan mediante la ecuación (1):
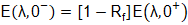 () 1
() 1
donde Rf (adimensional) es el coeficiente de reflexión obtenido de las fórmulas de Fresnel aplicadas a la interfase aire-agua (Fresnel_equations, 2021). Las irradiancias espectrales E(λ,z), (W/m2), a una profundidad z (m) dentro del biorreactor se obtienen a través de la ley de Lamber-Beer de la Óptica, dada por la ecuación (2):
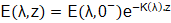 () 2
() 2
donde E(λ,0-) son las irradiancias espectrales justo debajo de la superficie acuática y K(λ), (m-1), son los coeficientes de atenuación de la luz. Para el caso de microalgas (fitoplancton), la ecuación (2) se aplica en toda la columna de agua. Para el caso de macroalgas una vez que la luz llega al tope de la canopia, su atenuación se incrementa debido al autosombreado. Existen varias maneras de modelar esta atenuación, que incluyen modelos geométricos de canopia y modificaciones de la ley de Lamber-Beer. En esta modelación preliminar se duplicaron los coeficientes de atenuación de la luz dentro de la canopia, puesto que la biomasa algal incrementa la atenuación de la luz, dejando para un trabajo futuro la comparación de estos resultados con los obtenidos por otros métodos (Zollmann y col., 2021).
Teniendo las irradiancias espectrales, se puede calcular el potencial fotosintético P(z), (mgC.m-3.h-1), a una profundidad z, normalizado respecto al máximo potencial posible Ps (mgC.m-3.h-1) mediante un modelo de fotosíntesis. Uno de los más usados es el llamado modelo E de fotosíntesis que muestra la ecuación (3):
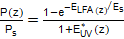 () 3
() 3
Es (W/m2) es el parámetro que mide la eficiencia de cada especie de alga en usar la radiación o luz fotosintéticamente activa (LFA): mientras más pequeño es este, mayor es la eficiencia de la especie para usar LFA.
En la anterior ecuación ELFA(z), (W/m2), es la irradiancia total de la radiación fotosintéticamente activa, calculada mediante la ecuación (4):
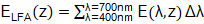 () 4
() 4
donde Δλ (nm) es la longitud del intervalo de longitudes de onda al que corresponde cada irradiancia espectral E(λ,z). La radiación ultravioleta está representada por E* UV(z), (adimensional), la cual se calcula por la ecuación (5):
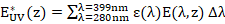 () 5
() 5
En esta ecuación el efecto inhibitorio que ejerce la radiación UV sobre la fotosíntesis está representado por el espectro de acción biológica ε(λ), (W-1.m2.nm-1).
El potencial fotosintético depende de la profundidad, por lo que para calcular su promedio <P/Ps>, (adimensional), en la dirección espacial vertical, se divide la columna de agua en N capas de igual espesor y se aplica la expresión (6):
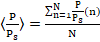 () 6
() 6
Para el caso de macroalgas sésiles solamente se considera la parte de la columna de agua en que realmente se hace fotosíntesis, o sea, la canopia algal. Los promedios obtenidos mediante la ecuación (6) son una medida de la productividad de biomasa del biorreactor.
RESULTADOS Y DISCUSIÓN
En este apartado se presentan resultados de la aplicación de la metodología propuesta para biorreactores con la macroalga Ulva lactuca. Se han diseñado biorreactores a escala de laboratorio en los que el agua tiene una profundidad de 40 cm, estimándose la altura promedio de la canopia algal de 20 cm (Roche, 2020).
Para calcular el potencial fotosintético primeramente se obtuvieron las irradiancias solares espectrales E(λ,0+) en la superficie acuática del biorreactor, utilizando el mencionado software de transferencia radiativa TUV. Como datos de entrada se utilizaron un ángulo solar zenital de 45 grados, una columna de ozono de 300 unidades Dobson, y densidades ópticas de la cobertura nubosa y de los aerosoles atmosféricos equivalentes a 0 y 0,235, respectivamente. Esto corresponde a condiciones de irradiación solar relativamente intensas, representativas de la mayor parte de los días en el archipiélago cubano. No obstante, la metodología propuesta es aplicable a otras condiciones de mayor o menor intensidad solar. Posteriormente se usaron las ecuaciones (1) y (2) para obtener las irradiancias E(λ,z) en la columna de agua y las ecuaciones (3) y (6) para determinar el potencial fotosintético a la profundidad z y el promedio en la canopia, respectivamente.
La Tabla 1 muestra el potencial fotosintético dentro de la canopia algal para las aguas costeras más claras (C1), mientras que la Tabla 2 lo hace para aguas costeras más turbias (C9). En ambos casos se observa un incremento del potencial a medida que aumenta la profundidad, lo cual se debe a que en columnas de agua someras (z<10 m) por lo general predomina el efecto inhibitorio de la radiación UV sobre el beneficioso de la LFA. Dentro de este intervalo de profundidades, a mayor profundidad mayor potencial fotosintético porque el efecto inhibitorio de la radiación UV es menor debido a su atenuación por el agua. Por la misma razón las aguas C9 muestran mayor potencial, debido a que bloquean más la radiación UV.
Aplicando la ecuación (6) se determinó que el potencial promedio dentro de la canopia, para aguas C1 es 56,5 % y para aguas C9 es 86,3 %. Se observa independencia del parámetro ES, el cual cuantifica la eficiencia en el uso de la LFA: otra consecuencia del predominio de la radiación UV sobre la LFA a las escasas profundidades del biorreactor. Por ende, las diferencias en productividad de biomasa para las distintas especies apuntan más a diferencias en la eficiencia en la asimilación de nutrientes (CO2 incluido) y a la tolerancia a la temperatura y pH. Esto concuerda con lo reportado en (Zollmann y col., 2021): la luz no siempre es el factor limitante en el cultivo de algas.
Tabla 1 Potencial fotosintético para las aguas costeras más claras (C1) y para las aguas costeras más turbias (C9)
| 0,2 | 43,4 | 43,4 | 0,2 | 75,4 | 75,4 |
| 0,22 | 51,6 | 51,6 | 0,22 | 78,2 | 78,2 |
| 0,24 | 52,9 | 52,9 | 0,24 | 80,9 | 80,9 |
| 0,26 | 54,3 | 54,3 | 0,26 | 83,2 | 83,2 |
| 0,28 | 55,6 | 55,6 | 0,28 | 85,3 | 85,3 |
| 0,3 | 56,9 | 56,9 | 0,3 | 87,2 | 87,2 |
| 0,32 | 58,2 | 58,2 | 0,32 | 88,9 | 88,9 |
| 0,34 | 59,5 | 59,5 | 0,34 | 90,3 | 90,3 |
| 0,36 | 60,8 | 60,8 | 0,36 | 91,6 | 91,6 |
| 0,38 | 62,0 | 62,0 | 0,38 | 92,7 | 92,7 |
| 0,4 | 63,2 | 63,2 | 0,4 | 93,7 | 93,7 |
CONCLUSIONES
La metodología propuesta permite estimar el potencial fotosintético de algas cultivadas intensivamente en biorreactores que utilizan dióxido de carbono residual industrial. Se presentaron resultados preliminares para biorreactores que serían alimentados con aguas marinas costeras, cubriendo todo el rango de turbidez de estas, desde las más claras hasta las más turbias. Para los casos analizados, los resultados muestran que en aguas costeras más turbias el potencial fotosintético es mayor que para aguas costeras más claras, lo cual está condicionado por el mayor poder inhibitorio de la radiación ultravioleta en aguas claras, a las pequeñas profundidades de los biorreactores. Asimismo, la eficiencia en el procesamiento de la radiación fotosintéticamente activa, característica de cada especie de alga, influye poco en el potencial fotosintético, por la dominancia de la radiación UV en aguas someras. Las investigaciones futuras se proyectan hacia la aplicación de un modelo más refinado de fotosíntesis que considere mejor las especificidades de la macroalga en cuestión, especialmente el aspecto del autosombreado.

