










Meu SciELO
 Serviços customizados
Serviços customizadosServiços Personalizados
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Cubana de Ciencias Forestales
versão On-line ISSN 2310-3469
Rev CFORES vol.11 no.1 Pinar del Río jan.-abr. 2023 Epub 03-Abr-2023
Original article
Model of self-organizational dynamics in a semi-deciduous microphyllous forest in eastern Cuba: a restoration approach
1Centro Oriental de Ecosistemas y Biodiversidad (BIOECO), Santiago de Cuba. Cuba.
2Universidad de Pinar del Río "Hermanos Saíz Montes de Oca". Pinar del Río, Cuba.
The use of systems modeling as a tool to simulate the dynamics in vegetal communities as responses to disturbances constitutes an alternative for management in protected areas. The objective of this study is to develop a model to simulate the possible self-organizing dynamics in the composition and functional structure of the microphyllous semi-deciduous forest under different disturbance scenarios. Population change equations for dynamogenetic species were determined based on the Leslie matrix, considering functional type and age structure. A conceptual model of the system is defined, that integrates the previous information available on functional composition, structure and interactions. The results of the simulations showed that disturbances occurred in some sectors of the area led the system to equilibrium states very different from the conserved states. The composition, structure, and functional interactions defined in the forest, together with local disturbances, condition the self-organizing dynamics of the system, which can lead to more than one state of equilibrium through different successional trajectories. These aspects facilitate the taking of actions for ecological restoration.
Key words: System dynamics models; disturbances; functional types; structure; nonlinear interactions.
INTRODUCTION
Self-organizing processes that occur after disturbances in plant communities, involve changes in populations and are strongly influenced by the functional traits of plants (Gillison et al., 2013, Verheijen et al., 2015), by the functional types present (Ostertag et al., 2015, Salmerón-López et al. 2016a, Vásquez-Valderrama and Solorza-Bejarano, 2018) and by the nature of plant-to-plant interactions (Salmerón-López and Geada-López 2018, Navarro-Cano et al., 2019).
Functional traits influence species presence, age structure, and regenerative states of the system in areas subjected to different levels of disturbance. In addition, these traits allow the definition of plant functional types, which constitute a valuable tool for the diagnosis of processes associated with disturbances (Gillison et al., 2013), in the long-term monitoring of ecosystem management strategies and specific restoration practices (Kooyman and Rossetto 2008, Vásquez-Valderrama and Solorza-Bejarano, 2018).
The use of system dynamics modeling is a way to understand possible changes in communities based on changes in composition, structure, interactions and functioning, but for this, it is necessary to incorporate information on the population and functional dynamics of most species and to identify the species most sensitive to changes in the system.
Ethnically structured matrix models are the basis for population viability studies and for the assessment of plant population status (Chirakkal and Gerber 2010, Bialic-Murphy et al., 2022). The so-called Leslie matrix is a convenient tool in the description of demographic processes (Damgaard 2005), which considers densodependence, transition probabilities and fecundity. In relation to densodependence in population models, it should be taken into account that when age classes (seedlings, juveniles and adults) are present, these interact in different ways, depending on the functional type to which a species belongs (Valiente-Banuet and Verdu 2008, Maestre et al., 2009, Soliveres et al., 2011, Salmerón-López and Geada-López 2018).
The semi-deciduous microphyllous forest of the Siboney Juticí Ecological Reserve has been previously studied by Salmeron-López et al. (2016a, Salmeron-López et al. (2016b; 2017); Salmeron-López and Geada-López (2018) from an ecological perspective. In this forest, three functional types of plants are defined according to the role they play in post-disturbance dynamics: cover, colonizing and stabilizing; and only a part of those (dynamogenetic) promote changes during recovery (Salmeron-López et al. 2016a). Significant differences in regenerative structure are reported among functional types in response to disturbances, as well as among sites Salmerón-López et al. (2016b). Both facilitation and competition interactions are documented in sites subjected to different levels of disturbance in this forest. Facilitation is more frequent in moderately and highly disturbed sites, and is more demanded by seedlings and juveniles of colonizing species. Competition is more frequent among adults of almost all functional types on lightly disturbed sites (Salmeron-López and Geada-López 2018).
Currently, disturbed sites with different characteristics in terms of composition and functional structure exist in the forest (Salmeron-López et al., 2016a). In some sectors, although disturbances have ceased, recovery remains unachieved. Meaning, they have led the system to states of equilibrium that will not evolve to states compatible with the conservation requirements of the protected area in the short or medium term, unless actions are taken to accelerate the recovery process.
Therefore, the objective of this study is to develop a model to simulate the possible self-organizational dynamics in the composition and functional structure of the semi-deciduous microphyllous forest under different disturbance scenarios.
MATERIALS AND METHODS
For the study of self-organizational dynamics, the species considered as dynamogenetic of the semideciduous microphyll forest were considered: Adelia ricinella L., Amyris elemifera L., Banara minutiflora (A. Rich.) Sleumer, Bourreria virgata (Sw.) G. Don. Bursera simaruba (L.) Sargent, Citharexylum spinosum L., Coccoloba diversifolia Jacq., Colubrina elliptica (Sw.) Brizicky et W. L. Stern, Cordia gerascanthus L., Coulteria linnaei (Griseb.) Acev.-Rodr., Crossopetalum rhacoma Crantz, Croton lucidus L., Diospyros grisebachii (Hiern.) Standl., Erithalis fruticosa L., Erythroxylum havanense Jacq, Erythroxylum rotundifolium Lunan, Eugenia cowelli Britton et P. Wilson, Eugenia iteophylla Krug et Urb, Eugenia smonticola (Sw.) DC., Exostema caribaeum (Jacq.) Roem. et Schult., Guettarda cueroensis Britton, Guettarda elliptica Sw., Gymnanthes lucida Sw., Heterosavia bahamensis (Britton) Petra Hoffm., Melochia tomentosa L., Picrodendron baccatum (L.) Krug et Urb. ex Urb., Plumeria obtusa L., Pseudocarpidium avicennioides (A. Rich.) Millsp. Pseudocarpidium ilicifolium (A. Rich.) Millsp., Randia aculeata L., Rauvolfia ligustrina Willd. ex Roem. et Schult., Senna atomaria (L). H. S. Irwin et Barneby, Sideroxylon salicifolium (L.) Lam., Tabebuia myrtifolia (Griseb.) Britton var. myrtifolia, Tecoma stans (L.) Juss. ex Kunth, Thouinia trifoliata Poit, Vachellia macracantha (Humb. et Bonpl. ex Willd.) Seigler et Ebinger, Zanthoxylum fagara (L.) Sarg., Zanthoxylum pistaciifolium Griseb Salmerón-López et al. (2016a. In addition, the functional types of responses to disturbances were considered: cover, colonizing and stabilizing, Salmerón-López et al. (2016a and the classification of sites according to the degree of disturbance: highly disturbed, moderately and lightly disturbed sites.
Conceptual model of the forest system
For the development of a conceptual model of the dynamics of the semi-deciduous microphyllous forest, the authors considered the information available from studies on composition by functional types, regenerative structure and interactions by Salmerón-López et al. (2016a, Salmerón-López et al. (2016b and Salmerón-López and Geada-López (2018).
Equations of change according to age classes and functional type
For each functional type, the equations of change were established starting from the Leslie matrix and considering the age classes defined by Salmerón-López et al. (2016b) (Equation 1, Equation 2, Equation 3, and Equation 4).
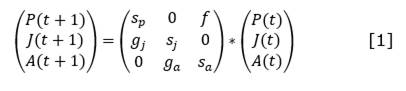
From where:
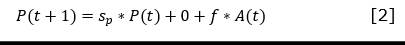
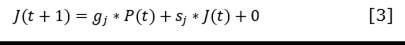
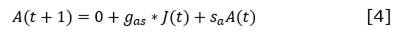
Where:
𝑃(𝑡), 𝑃(𝑡 + 1), 𝐽(𝑡), 𝐽(𝑡 + 1), 𝐴(𝑡) y 𝐴(𝑡 + 1) |
are the number of seedlings, juveniles and adults at t and t +1 respectively |
f |
is the fecundity of adults, understood as the number of new established seedlings appearing at t +1 due to adults present at t |
𝑠𝑝, 𝑠𝑗 and 𝑠𝑎 |
are the survivals of seedlings, juveniles and adults respectively, understood as the number of individuals at time t that will remain as seedlings, juveniles or adults at t +1 |
𝑔𝑗 and 𝑔𝑎 |
are the gains of juveniles and adults at t +1, understood as the number of seedlings at t that transform into juveniles at t +1 ( g j ) and the number of juveniles at t that transform into adults at t +1 (ga) |
In each equation, dense-dependence according to functional type was considered. For each species, population vitality rates were estimated in the equations from field data obtained during monitoring of the number of individuals in each age class, during the period from 2016 to 2018. Monitoring during three consecutive years of individuals allowed estimating changes in the numbers of seedling, juvenile and adult individuals of each species and whether these changes, in the case of seedlings and juveniles, are due to the death of the individual or the passage of the individual to the next age class.
For the estimation of life history traits and rates of change for each of the species considered as dynamogenetic, (φ𝑖, 𝑑𝑝𝑖, 𝛾𝑗𝑖, 𝑑𝑗𝑖, 𝛾𝑎𝑖 y 𝑑𝑎𝑖) are detailed in Salmerón-López (2016).
Simulation of the post-disturbance responses of the system
For the simulation of the post-disturbance temporal evolution of the populations, a system dynamics model was developed for each functional type, starting from the equations of change. In all cases, the following aspects were taken into account: the nonlinearity of population denso-dependence (due to intra- and interspecific interactions), environmental stochasticity and interactions (facilitation or competition) between species (Salmerón-López and Geada-López 2018). In all cases, STELLA 8.0 software was used for the development of the system dynamics models.
Three possible scenarios were generated for the simulation of disturbances: Scenario 1- starts with little disturbed level between years 100 to 120, then increases to moderately disturbed and in year 160 to very disturbed. Scenario 2 had high anthropogenic disturbances concentrated towards the beginning of the disturbance regime, which quickly raised the level to very disturbed. Scenario 3 begins with a medium level of disturbances from year 90 to year 180, and the evolution of the system when these ceases. In this case, the simulation was maintained up to 400 years to show how the functional groups respond.
Subsequently, the results of the simulations are compared with field data for the functional types in sites subjected to different levels of disturbance in the reserve. Based on the results of the field studies and simulations, recommendations are made for treatments for the semi-deciduous microphyllous forest in the reserve.
RESULTS
Conceptual model of the semi-deciduous microphyllous forest
In the semi-deciduous microphyllous forest of the Siboney Juticí Ecological Reserve, it was detected that in each functional type there was functional redundancy in several species. A group of these species were confirmed as true drivers of the recovery process (dynamogenetic species). Seven were classified as coverers, 10 as colonizers and only four as stabilizers (Salmerón-López et al. 2016ab).
Covering species, mostly, were species with legume-shaped fruits, follicles or capsules with winged or sometimes self-healing seeds, light-demanding plants. The species that invade and cover the clearings in the ecotones corresponding to these forests that generate an initial forest cover quickly are included.
The colonizing species generally corresponded to those tolerant to shade in their initial stages. This functional type was located in sites corresponding to almost all levels of disturbance. In general, they were species with the ability to establish their seedlings and juveniles under the tree canopy of the former, once the clearings had been previously occupied by the ground cover, and to colonize spaces that already had a previous forest cover. Upon reaching adulthood, most of these species compete with the previously established ground cover.
Stabilizing species also included shade tolerant species, although in some cases this tolerance was not verified. Seedlings, juveniles and adults of these species were present in both moderately disturbed and lightly disturbed sites, but generally with few individuals in the former.
The growth rates of each species depended on the density of individuals of the same species and other species of the same functional type, as well as on the availability of open or covered spaces, according to the requirements of the functional type.
In the semi-deciduous microphyllous forest, anthropic disturbances such as clear-cutting, selective logging, crop establishment, charcoal production, weeding, grazing and fires have been observed. Some of these disturbances have a greater impact on adults of all or some of the species, such as clear-cutting, selective logging and charcoal production, while weeding, grazing or fires affect seedlings or juveniles more. Crop establishment affects all age classes. Several of these disturbances generate clearings in the vegetation, but the size of these depends on the type and magnitude of the disturbance.
In the forest, two types of these interactions were recognized: facilitation, when the interaction between two individuals was positive (one of them facilitates the other) and competition, when the interaction is negative (both compete for space or light). Adults of groundcovers on seedlings and juveniles of colonizers mostly guarantee facilitation, although some stabilizers have been verified as facilitators. Competition, mainly for light, occurs between adults of almost all functional types. These interactions also influence the rates of species increase, the magnitude of this influence was at the same time conditioned by the functional type and by the age of the interacting species.
The transit between age classes for each species was theoretically determined by the dependencies described above, but was verified differently according to the age class itself and the functional type to which the species corresponds. Changes in interactions with varying age class were verifiable in the field for several species.
Equations of change for functional types
The derivation of the equations of change described in Salmerón-López (2016) obtained those seedlings of all species, in all functional types and starting from this, the following equation was obtained (Equation 5):
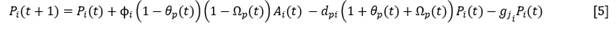
Where:
𝑃 𝑖 |
Number of seedlings at time t |
𝐴𝑖 |
Total number of adults of species i |
φ 𝑖 |
Fertility coefficient of species i |
𝑑𝑝𝑖 |
Mortality of seedlings of species i |
𝐾𝑝𝑖 |
Carrying capacity of the site for seedlings of species i |
𝑔𝑗𝑖 |
Gain of juveniles of species i, understood as the number of seedlings of that species at t transformed into juveniles at t +1 |
The term 𝜃𝑝(𝑡) contains the dependence with respect to the density of seedlings of the species itself at the site. It represents the ratio between the seedlings already established in year t and those that the space available for the species can assimilate in that year. In other words, part of the carrying capacity for seedlings of the species has already been covered.
The term Ω𝑝(𝑡)contains the dependence with respect to the density of seedlings of the rest of the species of the same functional type. It represents the ratio between the seedlings of the rest of the species already established in year t and those that the space available for those species, in the absence of species i, can assimilate. In other words, part of the carrying capacity for seedlings of the rest of the species of the same functional type, if species i did not exist, has already been covered.
The way in which both parameters depend on the available space is different for cover species and for colonizing and stabilizing species. For the juveniles of the groundcovers, starting from equation [3], the following equation was obtained (Equation 6 and Equation 7):
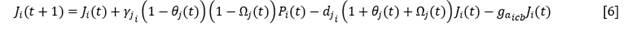
And for the colonizing or stabilizing ones:
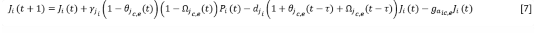
Where:
𝛾𝑗 𝑖i |
rate of increase of juveniles of species i |
𝑑𝑗𝑖i |
Mortality of juveniles of species i |
𝑔𝑎𝑖𝑐𝑏 (𝑔𝑎𝑖𝑐,𝑒) |
Gain of adults from juveniles for covers (colonizing or stabilizing) |
The terms 𝜃𝑗(𝑡) and Ω𝑗𝑐,𝑒(𝑡) represent the ratio between the juveniles of groundcovers, or colonizers and stabilizers, already established in year t and those that the space available to the species can assimilate, according to functional type.
The terms 𝜃𝑗𝑐,𝑒(𝑡 − 𝜏) and Ω𝑗𝑐,𝑒(𝑡) represent the proportion between the juveniles of the rest of the cover species, or colonizers and stabilizers, already established in year t and those that the space available for those species, in the absence of species i can assimilate.
The term 𝜃𝑗𝑐,𝑒(𝑡 − 𝜏) expresses the ratio between the number of juveniles of colonizing or stabilizing species i established in year t with respect to the number of juveniles that could assimilate a space equivalent to that which was covered by total adults (A_T) in year t-τ, that is, A_T (t-τ), where 𝜏 is the average time a juvenile of species i takes to reach adulthood.
Similarly, Ω𝑗𝑐,𝑒(𝑡 − 𝜏), expresses the ratio between the number of juveniles of the rest of the colonizing or stabilizing species established in year t with respect to the theoretical average carrying capacity of the space that was covered by total adults (A_T) in year t-τ , in the absence of species i.
For adults of covering species and, for some colonizers such as Gymnanthes lucida, Heterosavia bahamensis and others that need open spaces to reach adulthood, starting from equation [4], the following equation was obtained (Equation 8):
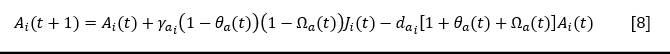
The term 𝜃𝑎(𝑡) expresses the ratio between the number of adults of the covering species (and colonizers or stabilizers whose adults demand light) established in year t, with respect to the number of adults that could assimilate the space available for that species. The term Ω_a (t) represents the ratio between the number of adults of the rest of the covering species established in year t with respect to the number of adults that could assimilate the space available for those species in the absence of species i.
Other colonizers and some stabilizers may have reached adulthood beneath adult individuals of other species of any of the functional types that dominate the arboreous canopy, or their juveniles may have become adults when uncovered. These species, once juveniles are established, reached adulthood regardless of whether then they remain covered or not. In this case, Adelia ricinella, Amyris elemifera¸ Erithalis fruticosa, Picrodendron baccatum were found and considered as subtype 2.
For these species the equation [4] took the form (Equation 9):
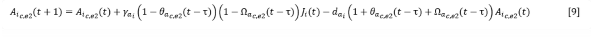
The term 𝜃𝑎𝑐,𝑒2(𝑡 − 𝜏) expresses the ratio between the number of adults of colonizing or stabilizing species i established in year t with respect to the number of these that could assimilate a space equivalent to that which was covered by total adults (𝐴𝑇) in year t-τ, that is, 𝐴𝑇(𝑡 − 𝜏) , when the juveniles that give rise to these adults were established. 𝜏 is the average time juvenile of species i take to reach adulthood, whether or not the site of establishment remains covered.
Responses to disturbances in the semi-deciduous microphyllous forest
Annex 1 shows the modules that compose the system dynamics model elaborated from the equations of change to simulate self-organizing dynamics, the available information on functional types, regenerative structure and functional interactions (Salmerón-López 2016, Salmerón-López et al., 2016a, Salmerón-López et al., 2016b, Salmerón-López and Geada-López 2018), as well as the life history traits of dynamogenetic species obtained from field monitoring.
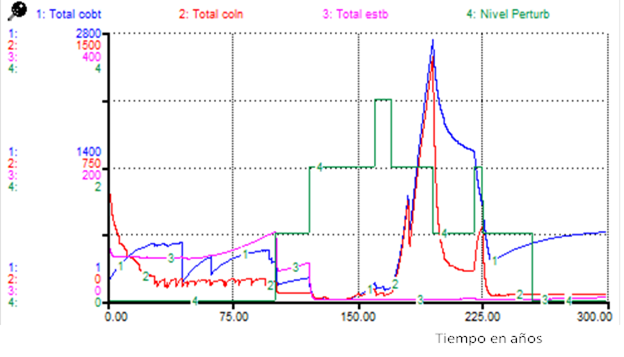
Figure 1 shows that all functional types are affected, particularly stabilizers. When performing an analysis for adults under the same Scenario 1 (Figure 2), it was observed that while covering adults maintained high numbers, those of colonizers almost cancelled out, suggesting that colonizers maintained the abundance observed as seedlings or juveniles (Figure 1).
 Note: Total cov: total cover, Total colo: total colonizers, Total stab: total stabilizers, Disturb level: disturbance level.
Note: Total cov: total cover, Total colo: total colonizers, Total stab: total stabilizers, Disturb level: disturbance level.Fig. 1. - Changes in the total number of individuals by functional type under the effects of disturbances (Scenario 1)
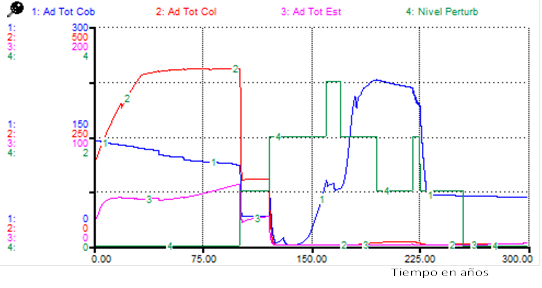 Note: Ad Tot cov: total adults of the cover, Ad Tot col: total adults of the colonizers, Ad Tot stab: total adults of the stabilizers, Disturb level: disturbance level
Note: Ad Tot cov: total adults of the cover, Ad Tot col: total adults of the colonizers, Ad Tot stab: total adults of the stabilizers, Disturb level: disturbance levelFig. 2. - Changes in total adults by functional type under the effects of disturbances (Scenario 1)
Figure 3 shows the changes under Scenario 2, in this case the behavior for the individuals of the three functional types was similar to the previous case but with differences in the totals and proportions (Figure 3).
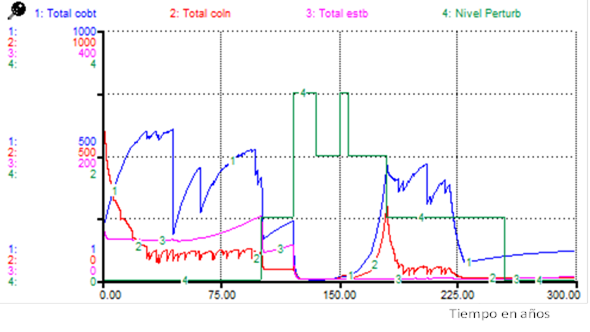 Note: Total cov: total coverts, Total colo: total colonizers, Total stab: total stabilizers, Disturb level: disturbance level
Note: Total cov: total coverts, Total colo: total colonizers, Total stab: total stabilizers, Disturb level: disturbance levelFig. 3. - Changes in the total number of individuals for the functional types due to disturbances (Scenario 2)
For the adults of all functional types, although the total number of individuals was higher for cover species in the post-disturbance period, the predominant adults were of colonizing species, contrary to what was observed for Scenario 1 ((Figure 1), (Figure 4).
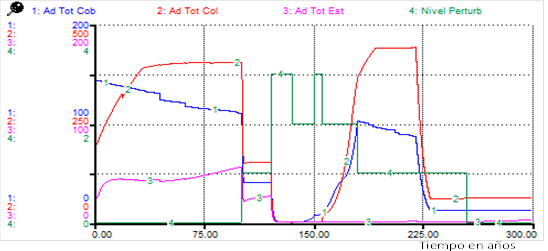 Note: Ad Tot cov: total adults of the coverts, Ad Tot col: total adults of the colonizers, Ad Tot stab: total adults of the stabilizers, Disturb level: level of disturbance.
Note: Ad Tot cov: total adults of the coverts, Ad Tot col: total adults of the colonizers, Ad Tot stab: total adults of the stabilizers, Disturb level: level of disturbance.Fig. 4. - Changes in total adults for functional types due to disturbances (Scenario 2)
Figure 5 shows the behavior of the totals by functional type during the occurrence of "Scenario 3", in which the most affected functional type is the stabilizers, which only after 300 years begin to increase their abundance of stabilizer individuals (Figure 5).
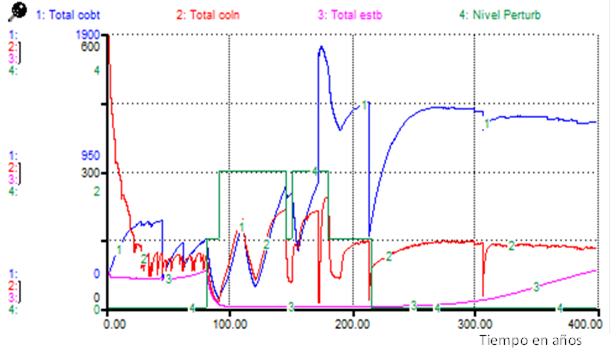
DISCUSSION
Conceptual model of the semi-deciduous microphyllous forest
The conceptual model presented here is a synthesis of the knowledge provided by previous studies of this system, in terms of its functional characteristics and the elements to be taken into account in the study of its self-organizational dynamics.
The equations of change over time for the functional types have a similar form because they are based on the Leslie matrix (Equation 1) considering the same age classes in each case. The essential differences in these equations are given by the terms 𝜃(𝑡) and 𝛺(𝑡) for seedlings, juveniles and adults of the different functional types. These terms contain the way the different age classes require open or covered spaces, as well as the dense-dependencies with respect to individuals of the same or other species. In all cases, it is these terms that include the non-linear relationships of these dependencies and the temporal delays in the changes of age classes, conferring a high degree of uncertainty to the responses of the system to disturbances or management actions that are carried out on it.
The non-linear equations proposed for the system in the section on equations of change, constitute the formalization of the available information on the functional roles of the species that condition the post-disturbance dynamics in the semi-deciduous microphyllous forest.
Under the effects of Scenario 1, the three functional types are affected (Figure 2), the first to recover their abundance are the cover species, followed by the colonizers and finally the stabilizers. In the simulation, after 200 years of maintaining a certain level of disturbance, the stabilizers and colonizers are affected and only the cover crops are abundant.
The results of the simulations (Figure 1) correspond to what is observed in the highly disturbed sites of Salmeron-López et al. (2016a), located in areas corresponding to the cover denominated as shrub forest with predominance of Vachellia macracantha by Reyes y Acosta (2005).
The comparison of the results of the simulations (Figures 2 and Figure 4) suggests that an evaluation of the level of disturbance based only on vegetation physiognomy (attending to successional changes) approximately 180 and 220 years from the onset of disturbances, could erroneously indicate the occurrence of different disturbances, when only the temporal distribution of these disturbances has changed.
The results demonstrate how slight changes in the disturbance regime can generate unequal recovery trajectories, with considerable differences in the regenerative structure, which derives from the non-linearity of such processes. These results argue what has been proposed by several authors regarding the influence of species functional traits and interactions between species on the community assembly process (Paine et al., 2011, Soliveres et al., 2011, Salmerón-López and Geada-López 2018).
In the case of the Scenario 3 simulations, the increase in individuals of stabilizing species around 100 years after the culmination of all disturbances (Figure 5), may correspond to what is observed in the forest today, moving from moderately disturbed to lightly disturbed states.
The functional model developed allows capturing the self-organizing dynamics of the system and reinforces the criterion that not only the type of disturbance, but also the way in which these acts on the regenerative structure of the system, at the moment of occurrence, influences the post-disturbance dynamics.
The existence of the functional composition proposed in this study, of a certain regenerative structure and of interactions such as those simulated in the functional model, determine that the system, from disturbance regimes with slight differences (Scenarios 1 and 2), evolves to different states. Both the existence of functional differences in the response to disturbances (existence of functional types) and the regenerative structure of these types and their interactions, condition the dynamics of post-disturbance recovery in the system. The previous statement confirms the importance of taking into account approaches based on functional diversity for the elaboration and monitoring, in the long term, of ecosystem management strategies and specific restoration practices as argued by Gillison et al. (2013) and Vásquez-Valderrama and Solorza-Bejarano (2018).
When comparing results of the simulations presented in Figures 1 and Figure 5, it is possible to verify that in the first case (Figure 1), the system is placed in conditions in which the recovery of its ecological integrity can be very difficult, without performing management actions directed to that purpose.
Recommended treatments for the semi-deciduous microphyllous forest
Activities aimed at recovering the integrity of the semi-deciduous microphyllous forest in the Siboney Juticí Ecological Reserve should take into account the existence of functional types of response to disturbances in the vegetation and should be oriented at obtaining functional diversity values close to those of sites with little disturbance.
In bare vegetation areas, particularly in highly disturbed sites, the introduction of cover species should be carried out, with adequate maintenance of diversity and favoring those species identified as dynamogenetic: Vachellia macracantha, Senna atomaria, Tecoma stans, Exostema caribaeum and Colubrina elliptica.
In sites identified as very disturbed, where few cover species dominate, colonizing species should be introduced to increase the diversity of functional types. As in the previous case, adequate diversity should be maintained and dynamogenetic species such as Randia aculeata, Erythroxylum havanense, Bourreria virgata, Adelia ricinella should be favored. After these species have established seedlings and juveniles, the introduction of stabilizing species (Guettarda elliptica, Erithalis fruticosa, Eugenia cowelli and Eugenia iteophylla) is recommended.
Any strategy for the restoration of the semi-deciduous microphyllous forest in the Siboney Juticí Ecological Reserve must pay special attention to the current poor regenerative structure in the species that condition the post-disturbance dynamics, especially in the medium and highly disturbed sites. Protecting these sites from activities that may impede the conversion of seedlings into juveniles and adults is a crucial action.
In the sites identified as moderately disturbed, the coverage of clearings should be favored with: cover species that advance rapidly to the adult stage (Vachellia macracantha and Senna atomaria); species with a rapid increase in the number of individuals such as Vachellia macracantha and Croton lucidus; the above combined with dynamogenetic cover species that seem to demand smaller clearings such as: Colubrina elliptica and Bursera simaruba.
In ecological restoration processes in the semi-deciduous microphyllous forest, attention should be paid to facilitation or competition (Salmeron-López and Geada-López 2018). It is necessary to propitiate the use of species identified as facilitators. Moreover, when planting those found as facilitated, it is vital attending their spatial location below the cover of the facilitators, to favor the success of the recovery.
Taking into account the changes in the direction of interactions when varying the regenerative state in some species, the top of cover species such as Vachellia macracantha and Senna atomaria should be pruned, when juveniles of heliophilous colonizing species such as Randia aculeata, Gymnanthes lucida, Heterosavia bahamensis, Coccoloba diversifolia and Erythroxylum havanense are established under them in the adult stage.
Some of the possible behaviors resulting from the model simulations may not be present in the Reserve, which does not necessarily mean that the model does not reflect the study conditions. It means that the functional model exceeds any possible mental model, capable of reflecting possible evolutions of the system that cannot be deduced in a simple manner, whether or not they are observable in practice, particularly in highly disturbed areas.
These results reinforce the need to consider system dynamics as a tool for managing actions in protected area management plans.
CONCLUSIONS
The system of equations of change for the system constitutes the formalization of the available information on the functional roles of the species that condition the post-disturbance dynamics in the forest.
The possible appearance of one or another species depends not only on the time elapsed after disturbances but also on the characteristics of the initial disturbance regime of the site.
The composition, structure and functional interactions together with the characteristics of the local disturbances, condition the self-organizational dynamics of the system and can lead through different successional trajectories to more than one state of equilibrium.
Annex I
System dynamics models
Module for Functional Types
Covering functional type
The model corresponds to the covering functional type (Figure 6).
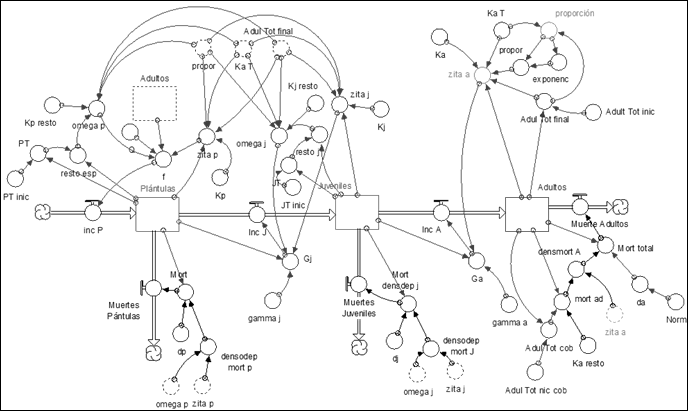
Colonizing and stabilizing functional type
System dynamics model for colonizing and stabilizing species that need open space to reach adulthood (Figure 7).
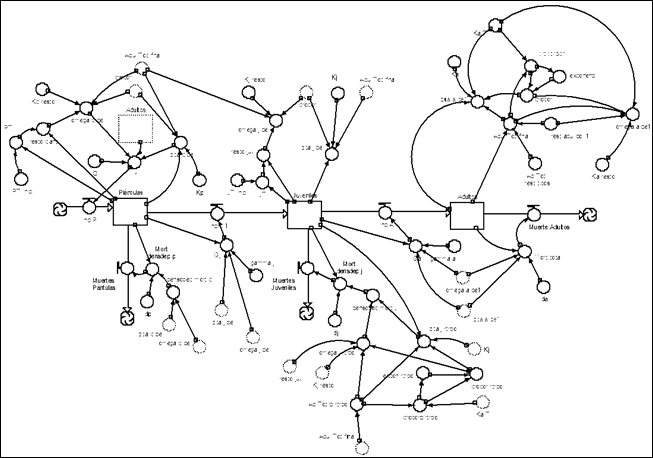
Colonizing and stabilizing functional type
System dynamics model for colonizing and stabilizing species that can reach adulthood below adults of other species Figure 8).
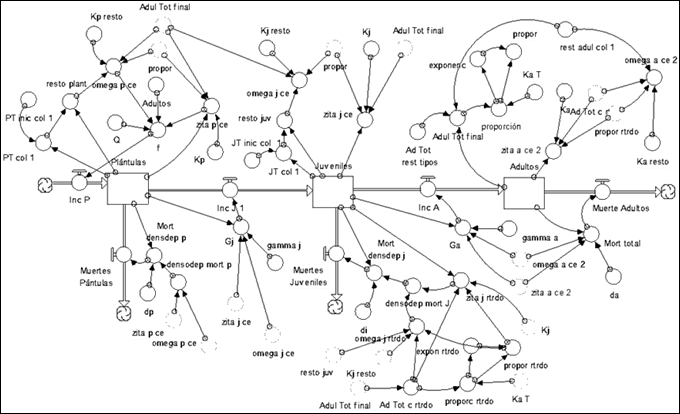
II.2. Interactions Module
Functional model corresponding to the interactions module. In this model the information related to the adults of the species verified as facilitators or excluding competitors was formalized, with the variables of seedling and juvenile change of the species on which some influence could be confirmed (Figure 9).
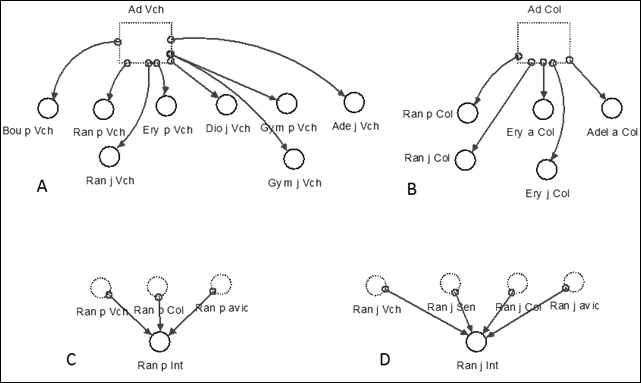 Note: Ad Vch, Ad Cl (Total adults of Vachelia machracantha and Colubrina elliptica) Bou p Vch, Ran p Vch, . Increase in seedlings of Randia aculeata, Bourreria virgata, due to the presence of adults of Vachelia machracantha (or Colubrina elliptica), the same for Ran j Vch or Ran j Col, in this case it is the increase in juveniles of the species. Ran p int, Ran j int is the total increment of seedlings or juveniles of Randia aculeata by the total interactions with the rest of the species.
Note: Ad Vch, Ad Cl (Total adults of Vachelia machracantha and Colubrina elliptica) Bou p Vch, Ran p Vch, . Increase in seedlings of Randia aculeata, Bourreria virgata, due to the presence of adults of Vachelia machracantha (or Colubrina elliptica), the same for Ran j Vch or Ran j Col, in this case it is the increase in juveniles of the species. Ran p int, Ran j int is the total increment of seedlings or juveniles of Randia aculeata by the total interactions with the rest of the species.Fig. 9. - Elements of the interaction module
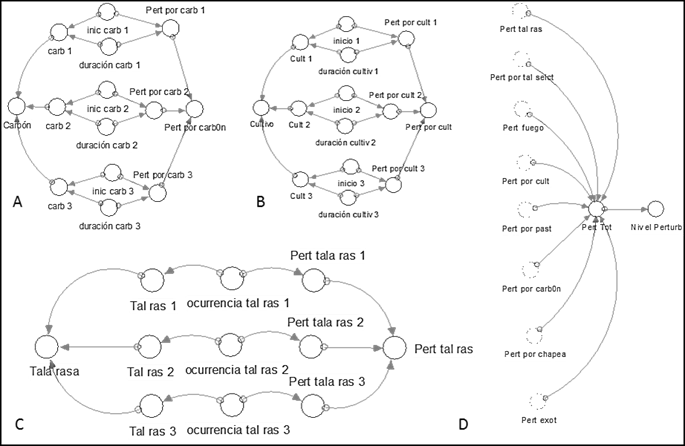
II.3. Module of disturbances
Figure 9 shows the disturbance module of the model. The disturbances in this module correspond to those presented by Salmerón-López et al. (2016a): crops, charcoal manufacture, clear cutting, selective logging, fires, weed clearing, and grazing, which coincided with the disturbances evidenced in the area. The model considered the magnitude, timing, and time passed after the occurrence to evaluate the level of disturbance of each site as very, little, or moderately disturbed (Figure 10).
REFERENCIAS BIBLIOGRÁFICAS
BIALIC-MURPHY, L., KNIGHT, T.M., KAWELO, K. y GAOUE, O.G., 2022. The Disconnect Between Short- and Long-Term Population Projections for Plant Reintroductions. Frontiers in Conservation Science [en línea], vol. 2. [Consulta: 06/03/2023 ]. ISSN 2673-611X. Disponible en: Disponible en: https://www.frontiersin.org/articles/10.3389/fcosc.2021.814863 [ Links ]
CHIRAKKAL, H. y GERBER, L.R., 2010. Short- and long-term population response to changes in vital rates: implications for population viability analysis. Ecological Applications [en línea], vol. 20, no. 3, pp. 783-788. [Consulta: 06/03/2023]. ISSN 1939-5582. DOI 10.1890/09-0560.1. Disponible en: Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1890/09-0560.1 . [ Links ]
DAMGAARD, C., 2004. Evolutionary Ecology of Plant-plant Interactions: An Empirical Modelling Approach [en línea]. S.l.: Aarhus University Press. ISBN 978-87-7934-116-6. Disponible en: https://books.google.com.cu/books/about/Evolutionary_Ecology_of_Plant_plant_Inte.html?id=8JjwAAAAMAAJ&redir_esc=y. [ Links ]
GEADA LÓPEZ, G., 2016. Estructura regenerativa funcional en el bosque semideciduo micrófilo de la Reserva Ecológica Siboney Juticí. Revista Forestal Baracoa [en línea], vol. 35, pp. 83-91. Disponible en: https://rc.upr.edu.cu/jspui/handle/DICT/2571 [ Links ]
GILLISON, A.N., BIGNELL, D.E., BREWER, K.R.W., FERNANDES, E.C.M., JONES, D.T., SHEIL, D., MAY, P.H., WATT, A.D., CONSTANTINO, R., COUTO, E.G., HAIRIAH, K., JEPSON, P., KARTONO, A.P., MARYANTO, I., N., G.G., VAN NOORDWIJK, M., SILVEIRA, E.A., SUSILO, F.-X., VOSTI, S.A. y NUNES, P.C., 2013. Plant functional types and traits as biodiversity indicators for tropical forests: two biogeographically separated case studies including birds, mammals and termites. Biodiversity and Conservation [en línea], vol. 22, no. 9, pp. 1909-1930. [Consulta: 06/03/2023]. ISSN 1572-9710. DOI 10.1007/s10531-013-0517-1. Disponible en: Disponible en: https://doi.org/10.1007/s10531-013-0517-1 . [ Links ]
KOOYMAN, R. y ROSSETTO, M., 2008. Definition of plant functional groups for informing implementation scenarios in resource-limited multi-species recovery planning. Biodiversity and Conservation [en línea], vol. 17, no. 12, pp. 2917-2937. [Consulta: 06/03/2023]. ISSN 1572-9710. DOI 10.1007/s10531-008-9405-5. Disponible en: Disponible en: https://doi.org/10.1007/s10531-008-9405-5 . [ Links ]
MAESTRE, F.T., CALLAWAY, R.M., VALLADARES, F. y LORTIE, C.J., 2009. Refining the stress-gradient hypothesis for competition and facilitation in plant communities. Journal of Ecology [en línea], vol. 97, no. 2, pp. 199-205. [Consulta: 06/03/2023]. ISSN 1365-2745. DOI 10.1111/j.1365-2745.2008.01476.x. Disponible en: Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1365 -2745.2008.01476.x . [ Links ]
NAVARRO CANO, J., GOBERNA, M. y VERDÚ, M., 2019. Plant facilitation as a tool to restore diversity and ecosystem functions. Ecosistemas [en línea], vol. 28, pp. 20-31. DOI 10.7818/ECOS.1747. Disponible en: https://www.researchgate.net/publication/334846896_Plant_facilitation_as_a_tool_to_restore_diversity_and_ecosystem_functions. [ Links ]
OSTERTAG, R., WARMAN, L., CORDELL, S. y VITOUSEK, P.M., 2015. Using plant functional traits to restore Hawaiian rainforest. Journal of Applied Ecology [en línea], vol. 52, no. 4, pp. 805-809. [Consulta: 06/03/2023]. ISSN 1365-2664. DOI 10.1111/1365-2664.12413. Disponible en: Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/1365-2664.12413 . [ Links ]
PAINE, C.E.T., BARALOTO, C., CHAVE, J. y HÉRAULT, B., 2011. Functional traits of individual trees reveal ecological constraints on community assembly in tropical rain forests. Oikos [en línea], vol. 120, no. 5, pp. 720-727. [Consulta: 06/03/2023]. ISSN 0030-1299. Disponible en: Disponible en: https://www.jstor.org/stable/29783565 . [ Links ]
RUIZ, A., 2005. Rapid Biological Inventories Report 10. Cuba: Siboney-Juticí. [en línea]. S.l.: s.n., pp. 47-50. ISBN 978-0-914868-58-3. Disponible en: https://www.researchgate.net/publication/257921577_Rapid_Biological_Inventories_Report_10_Cuba_Siboney-Jutici. [ Links ]
SALMERÓN-LOPEZ, A. 2016. Dinámica funcional del bosque semideciduo micrófilo en la Reserva ecológica Siboney Juticí, zona núcleo de la Reserva de la biosfera Baconao. https://rc.upr.edu.cu/jspui/handle/DICT/1952 [ Links ]
SALMERÓN LÓPEZ, A., GEADA LÓPEZ, G. 2018. Interacciones entre plantas en un bosque semideciduo micrófilo de Cuba Oriental. Bosque (Valdivia) [en línea], vol. 39, no. 2, pp. 347-356. [Consulta: 06/03/2023]. ISSN 0717-9200. DOI 10.4067/S0717-92002018000200347. Disponible en: Disponible en: http://www.scielo.cl/scielo.php?script=sci_abstract&pid=S0717-92002018000200347&lng=es&nrm=iso&tlng=es . [ Links ]
SALMERÓN LÓPEZ, A., GONZÁLEZ RODRÍGUEZ, A. y GEADA LÓPEZ, G., 2016. Tipos funcionales de plantas según su respuesta a las perturbaciones en un bosque semideciduo micrófilo costero de Cuba Oriental. Bosque (Valdivia) [en línea], vol. 37, no. 1, pp. 135-145. [Consulta: 06/03/2023]. ISSN 0717-9200. DOI 10.4067/S0717-92002016000100013. Disponible en: Disponible en: http://www.scielo.cl/scielo.php?script=sci_abstract&pid=S0717-92002016000100013&lng=es&nrm=iso&tlng=es . [ Links ]
SOLIVERES, S., ELDRIDGE, D.J., MAESTRE, F.T., BOWKER, M.A., TIGHE, M. y ESCUDERO, A., 2011. Microhabitat amelioration and reduced competition among understorey plants as drivers of facilitation across environmental gradients: Towards a unifying framework. Perspectives in Plant Ecology, Evolution and Systematics [en línea], vol. 13, no. 4, pp. 247-258. [Consulta: 06/03/2023]. ISSN 1433-8319. DOI 10.1016/j.ppees.2011.06.001. Disponible en: https://www.sciencedirect.com/science/article/pii/S1433831911000370. [ Links ]
VALIENTE-BANUET, A. y VERDÚ, M., 2008. Temporal Shifts from Facilitation to Competition Occur between Closely Related Taxa. Journal of Ecology [en línea], vol. 96, no. 3, pp. 489-494. [Consulta: 06/03/2023]. ISSN 0022-0477. Disponible en: Disponible en: https://www.jstor.org/stable/20143487 . [ Links ]
VÁSQUEZ VALDERRAMA, M. y SOLORZA-BEJARANO, J., 2017. Agrupación funcional de especies vegetales para la restauración ecológica de ecosistemas de montaña, Bogotá, Colombia. Colombia forestal [en línea], vol. 21, no. 1, pp. 5. DOI 10.14483/2256201X.11730. Disponible en: https://www.researchgate.net/publication/322014614_Agrupacion_funcional_de_especies_vegetales_para_la_restauracion_ecologica_de_ecosistemas_de_montana_Bogota_Colombia. [ Links ]
VERHEIJEN, L.M., AERTS, R., BROVKIN, V., CAVENDER-BARES, J., CORNELISSEN, J.H.C., KATTGE, J. y VAN BODEGOM, P.M., 2015. Inclusion of ecologically based trait variation in plant functional types reduces the projected land carbon sink in an earth system model. Global Change Biology [en línea], vol. 21, no. 8, pp. 3074-3086. [Consulta: 06/03/2023]. ISSN 1365-2486. DOI 10.1111/gcb.12871. Disponible en: Disponible en: https://onlinelibrary.wiley.com/doi/abs/10.1111/gcb.12871 . [ Links ]
Received: February 09, 2022; Accepted: March 06, 2023

