










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.31 no.3 La Habana sep.-dic. 2016
ORIGINAL ARTICLE
Induction of tomato resistance to Alternaria solani Sor. by biological and chemical activators in the field
Inducción de resistencia en tomate contra Alternaria solani Sor. mediante activadores químico y biológicos en campo
Simón Pérez-Martínez,I Ernestina Solórzano,II Daynet Sosa del Castillo,I,III Benedicto Martínez Coca,IV
IFacultad de Ingeniería, Universidad Estatal de Milagro (UNEMI), Km 1,5 vía al Km 26, Milagro 090150, Guayas, Ecuador.
II Facultad de Medio-Ambiente, Instituto Superior de Tecnologías y Ciencias Aplicadas, Habana, Cuba.
IIIEscuela Superior Politécnica del Litoral, ESPOL, Centro de Investigaciones Biotecnológicas del Ecuador (CIBE), Campus Gustavo Galindo Km 30.5 Vía Perimetral, P.O. Box 09-01-5863, Guayaquil, Ecuador.
IVGrupo de Fitopatología, Centro Nacional de Sanidad Agropecuaria (CENSA), San José de las Lajas, Mayabeque, Cuba.
ABSTRACT
Tomato early blight (Alternaria solani Sor.) is a prevalent disease in the humid subtropics of Western Cuba. The effect of pretreatment of the susceptible tomato cultivar HC-3880 with Glomus clarum, Arthrospira platensis and acibenzolar-S-methyl (ASM) was determined under field conditions. Seven treatments (mycorrhizas, spirulina, ASM, their combinations, and an untreated control) were studied to assess the plant enzymatic activity and response to the disease after artificial inoculations of a mixture of A. solani strains. The mycorrhizas were inoculated only once at planting by coating the seeds.The other inductors were applied by foliar spray, ASM one week and spirulina one day before inoculation of A. solani to all the plants, including the control, at 104 days of planting. The induction of six enzyme systems was determined at 0d, 1d, 7d and 10d after pathogen inoculation. After 10-12d of pathogen inoculation, the necrotic leaf area (NLA), number of spots per leaf (SPL) and yield were determined. The increase in enzyme activity and protection against the pathogen were minimal in the spirulina and control treatments. The combination mycorrhizas-ASM induced higher enzyme activity than the other treatments, with significant differences for glucanase, phenylalanine ammonia-lyase, peroxidase, and polyphenoloxidase. The NLA (14.04%) in this treatment was lower (but not significantly), than in the other five. The NLA discriminated the treatments better than the SPL and yield. In general, those treatments including ASM showed better results.
Key words: Arthrospira platensis, early blight, mycorrhizas, PR-proteins, systemic acquired resistance, spirulina.
RESUMEN
El tizón temprano (Alternaria solani Sor.) es una enfermedad prevalente en el subtrópico húmedo del occidente de Cuba. Se determinó el efecto de pretratamientos en la variedad susceptible al tizón temprano HC-3880 con Glomus clarum, Arthrospira platensis y acibenzolar-S-metilo (ASM) en condiciones de campo, contra A. solani. Se estudiaron siete tratamientos: mycorrhizas, spirulina, ASM, sus combinaciones y el Control (no tratadas) para determinar la respuesta de las plantas en términos de actividad enzimática y afectaciones foliares posterior a la infección artificial con A. solani. Las micorrizas se inocularon una sola vez mediante recubrimiento de las semillas y los otros agentes por aspersión foliar de cada uno. Todo el experimento fue asperjado con una mezcla de cepas de A. solani a los 104d, incluso el Control, cuando las plantas tenían 104d de tratadas con Micorriza, una semana con ASM y un día con Spirulina. Se determinó la inducción de seis sistemas enzimáticos a los 0d, 1d, 7d y 10d posteriores a la inoculación con el patógeno. A los 10-12d pos-inoculación se determinó el área foliar necrosada (NLA), el número de manchas por hoja (SPL) y el rendimiento. El incremento de las actividades enzimáticas fue mínimo en los tratamientos control y con spirulina, así como la protección contra el patógeno. La combinación de micorriza-ASM indujo mayor actividad enzimática respecto a los otros tratamientos, con diferencias significativas para glucanasa, fenilalanina amonio liasa, peroxidasa y polifenoloxidasa. El NLA (14,04%) fue menor (pero no significativamente) en este tratamiento, respecto a los otros cinco. El NLA discriminó mejor los tratamientos que el SPL y el rendimiento. En general, los tratamientos que incluyeron ASM mostraron mejores resultados.
Palabras claves: Arthrospira platensis, tizón temprano, micorrizas, PR-proteínas, resistencia sistémica adquirida, spirulina.
INTRODUCTION
Early blight disease of tomato (Alternaria solani Sor. is an endemic disease in hot and humid climates; it affects stems, fruits, and is particularly important due to the foliar necrosis it produces. Its control by fungicides, long-term crop rotations and soil fumigation (1) have been largely assayed. The complexity of the inheritance pattern and strong influence of the environmental conditions in natural epidemics of early blight hamper the commercial availability of resistant cultivars (1,2). Induction of resistance, especially by chemical or biological activators, is another alternative quite often used in crop protection to control the disease.
Among the chemical activators of the systemic acquired resistance are salicylic acid, its analogues 2,6-dichloro-isonicotinic acid and benzo (1,2,3) thiadiazole-7-carbothioic acid S-methyl ester, and its derivative benzo (1,2,3) thiadiazole with S methyl-benzo (1,2,3) thiadiazole-7-carbo-thiate. The latter, known as Acibenzolar-S-methyl (ASM), was developed to commercial product (BION®), and classified as plant activator and antifungal agrochemical (http://www.ebi.ac.uk/chebi/searchId.do?chebiId=CHEBI%3A73178). Induced resistance by chemicals such as ASM mimics the biological activation of systemic acquired resistance (SAR). ASM takes the place of salicylic acid (SA) in the SAR signal pathway inducing the same molecular markers and range of resistance (3). SAR is a long distance signaling mechanism that provides broad spectrum and long-lasting resistance to secondary infections throughout the plant (4). This unique feature makes SAR a highly desirable trait in crop production.
The resistance inductors have no direct antimicrobial effects, so it is assumed that the best results in field application are obtained by combining them with fungicides or antibacterial treatment (5). However, Fritz (15) obtained that ASM significantly inhibited the mycelial growth of Alternaria solani Sor. in vitro. Activation of resistance against A. solani in tomato is characterized by the induction of chitinase, β-1,3-glucanase, and lipoxigenase enzymes (6,7) in resistant cultivars. The ASM showed itself to be effective in the greenhouse, sufficing a single application of the product to achieve protection in the field; however, ASM does not always provide protection against pathogens (8).
Resistance in plants to both the avirulent or hypovirulent forms of a pathogen, like Fusariumm oxysporum Schltdl in tomato, can be induced by some biological active principles (9), or by symbionts such as the vesicular arbuscular mycorrhizas (VAM), which enhance tomato resistance to early blight by priming systemic defense response and the jasmonate signaling pathway (10). Several mycorrhizal species of Glomus, such as Glomus clarum (Nicolson & Schenck), Glomus mossae (Nicol. & Gerd./ Gerdeman & Trappe), and Glomus hoi-like (Berch & Trappe)(11,12), have shown beneficial effects on tomato. Glomus intraradices was highly efficient in harvest index values and fruit fresh weight, respectively (11). Enzymatic activities showed different responses in tomato seedlings according to VAM strains, being Glomus fasciculatum ((Thaxt.) Gerd. & Trappe) the most effective strain for this interaction under the studied conditions (13). In the market, there are commercial products whose active ingredients are different mycorhizal species, particularly Ecomic® in Cuba.
The cyanobacterium Arthrospira platensis ((Nordstedt) Gomont) (Spirulina) is marketed primarily as a nutritional supplement in humans (14). Particularly in Germany, it has been registered in the markets for organic products as a plant restorer, not been required to prove its effectiveness (15). Due to the presence of exopolysaccharides in its cultures, it is expected that there is a positive effect on the defense system of plants.
Achieving natural activation of the defense systems of plants as an alternative control is compatible for both cropping systems, where monoculture and the curative approach predominates, as well as in farms managed with an agroecological approach where the nonuse or reduction of traditional chemicals is imperative.
The objective of the research was to measure the protection by chemical and biological resistance activators to the damage of A. solani on a susceptible tomato cultivar in field trials by evaluating the kinetics of enzymes related to host defense systems and the leaf damage induced by the pathogen.
MATERIALS AND METHODS
Plant material
Plants of the tomato cultivar HC-3880 were used. This cultivar is susceptible to early blight and low inducing pathogenesis-related proteins (PR proteins) against A. solani (7).
The field experiment was carried out in Western Cuba, San José de Las Lajas, Mayabeque province, in winter from December 2003 to April 2004. Seedlings were sown in a dry heat sterilized mix of ferralitic red soil and peat contained in plastic trays. They were grown at room temperature in houses protected with anti-aphid mesh and ultraviolet-radiation-absorbent plexiglass roof for 45 days. Then, the seedlings were transplanted to the field in the morning at a planting distance of 0.7m x 0.25-0.30m. Previous soil analysis of the experimental area indicated an average number of viable young VAM spores of 18.6/g of soill, which was considered low (11).
Inoculation
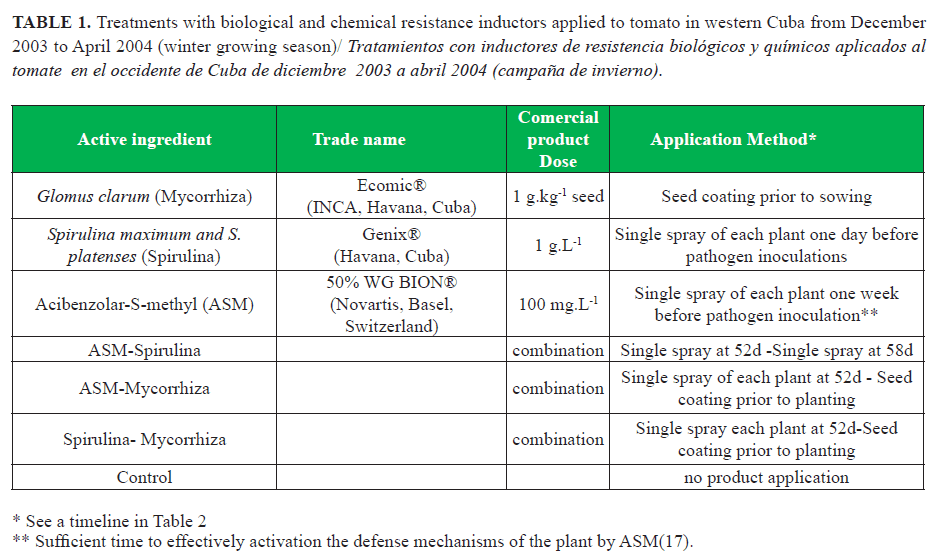
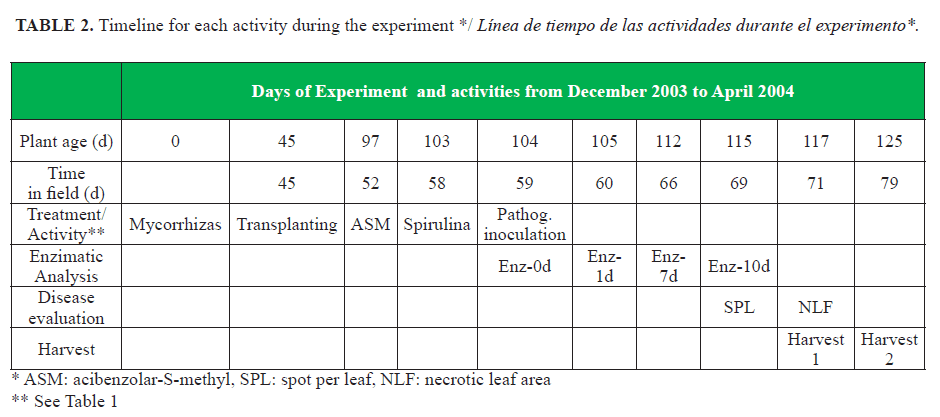
Table 1 shows the different resistance-inducing products and treatments. Doses of the commercial products were adjusted as recommended by the manufacturer, except in the case of Spirulina that was chosen arbitrarily. The experiment time was optimal for growing tomatoes, so that a low natural infection was expected. At 59 d post-transplant, all treatments were inoculated with a mixture of four Cuban pathogenic isolates of A. solani (Mycological Lab. National Center for Plant and Animal Health (CENSA), Mayabeque). The isolates were obtained from symptoms of tomato early blight with different degrees of aggressiveness against tomato genotypes (16).
The inoculum was prepared in static culture of each isolate in potato dextrose broth (200 g of fresh potato / 20 g dextrose), cultured for eight days in darkness at 27±1°C. Subsequently, the mycelium of each isolate was vacuum filtered through filter paper (Whatman # 1), weighed and proportionally fragmented into blender for 2-3 min. A final suspension of fresh mycelium in sterile distilled water (4.49 g.L-1) was prepared and each plant inoculated with 3-5 ml of the suspension with a 1-liter one-hand pressure sprayer after18:00 hours. It has been found that some inducers perform well when single pathogen isolates are used for inoculation, but not so well when mixtures of two or three pathogen isolates are used (8).
Enzymatic analysis
Leaf samples were taken from the plotstreated 59 days after planting, just before inoculation with the fungus. This time was considered time 0. Subsequently, samples at 1, 7 and 10d post-inoculation for the enzymatic dynamics (Table 2). Leaves between the 2nd to 4th positions from the ground were selected as the most susceptible to early blight (18).
Enzyme Extraction: a mixture of detached leaves was made and divided into three replicates of 5 g/treatment/time. The methodology (7) consisted of macerating with liquid nitrogen and homogenizing with sodium acetate (0.1 M; pH 5.2) in a 2:1 ratio (ml.g-1 fresh weight). The homogenate was incubated with stirring in an ice bath for 45 min, filtered through gauze and centrifuged (14.000 g /4°C/ 30min). The supernatant was stored at -20°C until use. Protein concentration was determined by the Bradford Coomassie brilliant blue assay is accomplished by measurement of absorbance at 595 nm. The standard bovine serum albumin curve was calibrated from a stock solution of 1mg.ml-1 (7).
Enzymatic activity: Each enzyme activity was determined using the methods previously standardized by Solórzano (7) and Rodríguez et al. (19).These methods can be described briefly as follows. ß-1,3-glucanase (GLUC): A discontinuous method based on laminarin (ß-1,3 glucan) hydrolysis. The product of reaction absorbs at 660 nm. Glucose (1 mg.ml-1) was used as standard in the calibration pattern curve. Phenylalanine ammonia lyase (PAL): a discontinuous method based on phenylalanine deamination. The reaction product, cinnamic acid, absorbs at 275 nm, and the reaction rate was determined with its readings. The calibration pattern curve was performed with cinnamic acid at1 mg.ml-1. Chitinase (CHIT): a discontinuous method based on the hydrolysis of the colloidal chitin to N-acetyl-glucosamine (585nm). Peroxidase (POX): a continuous method based on guaiacoloxidation. The reaction rate of oxidation was determined by recording the product absorbance at 470 nm. Variation in the optical density was determined for two minutes at 15 sec. intervals (ΔDO / At). Lipoxygenase (LOX): a continuous method where the linolenic acid is hydrolyzed (20). The reaction product absorbs at 234 nm. Polyphenoloxidase (PPO): a continuous method which is based on the oxidation of pyrogallol to quinones. The reaction product absorbs at 420 nm, the (ΔDO / Dt) was determined the same way as in the previous method. Enzyme activity was expressed inμmol of formed product min-1 mL enz-1, except where the PPO was expressed as ΔDO / Δt / min / ml of enzyme, as the extinction coefficient of pyrogallol was lacked (19). The specific activity of each enzyme was determined as Specific Act. = Enzime Act. Units / protein concentration (EAU.mg Prot-1).
Foliar damage by A. solani and yield
Leaf damage were determined by two recommended methods for this plant-pathogen interaction (18). After 10 days of inoculating the pathogen, the numbers of spots per leaf (SPL) were counted in three plants per treatment. At 12 days post-inoculation, the percentage of necrotic area per leaf (NLA) was determined in all plants by visual assessment by two evaluators (see time line in Table 2). The yield of the harvests at 70 and 79d from transplanting was measured. Not marketable fruits were discarded.
Experimental design and analysis
A random block design with three replications and seven treatments per block was used. Each treatment consisted of six rows with about 14 plants in each; the two outer rows were discarded to eliminate edge effects. Statistical analysis: Variance analysis was performed. The analysis of the nine response variables was integrated using analysis of the principal components with standardized data (due to the different scales) on a correlation matrix (21). This analysis was combined with a Biplot representation by Info-Stat program (Córdoba, Argentina: http://www.infostat.com.ar/).
RESULTS
Enzyme induction
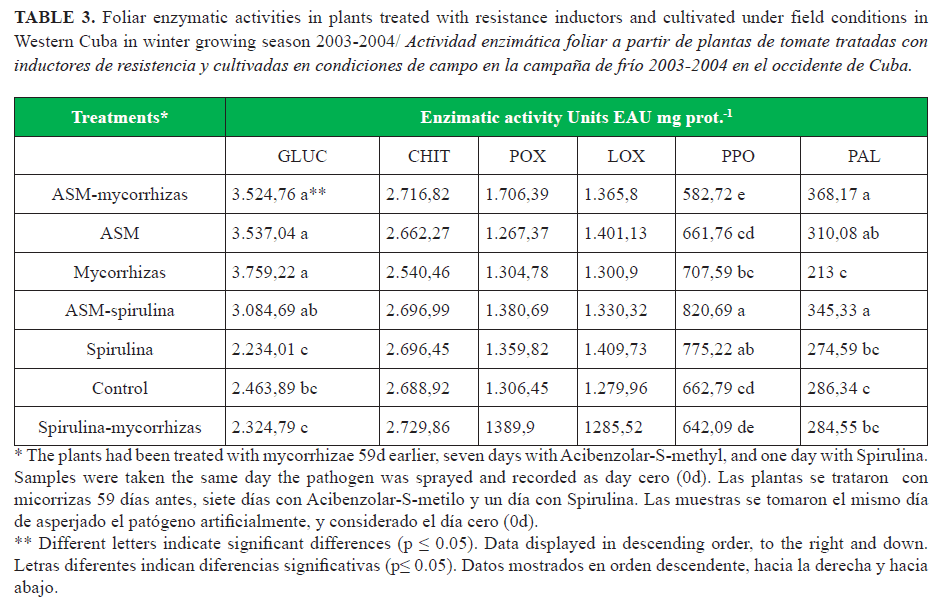
Detectable levels of all enzymes were observed in the control treatment (Table 3), confirming the existence of a constitutive enzyme level in the cultivar. The enzymes GLUC, PPO and PAL showed activities with significant differences between some of the treatments, unlike the other three enzymes (Table 3). GLUC was significantly high in the plants treated with mycorrhizas, acibenzolar-S-methyl (ASM) and the ASM-mycorrhizas combination. PPO was higher with spirulina and ASM-spirulina treatments. PAL particularly induced with ASM alone and with the combinations ASM-spirulina and ASM-mycorrhizas. The control plants showed the lowest enzyme levels in almost all treatments. It should be pointed out that there were a low level of mycorrhizas in the field blocks (22), between 0-40 young viable spores (data not shown).
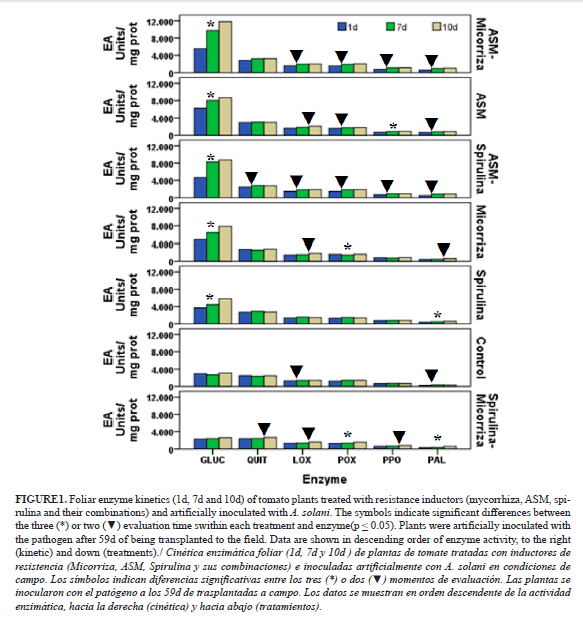
All treatments with acibenzolar-S-methyl induced the highest levels of foliar enzyme activity after A. solani inoculation (Fig. 1). The treatments with spirulina-mycorrhizas, spirulina alone, and the control (not treated) showed the lowest level of all the enzymes analyzed (Fig. 1). On average from all enzymes, covering the seeds with mycorrhizas showed an intermediate position among of all the treatments (Fig. 1). In most treatments, glucanase was notably the enzyme with higher level over 4.000 EAU mg prot.-1, but the effects of spirulina-mycorrhizas and the control. On the other hand, none of the treatments showed enzymatic activity over 4.000 EAU mg prot.-1 before pathogen inoculation (Table 3).The content of enzymes increased with time (1d to 10d) (Fig. 1), but not always significantly (see */▼). It should be noticed that only the treatment with mycorrhizas and spirulina-mycorrhizas stimulated some enzymes between 7d to 10d significantly (Fig. 2), while with most treatments the significance was shown between 1 and 7 days.
The enzyme CHIT showed no significant differences in none of the treatment at 0d, nor elicited at 24h after inoculations of the pathogen (data not shown), and in general at none of the times evaluated. Only in ASM-spirulina and spirulina-mycorrhizas treatments, the differences were significant (Fig. 1).
Ten days after inoculations with A. solani, the levels of all enzymes were two fold the levels of the day of inoculations (see 0d Fig. 1 and Fig. 2). Comparing the effects of the treatments at 10d, the ASM-mycorrhiza combination induced significantly higher level of enzymes than the worst treatments (control and spirulina-mycorrhizas). Moreover, all treatments that included ASM were statistically superior to the control, excepting ASM-spirulina that did not show significant differences. Minor and no significant differences were observed among the other treatments.
Foliar damage
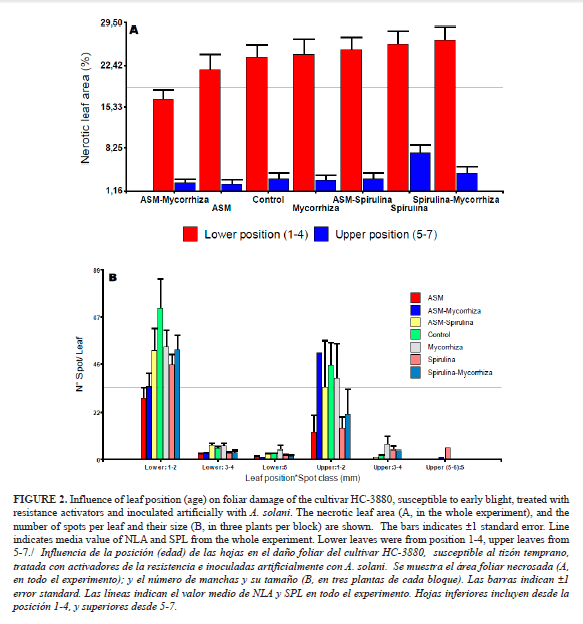
All inoculated plants were infected, although the level of infection in all treatments was low. The overall average was 18.04% (1-100%) for necrotic area and 22.33 for spots leaf-1 (1-157 spots), with the prevalence of 1-2 mm spots. Leaf necrosis development was influenced by its position on the plant (age) as expected for A. solani-tomato interaction (18). No significant differences among treatments were observed with ASM-mycorrhizas, although more damage was observed in all the treatments with spirulina (Fig. 2 A).
Figure 2 shows older leaves to be more susceptible to the pathogen infections with more necrotic areas (A) and generally more number of spots (B). Spot size 1-2 mm predominated in all treatments, irrespective of leaf position (Fig. 2 B). It must be pointed out that during this experiment, inoculum pressure remained low, judged by the surrounding tomato fields, which showed no early blight epidemics or significant damage. The site is characterized by low early blight apparent infection rate “r” and minimum temperatures of 20.1°C (23).
The maximum damage in terms of SPL and NLA matched the low-medium degrees of the scales proposed to evaluate the disease in the same province (18), where up to 20 spots were included in the lowest leaves 1-4.
The lowest percentage of NLA was achieved in the combination of ASM-mycorrhizas, while the spirulina treatment was the most affected (Fig. 2 A and B). The results showed herein suggested the ASM-mycorrhizas treatment to reduce the progression of 1-2 mm necrotic spots in the host (lowest NLA) rather than to prevent infection events (highest SPL). The opposite effect was only observed in the plants treated with Spirulina (Fig. 2).
Yield
The highest yield in this experiment was 12 kg total, equivalent to 3.8 TM.ha-1 and 0.48 kg.pl-1, which was achieved by coating the seeds with Mycorrhiza only. The other treatments followed with the values 10.25; 9.0; 4.95; 4.43; 3.68 and 3.30 kg for Spirulina-Mycorrhizas, Control, ASM-Spirulina, ASM-Mycorrhiza, Spirulina and ASM, respectively. However, these values are only for reference because they were the result of a block, the others were damaged at harvest by ants.
Integrated analysis
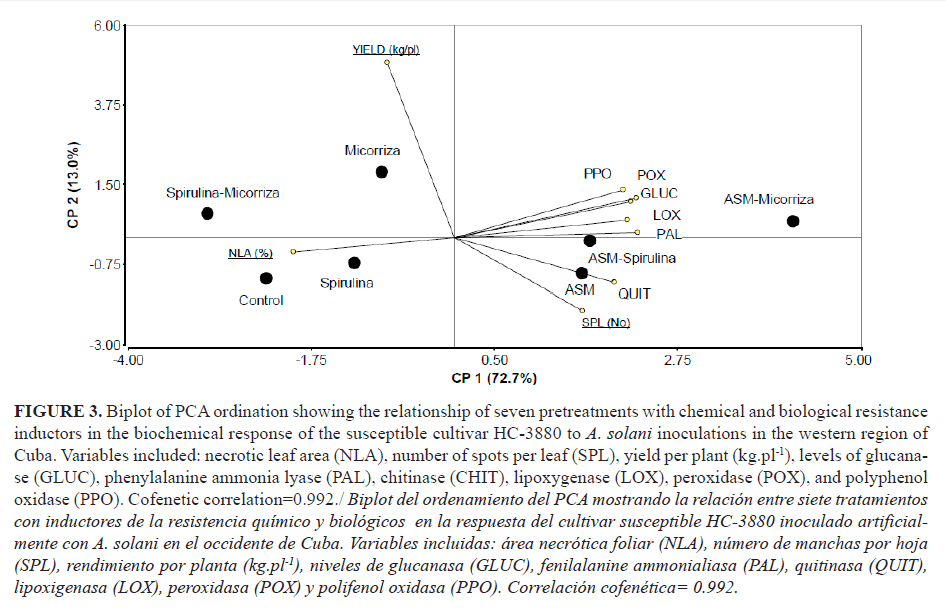
A comprehensive analysis conducted by the PCA-Biplot combination allowed reflecting simultaneously relationships among the treatments and the variables evaluated (Fig. 3). By the PCA, the first two components were found to explain a high proportion of the total variability (86.0%, see axes of Fig. 3).
The first principal component (CP1) was strongly and positively correlated with the original enzymatic variables, while NLA was also strongly correlated with these variables but negatively. This suggests that the treatments where ASM was included also tended to have more necrotic areas on the leaf, the second principal component increased with increasing yield.
Yield (kg.pl-1) position in the CP2 indicated that it had a strong correlation with it, which only explained 13% of the total variability. This result showed that the ASM-mycorrhizas significantly differed from the other treatments by the NLA and by the activity of the enzymes GLUC, PAL, POX and PPO, which showed significant differences at 10d.
DISCUSSION
The results achieved showed that a cultivar susceptible to early blight like HC-3880 could be protected from pathogen attack by the combined application of mycorrhizas and ASM. Untreated plants showed low enzyme levels, but not necessarily lower levels of pathogen infection. In fact, ASM-mycorrhizas was the only treatment that markedly (but not significantly) differed from five of the total seven treatments, considering the NLA.
At time of transplanting, the mycorrhizas are sufficiently established (13,19). From here onwards, the VAM-plant symbiosis enters into a parasitic stage (24), in which the fungal growth rate decreases and root colonization becomes low. The highest colonization levels are reached at 60 days, coinciding with the full flowering stage of the crop, when water and nutrients are highly required and the micorrhizas-plant symbiosis is more efficiently expressed. The level of fungal colonization begins to decrease at 120 days, a fact whichis closely related to host senescence (25).
The enzyme inducing effect of the micorrhizas, could be reinforced by ASM, since mycorrhizas or ASM by themselves showed no significant effects on reducing infection by A. solani. Plants pre-treated with ASM reduced severity of Clavibacter michiganensis subsp. michiganensis (Smith 1910), (Davis et al. 1984) up to 76.3% in tomato; this resistance was associated with a significant increase of the enzymes POX and CHIT (6). The mechanism of action of ASM, by either over- or under-expression, modifies genes encoding defense proteins (4). Similar studies showed that the content of phenolic compounds, PR-protein accumulation (such as CHIT and LUC) and PR-like thaumatins were observed more strongly in rice plants pretreated with ASM and inoculated with Xanthomonas oryzae pv. oryzae (26). Similar experiments in Australia, but with potato in production greenhouses and a dose of 100 mg a.i L-1 of ASM, reported an average in leaf damage of three spots per plant at 67d (27). In the same experiment, the field tests showed that foliar necrosis was only 20% of NLA. For Erysiphe cichoracearum DC, another foliar pathogen of potato, a dose of 50 mg i.a. L-1 of ASM was enough to reduce damage to less than 1% compared with 11.7% in the control under greenhouse conditions (27).
Spraying acibenzolar-S-methylol once at 55d after seeding showed promising results (Fig. 2). Despite the benefits of ASM, experiments in tomato leaf discs and different concentrations of A. solani inoculum showed contradictory ASM responses to pathogen control (15). Furthermore, fruit size reduction has been also reported in the tomato cultivar Easy Harvest treated with ASM, with significant dry weight reduction of about 50% in comparison with untreated plants (5). Even, the same authors indicated ASM had no measurable effect on the early and late blight of potato when applied without the accompaniment of fungicides underfield conditions in the UU.SS. Greenhouse aspersions resulted in almost complete control of Alternaria (27); however, its severity reduction was less impressive in the field. In general, yield was low in the experiment here in reported, despite the fact that it was performed in the optimum growing season (winter). In warmer field conditions in the eastern region of Cuba and with similar early blight damage (<10%), the tomato cultivar HC-3380 yielded less than 3 TM.ha-1 (28), and by far less than other cultivars with 2.20-2.75 kg.pl-1 in the western region in winter (16).
Spirulina showed no favorable results regarding the reduction of early blight damage (Fig. 2 A). Spirulina applications in greenhouse tests favored the reduction of dry and fresh tomato weight (15). The concentration of microalgae is not influential, in this case 1 g.L-1 was used and Fritz (15) obtained the same result in greenhouses with 2.5 and 10 g.L-1. However, an increase of two enzymes (GLUC and PAL) until 10d of confronting the plants with the pathogen was confirmed (Fig. 1). The same effect was shown on both NLA and the induced enzymes by combining spirulina with ASM or mycorrhizas.
By themselves, mycorrhizas and ASM showed no noticeable effects in reducing infection by A. solani. However, with their combination (at the planting time and a spray at 55d), a reduction in NLA was observed. From the practical perspective, it is useful to achieve protection of a cultivar like HC-3880, which, even being susceptible, is prized for its agro-ecological adaptability and for consumption (29). The favorable effect of MVA on tomato yield is well known under Cuban field conditions (25) as well as in reducing infection by A. solani in terms of necrosis or chlorosis compared withnon-mycorrhized plants (30). In practice, different methods of application of a phytosanitary product as well as the times and frequency of application determine the crop's response to disease(8). It has been suggested to combine acquired systemic resistance inductors to raise the levels of resistance early in the infected plants (31). The results reported here suggest that enhancement of natural resistance of tomato cultivars will help to reduce the number of fungicide applications even if they are susceptible to early blight.
In the case of A. solani, predominant pathogen populations in the agricultural ecosystem should be considered. Cuban populations of A. solani show intraspecific differences in genotyping, sensitivity to fungicides, and aggressiveness (15,32). Similarly, different isolates produce different biochemical responses depending on the cultivar (33). Excessive phosphorus availability is not favorable for VAM to express their potential in tomato (30), since levels of this element influence indirectly via reduced mycorrhizal colonization and this in turn reduces resistance.
It appears that the yield-improved modern cultivars have lost a considerable part of the basal and induced broad-spectrum resistance that characterizes their wild relatives and landraces (34). In addition to the combination of different products, it is necessary to determine the best variants in different localities. The presented results show a positive effect of combining seed coating with mycorrhizas and ASM application once the plants are established after transplantation. Activation of induced resistance by means of the six enzymes included in this study can also protect the crop from other pathogens. It remains to determine the response of tomato under more favorable conditions for the pathogen infection, such as planting outside the optimum growing season, where temperatures favor the epidemic, or in areas with high rates of infection.
ACKNOWLEDGEMENTS
Project funded by the National Science and Technology Program of the Ministry of Science, Technology and Environment of Cuba. The first author is grateful to the Ecuadorian Program PROMETEO-SENESCYT for the financial support during the writing of this manuscript. We thank Maendy Fritz for the assistance during the execution of the experiments, Marta Álvarez for supplying seeds of cultivar HC-3880 and Eduardo Sictach for polish up the writing. We are also grateful to anonymous reviewers for their suggestions on the manuscript.
REFERENCES
1. Özer Çalış, Topkaya Ş. Genetic analysis of resistance to early blight disease in tomato. African J Biotechnol. 2011;10(79):18071-18077.
2. Kumar S, Srivastava K. Screening of Tomato Genotypes Against Early Blight (Alternaria solani) under field condition. The Bioscan. 2013;8(1):189-193.
3. Zuluaga AP, Vega-Arreguín JC, Fry WE. Transcriptome profile of acibenzolar-S-methyl-induced genes in tomato suggests a complex polygenic effect on resistance to Phytophthora infestans. Physiol Mol Plant Pathol. 2013;81:97-106.
4. Gao Q-M, Zhu S, Kachroo P, Kachroo A. Signal regulators of systemic acquired resistance. Frontiers in Plant Sci. 2015;6:228. doi: 10.3389/fpls.2015.00228.
5. Louws FJ, Wilson M, Campbell HL, Cuppels DA, Jones JB, Shoemaker PB, et al. Field Control of Bacterial Spot and Bacterial speck of tomato using a plant activator. Plant Disease. 2001;85(5): 481-488.
6. Baysal Ö, Soylu S, Soylu E. Induction of defence-related enzymes and resistance by the plant activator acibenzolar-S-methyl in tomato seedlings against bacterial canker caused by Clavibacter michiganensis ssp. michiganensis. Plant Pathol. 2003;52(6):747-753.
7. Solórzano E. Estudio de proteinas y enzimas de defensa en la interacción Lycopersicon esculentum Mill -Alternaria solani (Ellis & Martin) Jones y Grout. Universidad Agraria de la Habana. Centro Nacional de Sanidad Agropecuaria. PhD thesis. 2002. 100 pp.
8. Walters DR, Ratsep J, Havis ND. Controlling crop diseases using induced resistance: Challenges for the future. J Exp Bot. 2013;64(5):1263-1280.
9. Parvatha RP. Non-pathogenic strains. In: Recent advances. Pavarta (ed). Springer India; 2013.p. 103-120.
10. Song Y, Chen D, Lu K, Sun Z, Zeng R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front Plant Sci. 2015 Sep 28;6:786. doi: 10.3389/fpls.2015.00786.
11. Ley-Rivas JF, Sánchez JA, Ricardo NE, Collazo E. Efecto de cuatro especies de hongos micorrizógenos arbusculares en la producción de frutos de tomate. Agron Costarric. 2015;39(1):47-59.
12. Mujica Y. Inoculación de hongos micorrízicos arbusculares (HMA) por dos vías diferentes en el cultivo del tomate (Solanum lycopersicum L.). Cultiv Trop. 2012;33(4):71-76.
13. Rodríguez Y, de la Noval B, Fernández F, Rodríguez P, Rodríguez-Yon Y. Estudio comparativo del comportamiento de seis cepas de hongos micorrízicos arbusculares en su interacción con el tomate (Lycopersicon esculentum M. var “Amalia”). Ecol Apl. 2004;3(1-2).
14. Abomohra AE, El-shouny W, Sharaf M, Abp-eleneen M. Effect of Gamma Radiation on Growth and Metabolic Activities of Arthrospira platensis. Brazilian Arch Biol Technol. 2016;59:1-11.
15. Fritz M. Resistance induction in the pathosystem tomato-Alternaria solani . [Gießen]: Biometrie und Populations genetik. Pflanzenbau und Pflanzenzüchtung II der Justus-Liebig-Universität Giessen; 2005.
16. Álvarez M, Moya C, Dominí ME, Arzuaga J, Martínez B, Pérez-Martínez S, et al. “ Amalia”: A Medium-fruit-size, Heat-tolerant Tomato Cultivar for Tropical Conditions. HortScience. 2004;39(6):1503-15034.
17. Puerari HH, Dias-Arieira CR, Dadazio TS, Mattei D, Da Silva TRB, Ribeiro RCF. Evaluation of acibenzolar-S-methyl for the control of Meloidogyne javanica and effects on the development of susceptible and resistant soybean. Trop Plant Pathol. 2013;38(1):44-48.
18. Cornide MT, Izquierdo F. Aplicación de dos métodos evaluativos para el estudio de la resistencia horizontal de variedades y líneas de tomate ante el ataque foliar del tizón temprano. Rev Ciencias Biológicas. 1979;10(2):245-261.
19. Rodríguez Y, Fernández A, Solórzano E, Peteira B, Fernández F, Rodríguez Yon Y. Inducción de enzimas de defensa en dos variedades de arroz (Oryza sativa) por el hongo micorrízico arbuscular G. mosseae. Rev Colomb Biotecnol. 2006;7(2): 35-49.
20. Solórzano E, Pilar M del. Inducción de lipoxigenasas en células de tomate tratadas con el hongo Alternaria solani. Rev Protección Veg. 2003;18(3):196-169.
21. Balzarini M, Bruno C, Arroyo A. Análisis de ensayos agrícolas multi-ambientales. Ejemplos con Info-Gen. Córdoba, Argentina; 2010. 141 p.
22. Morell F, Hernández A. Degradación de las propiedades agrobiológicas de los suelos ferralíticos rojos lixiviados por la influencia antrópica y su respuesta agroproductiva al mejoramiento. Agron Trop. 2008;58(4):335-343.
23. Gómez G, Almaguel L, Jimenez S, Secada I, Cortinas J. Zonificacion de Alternaria solani en papa en la provincia de La Habana. Fitosanidad. 1998;2(3-4):15-19.
24. Terry E. Microorganismos benéficos y productos bioactivos como alternativas para la producción ecológica de tomate (Solanum lycopersicum, Mill). Instituto Nacional de Ciencias Agrícolas. Universidad Agraria de la Habana, Instituto Nacional de Ciencias Agrícolas (INCA). PhD Thesis. Habana; 2005. 100 pp.
25. Terry E, Ruiz J. Evaluación de bioproductos para la producción de tomate (Solanum lycopersicum, Mill) bajo sistema de cultivo protegido. Cultiv Trop. 2008;29:11-15.
26. Mohan Babu R, Sajeena A, Vijaya Samundeeswari A, Sreedhar A, Vidhyasekeran P, Reddy M. Induction of bacterial blight (Xanthomonas oryzae pv. oryzae) resistance in rice by treatment with acibenzolar-S-methyl. Ann Appl Biol. 2003;143:333-340.
27. Bokshi AI, Deverall BJ, Morris SC. Effects of benzothiadiazole and acetylsalicylic acid on β-1,3-glucanase activity and disease resistance in potato. Plant Pathol. 2003;52(1):22-27.
28. Moya C, Álvarez M, J. Arzuaga, Fe C de la, Caballero A, Ponce M, et al. La selección participativa de variedades (SPV) en el cultivo del tomate. Cultiv Trop. 2010;31(1):87-92.
29. Moya C, Álvarez M, Dominí ME, Arzuaga J. MARA, nueva variedad de tomate de mesa. Cultiv Trop . 2004;25(2):69.
30. Fritz M, Jakobsen I, Foged M, Thordal-Christensen J, Pons-Kühnemann J. Arbuscular mycorrhiza reduces susceptibility of tomato to Alternaria solani. Mycorrhiza. 2006;16(6):413-419.
31. Usman Ghazanfar M, Wakil W, Talib Sahi S. Induction of resistance in chickpea (Cicer arietinum L.) against Ascochyta rabiei by applying chemicals and plant extracts. Chil J Agric Res. 2011;71(1):52-62.
32. Lantigua S, Martínez-Coca B, Pons-Kühnemann J, Pérez-Martínez S. Grupos intraespecíficos en Alternaria solani de tomate y papa. Fitopatol Venez. 2008;21:44-50.
33. Shahbazi H, Aminian H, Sahebani N, Halterman DA. Biochemical evaluation of resistance responses of potato to different isolates of Alternaria solani. Phytopathology. 2010;100(5):454-459.
34. Córdova-Campos O, Adame-Álvarez RM, Acosta-Gallegos JA, Heil M. Domestication affected the basal and induced disease resistance in common bean (Phaseolus vulgaris). Eur J Plant Pathol. 2012;134(2):36.
Recibido: 6/11/2015
Aceptado: 8/05/2016
Simón Pérez-Martínez, Facultad de Ingeniería, Universidad Estatal de Milagro (UNEMI), Guayas, Ecuador. Email: sperezm2@unemi.edu.ec.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}