Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Los procesos auto-organizativos posteriores a la ocurrencia de perturbaciones en las comunidades vegetales implican cambios en las poblaciones y están fuertemente influenciados por los rasgos funcionales de las plantas (Gillison et al. 2013, Verheijen et al. 2015), por los tipos funcionales presentes (Ostertag et al., 2015, Salmerón-López et al. 2016a, Vásquez-Valderrama y Solorza-Bejarano, 2018) y por la naturaleza de las interacciones plantas-plantas (Salmerón-López y Geada-López 2018, Navarro-Cano et al. 2019).
Los rasgos funcionales influyen la presencia de las especies, la estructura etaria y los estados regenerativos del sistema en áreas sometidas a diferentes niveles de perturbaciones. Además, estos rasgos permiten definir los tipos funcionales de plantas, los que constituyen una valiosa herramienta para el diagnóstico de procesos asociados a las perturbaciones (Gillison et al. 2013), en el monitoreo a largo plazo de estrategias de manejo de ecosistemas y prácticas específicas de restauración (Kooyman y Rossetto 2008, Vásquez-Valderrama y Solorza-Bejarano, 2018).
El uso de la modelación con dinámica de sistemas es una vía de entender los posibles cambios en las comunidades a partir de los cambios en la composición, estructura, interacciones y funcionamiento, pero para esto, es necesario la incorporación de información de las dinámicas poblaciones y funcionales de la mayor parte de las especies, la identificación de las especies más sensibles a los cambios del sistema.
Los modelos matriciales estructurados etariamente constituyen la base para los estudios de viabilidad poblacional y para la evaluación del estado de poblaciones de plantas (Chirakkal y Gerber 2010, Bialic-Murphy et al., 2022). La denominada matriz de Leslie es una herramienta conveniente en la descripción de los procesos demográficos (Damgaard 2005), que considera la densodependencia, las probabilidades de transición y la fecundidad. En relación con la densodependencia en los modelos poblacionales, debe tenerse en cuenta que cuando se presentan las clases etarias (plántulas, juveniles y adultos), estos interactúan de maneras diferentes, dependiendo del tipo funcional al que pertenece una especie (Valiente-Banuet y Verdu 2008, Maestre et al. 2009, Soliveres et al. 2011, Salmerón-López y Geada-López 2018).
El bosque semideciduo micrófilo de la Reserva Ecológica Siboney Juticí ha sido estudiado previamente por Salmerón-López et al. (2016a, Salmerón-López et al. 2016b; 2017); Salmeron-López y Geada-López (2018) desde una perspectiva ecológica. En este se definen tres tipos funcionales de plantas según el rol que desempeñan en la dinámica postperturbación: cobertoras, colonizadoras y estabilizadoras y solo una parte de estas son las que promueven los cambios durante la recuperación llamadas dinamogenéticas (Salmerón-López et al. 2016a). Se reportan diferencias significativas en la estructura regenerativa entre los tipos funcionales de respuesta a las perturbaciones, así como entre los sitios sometidos a diferentes niveles de perturbación Salmerón-López et al. (2016b). Se documentan interacciones tanto de facilitación como de competencia en sitios sometidos a diferentes niveles de perturbación en este bosque. La facilitación es más frecuente en los sitios medianamente y muy perturbados, y resulta más demandada por las plántulas y juveniles de las especies colonizadoras. La competencia es más frecuente entre los adultos de casi todos los tipos funcionales en los sitios poco perturbados (Salmeron-López y Geada-López 2018).
En la actualidad, en el bosque existen sitios perturbados con características diferentes en cuanto a composición y estructura funcional (Salmerón-López et al. 2016a). En algunos sectores, aunque las perturbaciones han cesado no se logra la recuperación por lo que han conducido al sistema a estados de equilibrio que no evolucionarán a estados compatibles con las exigencias de conservación del área protegida en el corto o mediano plazo, a menos que se realicen acciones que aceleren el proceso de recuperación.
Por lo que, el objetivo de este estudio es desarrollar un modelo para simular las posibles dinámicas auto-organizativas en la composición y estructura funcional del bosque semideciduo micrófilo bajo diferentes escenarios de perturbación.
MATERIALES Y MÉTODOS
Para el estudio de las dinámicas autoorganizativas se consideraron las especies consideradas como dinamogenéticas del bosque semideciduo micrófilo: Adelia ricinella L., Amyris elemifera L., Banara minutiflora (A. Rich.) Sleumer, Bourreria virgata (Sw.) G. Don. Bursera simaruba (L.) Sargent., Citharexylum spinosum L., Coccoloba diversifolia Jacq., Colubrina elliptica (Sw.) Brizicky et W. L. Stern, Cordia gerascanthus L., Coulteria linnaei (Griseb.) Acev.-Rodr., Crossopetalum rhacoma Crantz, Croton lucidus L., Diospyros grisebachii (Hiern.) Standl., Erithalis fruticosa L., Erythroxylum havanense Jacq., Erythroxylum rotundifolium Lunan, Eugenia cowelli Britton et P. Wilson, Eugenia iteophylla Krug et Urb, Eugenia smonticola (Sw.) DC., Exostema caribaeum (Jacq.) Roem. et Schult., Guettarda cueroensis Britton, Guettarda elliptica Sw., Gymnanthes lucida Sw., Heterosavia bahamensis (Britton) Petra Hoffm., Melochia tomentosa L., Picrodendron baccatum (L.) Krug et Urb. ex Urb., Plumeria obtusa L., Pseudocarpidium avicennioides (A. Rich.) Millsp., Pseudocarpidium ilicifolium (A. Rich.) Millsp., Randia aculeata L., Rauvolfia ligustrina Willd. ex Roem. et Schult., Senna atomaria (L). H. S. Irwin et Barneby, Sideroxylon salicifolium (L.) Lam., Tabebuia myrtifolia (Griseb.) Britton var. myrtifolia, Tecoma stans (L.) Juss. ex Kunth, Thouinia trifoliata Poit., Vachellia macracantha (Humb. et Bonpl. ex Willd.) Seigler et Ebinger, Zanthoxylum fagara (L.) Sarg., Zanthoxylum pistaciifolium Griseb (Salmerón-López et al. 2016a). Además, se consideraron los tipos funcionales de respuestas a perturbaciones: cobertoras, colonizadoras y estabilizadoras, (Salmerón-López et al. 2016a) y la clasificación de los sitios en función del grado de perturbación: sitios muy perturbados, medianamente y poco perturbados.
Modelo conceptual del sistema boscoso
Para la elaboración de un modelo conceptual de la dinámica del bosque semideciduo micrófilo se consideró la información disponible de estudios sobre la composición por tipos funcionales, estructura regenerativa e interacciones de (Salmerón-López et al. 2016a), Salmerón-López et al. 2016b y Salmerón-López y Geada-López (2018).
Ecuaciones de cambio según las clases de edades y tipo funcionales
Para cada tipo funcional se establecieron las ecuaciones de cambio partiendo de la matriz de Leslie y considerando las clases de edades definidas por Salmerón-López et al. (2016b) (Ecuación 1, Ecuación 2, Ecuación 3 y Ecuación 4)
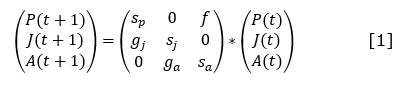
De donde:
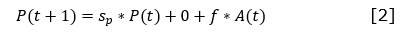
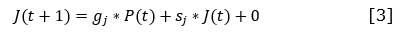
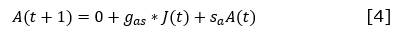
Donde:
P(t), P(t+1), J(t), J(t+1), A(t) y A(t+1) |
son el número de plántulas, juveniles y adultos en t y t +1 respectivamente |
f |
es la fecundidad de los adultos, entendida como el número de plántulas establecidas nuevas que aparecen en t +1 debidas a los adultos presentes en t. |
s p, sj y s a |
son las supervivencias de las plántulas, juveniles y adultos respectivamente, entendidas como el número de individuos del tiempo t que permanecerán como plántulas, juveniles o adultos en el t +1 |
g 1, y g 2 |
son las ganancias de juveniles y adultos en t +1, entendidas como el número de plántulas en t que se transforman en juveniles en t +1 () y el número de juveniles en t que se transforman en adultos en t +1 (g a |
En cada ecuación, se consideró la denso-dependencia de acuerdo al tipo funcional. Para cada especie se estimaron las tasas de vitalidad poblacional en las ecuaciones a partir de los datos de campo obtenidos durante el monitoreo del número de individuos en cada clase etaria, durante el periodo de 2016 a 2018. El monitoreo durante tres años consecutivos de los individuos permitió estimar los cambios en los números de individuos de plántulas, juveniles y adultos de cada especie y si estos cambios, en el caso de las plántulas y juveniles, se deben a la muerte del individuo o al paso del individuo a la clase de edad siguiente.
Para la estimación de los rasgos de vida y las tasas de cambio para cada una de las especies consideradas como dinamogenéticas (φ𝑖, 𝑑𝑝𝑖, 𝛾𝑗𝑖, 𝑑𝑗𝑖, 𝛾𝑎𝑖 y 𝑑??𝑖) se detallan en Salmerón-López (2016).
Simulación de las respuestas postperturbación del sistema
Para la simulación de la evolución temporal postperturbación de las poblaciones se elaboró un modelo en dinámica de sistemas para cada tipo funcional partiendo de las ecuaciones de cambio. En todos los casos se tuvieron en cuenta los siguientes aspectos: la no linealidad de la densodependencia de las poblaciones (debidas a interacciones intra e interespecíficas), la estocasticidad ambiental y las interacciones (facilitación o competencia) entre las especies (Salmerón-López y Geada-López 2018). En todos los casos, para la elaboración de los modelos en dinámica de sistemas se utilizó el programa STELLA 8.0.
Para la simulación de las perturbaciones se generaron tres posibles escenarios: Escenario 1- comienza con nivel de poco perturbado entre los años 100 al 120, después se incrementa a medianamente perturbado y en el año 160 a muy perturbado. Escenario 2- altas perturbaciones antrópicas concentradas hacia el inicio del régimen de perturbación, lo que elevó rápidamente el nivel a muy perturbado. Escenario 3. Inicia con un nivel medio de perturbaciones desde el año 90 hasta el año 180, y la evolución del sistema cuando estas cesan. En este caso, se mantuvo la simulación hasta 400 años para mostrar cómo responden los grupos funcionales.
Posteriormente, se comparan los resultados de las simulaciones con los datos de campo para los tipos funcionales en sitios sometidos a diferentes niveles de perturbación en la reserva. Atendiendo a los resultados de los estudios de campo como de las simulaciones se realizan recomendaciones para los tratamientos al bosque semideciduo micrófilo de la reserva.
RESULTADOS
Modelo conceptual del bosque semideciduo micrófilo
En el bosque semideciduo micrófilo de la Reserva Ecológica Siboney Juticí se detectó que en cada tipo funcional existía redundancia funcional en varias especies. Un grupo de estas especies se confirmaron como verdaderas dinamizadoras del proceso de recuperación (especies dinamogenéticas). Siete clasificaron como cobertoras, diez como colonizadoras y solo cuatro dentro de las estabilizadoras (Salmerón-López et al. 2016ab).
Las especies cobertoras, en su mayoría, fueron especies con frutos en forma de legumbre, folículos o cápsulas con semillas aladas o autocoras en ocasiones, plantas demandantes de luz. Estas incluyen a las especies que invaden y cubren los claros en los ecótopos correspondientes a estos bosques que generan una cobertura boscosa inicial de manera rápida.
Las especies colonizadoras se correspondieron en general con aquellas tolerantes a la sombra en sus estadios iniciales. Este tipo funcional se localizó en sitios correspondientes a casi todos los niveles de perturbación. En general fueron especies con habilidad para, una vez que los claros hayan sido previamente ocupados por las cobertoras, establecer sus plántulas y juveniles debajo del dosel arbóreo de las primeras y colonizar espacios que ya tienen una cobertura boscosa previa. Al llegar a la adultez, la mayoría de estas especies compiten con las cobertoras previamente establecidas.
Las especies estabilizadoras incluyeron también las tolerantes a la sombra, aunque en algunos casos no fue verificada esta tolerancia. Las plántulas, juveniles y adultos de estas especies estuvieron presentes tanto en sitios medianamente perturbados como en los sitios poco perturbados, pero generalmente con escasos individuos en los primeros.
Las tasas de incremento de cada especie, dependieron tanto de la densidad de individuos de la misma y de otras especies del mismo tipo funcional, como de la disponibilidad de espacios abiertos o cubiertos, según las exigencias del tipo funcional.
En el bosque semideciduo micrófilo, se han evidenciado perturbaciones antrópicas tales como: tala rasa, tala selectiva, establecimiento de cultivos, obtención de carbón, limpia de malezas, pastoreo e incendios. Algunas de estas perturbaciones tienen más incidencia sobre los adultos de todas las especies o de algunas de estas, como la tala rasa o selectiva y la obtención de carbón; mientras que la limpia de malezas, el pastoreo o los incendios afectan más a las plántulas o los juveniles. El establecimiento de cultivos afecta a todas las clases de edad. Varias de estas perturbaciones generan claros en la vegetación, pero el tamaño de estos depende del tipo y magnitud de la perturbación.
En el bosque, se reconocieron dos tipos de estas interacciones: de facilitación, cuando entre dos individuos la interacción fue positiva (uno de ellos facilita al otro) y de competencia, cuando la interacción es negativa (ambos compiten por espacio o luz). La facilitación se produce casi siempre por adultos de cobertoras sobre plántulas y juveniles de colonizadoras, aunque algunas estabilizadoras han sido verificadas como facilitadoras. La competencia, principalmente por la luz, se produce entre adultos de casi todos los tipos funcionales. Estas interacciones influyen también en las tasas de incremento de las especies, la magnitud de esta influencia estuvo condicionada a su vez por el tipo funcional y por la edad de las especies interactuantes.
El tránsito entre clases de edad para cada especie ha estado determinado, teóricamente, por dependencias anteriormente descritas, pero se verificó de manera diferente según la propia clase de edad y el tipo funcional al que corresponde la especie. Cambios en las interacciones al variar la clase etaria fueron verificables en el campo para varias especies.
Ecuaciones de cambio para los tipos funcionales
La deducción de las ecuaciones de cambio descritas en Salmerón-López (2016) se obtiene que, para las plántulas de todas las especies, en todos los tipos funcionales y partiendo de esta se obtuvo la siguiente ecuación (Ecuación 5):

Donde:
𝑃𝑖 |
Número de plántulas en el tiempo t |
𝐴𝑖 |
Número total de adultos de la especie i |
φ𝑖 |
Coeficiente de fertilidad de la especie i |
𝑑𝑝𝑖 |
Mortalidad de las plántulas de la especie i |
𝐾𝑝𝑖 |
Capacidad de carga del sitio para las plántulas de la especie i |
𝑔𝑗𝑖 |
Ganancia de juveniles de la especie i, entendida como el número de plántulas de esa especie en t transformadas en juveniles en t +1 |
El término 𝜃𝑝(𝑡) contiene la dependencia respecto a la densidad de plántulas de la propia especie en el sitio. Representa la proporción entre las plántulas ya establecidas en el año t y las que el espacio disponible para la especie puede asimilar en ese año, en otras palabras, que parte de la capacidad de carga para las plántulas de la especie ha sido ya cubierta.
El término Ω𝑝(𝑡) contiene la dependencia respecto a la densidad de plántulas del resto de las especies del mismo tipo funcional. Representa la proporción entre las plántulas del resto de las especies ya establecidas en el año t y las que el espacio disponible para esas especies, en ausencia de la especie i, puede asimilar, o, en otras palabras, que parte de la capacidad de carga para las plántulas del resto de las especies del mismo tipo funcional, si no existiera la especie i, ha sido ya cubierta.
La forma en que ambos parámetros dependen del espacio disponible es diferente para las especies cobertoras y para las especies colonizadoras y estabilizadoras. Para los juveniles de las cobertoras, partiendo de la ecuación [3], se obtuvo la siguiente ecuación (Ecuación 6) y (Ecuación 7):
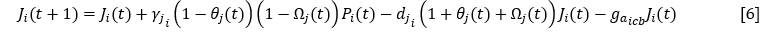
Y para las colonizadoras o estabilizadoras:
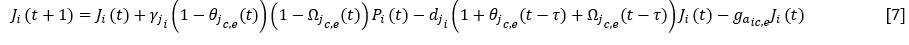
Donde:
𝛾𝑗𝑖 |
Tasa de incremento de juveniles de la especie i |
𝑑𝑗𝑖 |
Mortalidad de los juveniles de la especie i |
𝑔𝑎𝑖𝑐𝑏 (𝑔𝑎𝑖𝑐,𝑒): |
Ganancia de adultos de cobertoras (colonizadoras o estabilizadoras) a partir de los juveniles |
Los términos 𝜃𝑗(𝑡) y 𝜃𝑗𝑐,𝑒(𝑡) representan la proporción entre los juveniles de cobertoras, o de colonizadoras y estabilzadoras, ya establecidos en el año t y los que el espacio disponible para la especie puede asimilar, según el tipo funcional.
Los términos Ω𝑗(𝑡) y Ω𝑗𝑐,𝑒(𝑡) representan la proporción entre los juveniles del resto de las especies cobertoras, o colonizadoras y estabilizadoras, ya establecidos en el año t y los que el espacio disponible para esas especies, en ausencia de la especie i puede asimilar.
El término 𝜃𝑗𝑐,𝑒(𝑡 − 𝜏) expresa la proporción entre el número de juveniles de la especie colonizadora o estabilizadora i establecidos en el año t respecto del número de juveniles que podría asimilar un espacio equivalente al que se encontraba cubierto por los adultos totales (A T ) en el año t-ô, es decir, A T (t-ô), donde ô es el tiempo medio que demora un juvenil de la especie i en alcanzar la adultez.
De manera similar, Ω𝑗𝑐,𝑒(𝑡 − 𝜏), expresa la proporción entre el número de juveniles del resto de las especies colonizadoras o estabilizadoras establecidos en el año t respecto de la capacidad de carga media teórica del espacio que se encontraba cubierto por los adultos totales (A T ) en el año t-ô , en ausencia de la especie i.
Para los adultos de las especies cobertoras y, para algunas colonizadoras como Gymnanthes lucida, Heterosavia bahamensis y otras que necesitan espacios abiertos para llegar a la adultez, partiendo de la ecuación [4], se obtuvo la siguiente ecuación (Ecuación 8):

El término 𝜃𝑎(𝑡) expresa la proporción entre el número de adultos de la especie cobertora (y colonizadoras o estabilizadoras cuyos adultos demandan luz) establecidos en el año t, respecto del número de adultos que podría asimilar el espacio disponible para esa especie.
El término Ω𝑎(𝑡) representa la proporción entre el número de adultos del resto de las especies cobertoras establecidos en el año t respecto del número de adultos que podría asimilar el espacio disponible para esas especies en ausencia de la especie i.
Otras colonizadoras y algunas estabilizadoras pudieron alcanzar la adultez debajo de individuos adultos de otras especies de cualquiera de los tipos funcionales que dominen la cobertura del dosel arbóreo, o bien sus juveniles se pudieron convertir en adultos al quedar descubiertos. Estas especies, una vez que se establecen los juveniles, alcanzaron la adultez independientemente de que luego se mantengan o no cubiertos. En este caso se encontraron Adelia ricinella, Amyris elemifera¸ Erithalis fruticosa, Picrodendron baccatum. Estas se consideraron como subtipo 2.
Para estas especies la ecuación [4] tomó la forma (Ecuación 9):
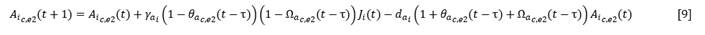
El término 𝜃𝑎𝑐,𝑒2(𝑡 − 𝜏) expresa la proporción entre el número de adultos de la especie colonizadora o estabilizadora i establecidos en el año t respecto del número de estos que podría asimilar un espacio equivalente al que se encontraba cubierto por los adultos totales (A T ) en el año t-ô, es decir, 𝐴𝑇(𝑡 − 𝜏) , cuando se establecieron los juveniles que dan lugar a estos adultos. es el tiempo medio que demora un juvenil de la especie i en alcanzar la adultez, independientemente de que el lugar donde se estableció se mantenga o no cubierto.
Respuestas a las perturbaciones en el bosque semideciduo micrófilo
El Anexo 1 muestra los módulos que componen el modelo de dinámica de sistemas elaborado desde de las ecuaciones de cambio para simular las dinámicas auto-organizativas, la información disponible sobre tipos funcionales, estructura regenerativa e interacciones funcionales (Salmerón-López 2016, Salmerón-López et al. 2016a, Salmerón-López et al. (2016b), Salmerón-López y Geada-López 2018), así como los rasgos de vida de las especies dinamogenéticas obtenidos del monitoreo en campo.
La Figura 1 muestra que todos los tipos funcionales se afectan, en particular las estabilizadoras. Al realizar un análisis para los adultos bajo el mismo Escenario 1 Figura 2), se observó que mientras los adultos de cobertoras mantuvieron un número elevado, los de colonizadoras casi se anularon, lo que sugiere que las colonizadoras mantuvieron la abundancia observada como plántulas o juveniles (Figura 1).
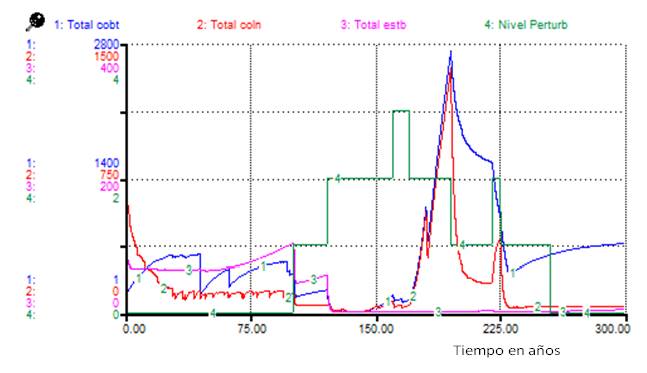 Nota: -Total cobt: total de cobertoras, Total coln: total de colonizadoras, Total estb: total de estabilizadoras, Nivel Perturb: nivel de perturbación
Nota: -Total cobt: total de cobertoras, Total coln: total de colonizadoras, Total estb: total de estabilizadoras, Nivel Perturb: nivel de perturbaciónFig. 1. - Cambios en el total de individuos por tipos funcionales bajo los efectos del Escenario 1 de perturbaciones
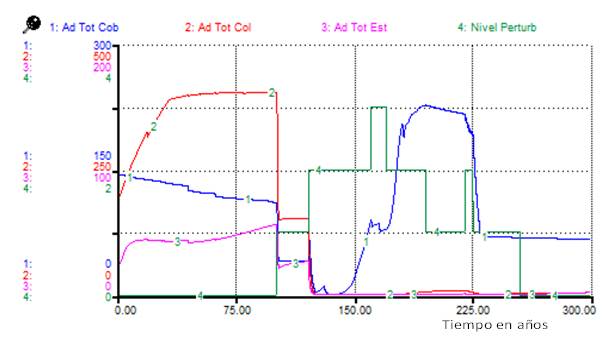 Nota: Ad Tot cob: adultos totales de las cobertoras, Ad Tot col: adultos totales de las colonizadoras, Ad Tot est: adultos totales de las estabilizadoras, Nivel Perturb: nivel de perturbación
Nota: Ad Tot cob: adultos totales de las cobertoras, Ad Tot col: adultos totales de las colonizadoras, Ad Tot est: adultos totales de las estabilizadoras, Nivel Perturb: nivel de perturbaciónFig. 2. - Cambios en el total de adultos por tipos funcionales bajo los efectos del Escenario 1 de perturbaciones
La Figura 3 muestra los cambios bajo el Escenario 2, en este caso el comportamiento para los individuos de los tres tipos funcionales fue similar al caso anterior, pero con diferencias en los totales y en las proporciones (Figura 3).
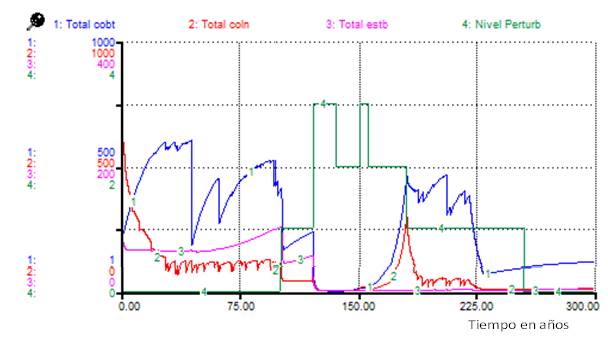 Nota: Total cobt: total de cobertoras, Total coln: total de colonizadoras, Total estb: total de estabilizadoras, Nivel Perturb: nivel de perturbación
Nota: Total cobt: total de cobertoras, Total coln: total de colonizadoras, Total estb: total de estabilizadoras, Nivel Perturb: nivel de perturbaciónFig. 3. - Cambios en el total de individuos para los tipos funcionales debidos al Escenario 2 de perturbaciones
Para los adultos de todos los tipos funcionales, se observa, aunque el número total de individuos fue mayor para cobertoras en el periodo postperturbación, los adultos predominantes fueron de especies colonizadoras, contrario a lo que se observó para el Escenario 1 (Figura 1) (Figura 4).
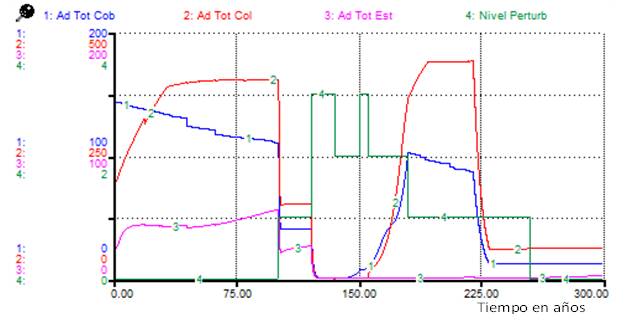 Nota: Ad Tot cob: adultos totales de las cobertoras, Ad Tot col: adultos totales de las colonizadoras, Ad Tot est: adultos totales de las estabilizadoras, Nivel Perturb: nivel de perturbación
Nota: Ad Tot cob: adultos totales de las cobertoras, Ad Tot col: adultos totales de las colonizadoras, Ad Tot est: adultos totales de las estabilizadoras, Nivel Perturb: nivel de perturbaciónFig. 4. - Cambios en el total de adultos para los tipos funcionales debidos al Escenario 2 de perturbaciones
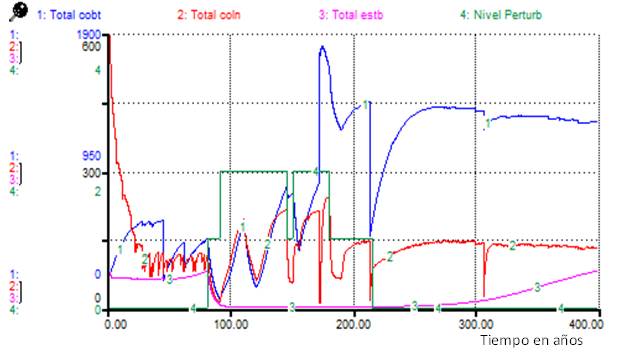
La Figura 5 representa el comportamiento de los totales por tipos funcionales durante la ocurrencia del "Escenario 3", en este el tipo funcional más afectado es el de las estabilizadoras que solo después de los 300 años comienzan a incrementar su abundancia los individuos de estabilizadoras.
DISCUSIÓN
Modelo conceptual del bosque semideciduo micrófilo
El modelo conceptual expuesto constituye una síntesis del conocimiento aportado por los estudios precedentes de este sistema, en cuanto a sus características funcionales y los elementos a tener en cuenta en el estudio de sus dinámicas autoorganizativas.
Las ecuaciones de cambio en el tiempo para los tipos funcionales tienen forma similar debido a que se parte de la matriz de Leslie (ecuación 1) considerando las mismas clases de edades en cada caso. Las diferencias esenciales en esas ecuaciones están dadas por los términos y para las plántulas, juveniles y adultos de los diferentes tipos funcionales. Estos términos contienen la manera que las diferentes clases de edad requieren espacios abiertos o cubiertos, así como las denso-dependencias respecto a los individuos de la misma o de otras especies. En todos los casos, son estos términos los que incluyen las relaciones no lineales de estas dependencias y los retrasos temporales en los cambios de clases de edad, confiriéndole un alto grado de incertidumbre a las respuestas del sistema a las perturbaciones o acciones de manejo que sobre el mismo se realicen.
Las ecuaciones no lineales propuestas para el sistema en el acápite de ecuaciones de cambio, constituyen la formalización de la información disponible sobre los roles funcionales de las especies que condicionan la dinámica postperturbación en el bosque semideciduo micrófilo.
Bajo los efectos del Escenario 1 los tres tipos funcionales se ven afectados (Figura 2), las primeras en recuperar su abundancia son las cobertoras, algo más retardadas las colonizadoras y por último las estabilizadoras. En la simulación, después de 200 años manteniéndose un nivel de perturbación, se afectan las estabilizadoras y las colonizadoras y abundan solo las cobertoras.
Los resultados de las simulaciones (Figura 1) se corresponden con lo que se observa en los sitios muy perturbados de Salmeron-López et al. (2016a), ubicados en áreas correspondientes a la cobertura denominada como bosque arbustivo con predominio de Vachellia macracantha por Reyes y Acosta (2005).
La comparación de los resultados de las simulaciones (Figuras 2 y Figura 4) sugiere que una evaluación del nivel de perturbación basada solamente en la fisonomía de la vegetación (atendiendo a cambios sucesionales) aproximadamente entre 180 y 220 años del inicio de las perturbaciones, podría indicar erróneamente la ocurrencia de perturbaciones diferentes, cuando solo se ha cambiado la distribución temporal de estas.
Los resultados demuestran como cambios ligeros en el régimen de perturbación pueden generar trayectorias de recuperación desiguales, con diferencias considerables en la estructura regenerativa, lo que se deriva de la no linealidad de tales procesos. Estos resultados argumentan lo planteado por varios autores respecto a la influencia de los rasgos funcionales de las especies y las interacciones entre estas, en el proceso de ensamblaje de las comunidades (Paine et al. 2011, Soliveres et al. 2011, Salmerón-López y Geada-López 2018).
En el caso de las simulaciones del Escenario 3, el incremento de los individuos de especies estabilizadoras alrededor de 100 años después de cesadas todas las perturbaciones (Figura 5), puede corresponderse con lo que se observa en el bosque actualmente, al pasar de estados medianamente perturbados a poco perturbados.
El modelo funcional elaborado permite captar las dinámicas auto organizativas del sistema y refuerza el criterio de que no solo el tipo de perturbación, sino la manera en que estas actúan sobre la estructura regenerativa del sistema al momento de ocurrir, influye en la dinámica pos perturbación.
La existencia de la composición funcional planteada en este estudio, de una determinada estructura regenerativa y de interacciones como las que en el modelo funcional se simulan, determinan que el sistema, a partir de regímenes de perturbación con diferencias ligeras (Escenarios 1 y 2), evolucione a estados diferentes. Tanto la existencia de diferencias funcionales en la respuesta a las perturbaciones (existencia de tipos funcionales) como la estructura regenerativa de estos tipos y sus interacciones, condicionan la dinámica de la recuperación postperturbación en el sistema. Lo anterior confirma la importancia de tener en cuenta los enfoques basados en la diversidad funcional para la elaboración y monitoreo, a largo plazo, de estrategias de manejo de ecosistemas y prácticas específicas de restauración conforme sostienen Gillison et al. (2013) y Vásquez-Valderrama y Solorza-Bejarano (2018).
Al comparar resultados de las simulaciones presentados en las Figuras 1 y 5, se verifica que en el primer caso (Figura 1), se coloca al sistema en condiciones en las que la recuperación de su integridad ecológica puede ser muy difícil, sin realizar acciones de manejo dirigidas a ese fin.
Tratamientos recomendados para el bosque semideciduo micrófilo
Las actividades dirigidas a la recuperación de la integridad del bosque semideciduo micrófilo en la Reserva Ecológica Siboney Juticí, deben tener en cuenta la existencia de tipos funcionales de respuesta a las perturbaciones en la vegetación y orientarse a la obtención de valores de diversidad funcional próximos a los de los sitios poco perturbados.
En las áreas descubiertas de vegetación, particularmente en sitios muy perturbados, se debe realizar la introducción de especies cobertoras, con el mantenimiento adecuado de la diversidad y favorecer aquellas especies identificadas como dinamogenéticas: Vachellia macracantha, Senna atomaria, Tecoma stans, Exostema caribaeum y Colubrina elliptica.
En los sitos identificados como muy perturbados, donde dominan pocas especies cobertoras, se deben introducir especies colonizadoras para propiciar el incremento de la diversidad de tipos funcionales. Como en el caso anterior, debe mantenerse una adecuada diversidad y favorecer las especies dinamogenéticas: Randia aculeata, Erythroxylum havanense, Bourreria virgata, Adelia ricinella. Luego de que estas especies hayan establecido plántulas y juveniles, se recomienda la introducción de las especies estabilizadoras: Guettarda elliptica, Erithalis fruticosa, Eugenia cowelli y Eugenia iteophylla.
Cualquier estrategia para la restauración del bosque semideciduo micrófilo en la Reserva Ecológica Siboney Juticí, debe prestar especial atención a la actual pobreza en la estructura regenerativa en las especies que condicionan la dinámica postperturbación, fundamentalmente en los sitios medianamente y muy perturbados. Estos sitios deben ser protegidos de las actividades que pueden impedir la conversión de plántulas, en juveniles y adultos.
En los sitios identificados como medianamente perturbados, se debe favorecer la cobertura de los claros con: especies cobertoras de rápido avance al estadio adulto (Vachellia macracantha y Senna atomaria); especies de rápido incremento en el número de individuos como Vachellia macracantha y Croton lucidus; las anteriores combinadas con especies cobertoras dinamogenéticas que parecen demandar claros de menores dimensiones como: Colubrina elliptica y Bursera simaruba.
En los procesos de restauración ecológica, en el bosque semideciduo micrófilo se debe prestar atención a la facilitación o la competencia (Salmeron-López y Geada-López 2018). Propiciar la utilización de las especies identificadas como facilitadoras y cuando se realize la plantación de las encontradas en categoría de facilitadas, atender su ubicación espacial debajo de la cobertura de las facilitadoras, para favorecer el éxito de la recuperación.
Teniendo en cuenta los cambios en el sentido de las interacciones al variar el estado regenerativo en algunas especies, se debe podar la copa de especies cobertoras: Vachellia macracantha y Senna atomaria, cuando bajo estas se hayan establecido juveniles de especies colonizadoras heliófilas en el estado adulto como Randia aculeata, Gymnanthes lucida, Heterosavia bahamensis, Coccoloba diversifolia y Erythroxylum havanense.
Algunos de los posibles comportamientos, resultantes de las simulaciones del modelo, pudieran no estar presentes en la Reserva, lo que no significa necesariamente que el modelo no refleje las condiciones del estudio. Significa, que el modelo funcional supera cualquier modelo mental posible, capaz de reflejar posibles evoluciones del sistema que no son deducibles de manera simple, sean estas observables o no en la práctica particularmente en zonas muy perturbadas.
Estos resultados refuerzan la necesidad de considerar la dinámica de sistema como una herramienta para la gestión de acciones en los planes de manejo de un área protegida.
CONCLUSIONES
El sistema de ecuaciones de cambio para el sistema constituye la formalización de la información disponible sobre los roles funcionales de las especies que condicionan la dinámica postperturbación en el bosque.
La posible aparición de unas u otras especies depende no solo del tiempo transcurrido después de las perturbaciones sino de las características del régimen de perturbación inicial del sitio.
La composición, estructura e interacciones funcionales junto con las características de las perturbaciones locales, condicionan las dinámicas auto-organizativas del sistema y pueden llevar por diferentes trayectorias sucesionales a más de un estado de equilibrio.
Anexo I
Modelos en dinámica de sistemas
Módulo tipos funcionales
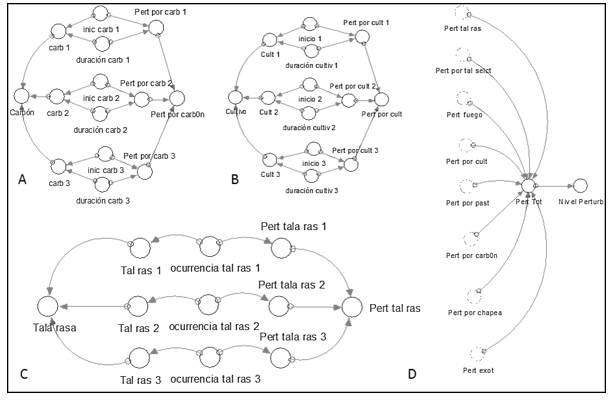
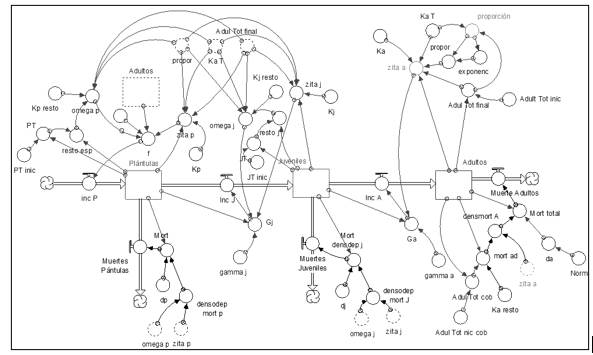
Tipo funcional cobertoras
El modelo corresponde al tipo funcional cobertoras (Figura 6).
Tipo funcional colonizadoras y estabilizadoras
Modelo en dinámica de sistemas para las especies colonizadoras y estabilizadoras que necesitan espacio descubierto para alcanzar la adultez (Figura 7).
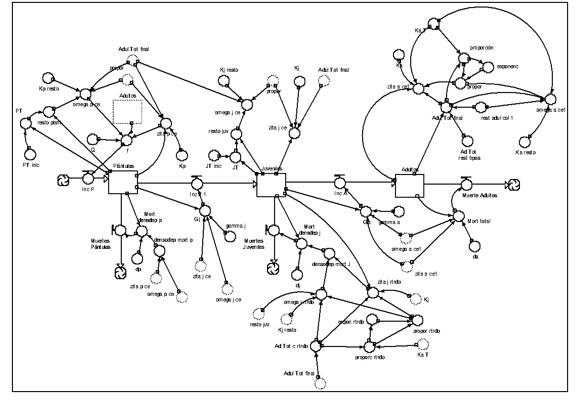
Fig. 7. - Modelo en Dinámica de Sistemas el para tipo funcional colonizadoras y estabilizadoras del tipo 1
Tipo funcional colonizadoras y estabilizadoras
Modelo en dinámica de sistemas para las especies colonizadoras y estabilizadoras que pueden alcanzar la adultez debajo de los adultos de otras especies (Figura 8).
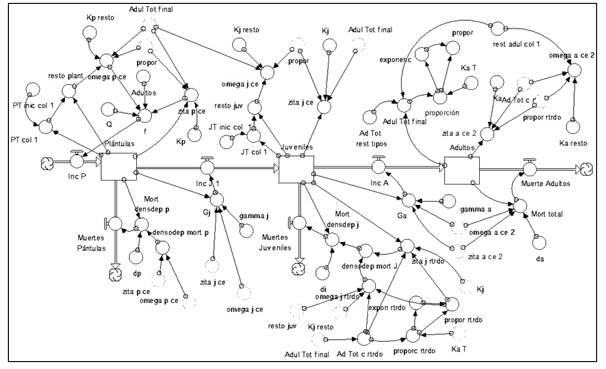
II.2. Módulo Interacciones
Modelo funcional correspondiente al módulo de interacciones. En este modelo se formalizó la información relacionada con los adultos de las especies verificadas como facilitadoras o competidoras excluyentes, con las variables de cambio de plántulas y juveniles de las especies sobre las cuales se pudo verificar alguna influencia (Figura 9).
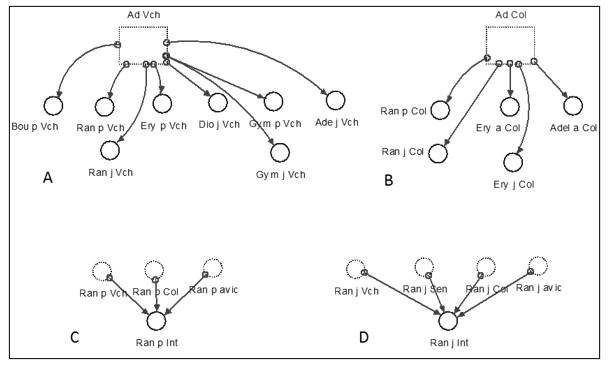 Nota: Ad Vch, Ad Cl (Adultos totales de Vachelia macracanta y Colubrina elliptica) Bou p Vch, Ran p Vch, Incremento de las plántulas de Randia aculeata, Bourreria virgata, debido a la presencia de adultos de Vachelia machracantha (o a Colubrina elliptica), lo mismo para Ran j Vch o Ran j Col, en este caso es el incremento de los juveniles de las especies. Ran p int, Ran j int, es el incremento total de las plántulas o juveniles de Randia aculeata por las interacciones totales con el resto de las especies.
Nota: Ad Vch, Ad Cl (Adultos totales de Vachelia macracanta y Colubrina elliptica) Bou p Vch, Ran p Vch, Incremento de las plántulas de Randia aculeata, Bourreria virgata, debido a la presencia de adultos de Vachelia machracantha (o a Colubrina elliptica), lo mismo para Ran j Vch o Ran j Col, en este caso es el incremento de los juveniles de las especies. Ran p int, Ran j int, es el incremento total de las plántulas o juveniles de Randia aculeata por las interacciones totales con el resto de las especies.Fig. 9. - Elementos del módulo de interacciones
II.3. Módulo de perturbaciones
En la Figura 9, se representa el módulo de perturbaciones del modelo. Las perturbaciones en este módulo corresponden con las expuestas por Salmerón-López et al (2016a): cultivos, fabricación de carbón, tala rasa, tala selectiva, incendios, limpia de malezas, y pastoreo, que coincidieron con las perturbaciones evidenciadas en el área. En el modelo, se consideró la magnitud, el momento y el tiempo pasado tras la ocurrencia para evaluar el nivel de perturbación de cada sitio en muy, poco o medianamente perturbado (Figura 10).