










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En la actualidad, uno de los mayores retos para la conservación y el manejo sostenible de la biodiversidad es mantener poblaciones viables en sus entornos naturales (Aitken et al., 2008). La conservación de la biodiversidad requiere igualmente un conocimiento razonable sobre la distribución de las especies y sus poblaciones (Margules y Sarkar 2007; Rehfeldt et al., 2015). Sin embargo, la falta de información detallada sobre la distribución geográfica de estas constituye una limitante ante tal demanda (Pinkard et al., 2015).
La información disponible para realizar iniciativas de conservación en sitios degradados o en aquellos compuestos por especies raras o escasamente estudiadas, muchas veces puede estar constituida por antecedentes incompletos (Rehfeldt et al., 2015). Esta dificultad para obtener información del sitio puede ser particularmente relevante cuando se intenta trabajar con especies o ecosistemas circunscritos a pequeños fragmentos inmersos dentro de una matriz alterada por el ser humano. La falta de información limita la capacidad para generar programas a gran escala, y como consecuencia las iniciativas propuestas, en muchos casos, tienen un éxito limitado. Por ende, un tema relevante en esta materia, es poder predecir cuáles especies o grupos de especies podrían obtener mejores resultados para ser utilizadas en proyectos de conservación para sitios específicos (Contreras-Medina et al., 2010; Morales 2012).
Para sobrepasar estas barreras de información, los modelos de distribución de especies son una valiosa herramienta para determinar la distribución de especies poco estudiadas y pueden ser de gran ayuda en la generación de información biológica base no disponible. Los modelos de distribución de especies son una herramienta empírica, robusta, repetible y fácil de utilizar, que pueden ayudar a identificar zonas potenciales a conservar y/o restaurar (Phillips y Dunik, 2008 y Arribas et al., 2012). El valor de estos estudios en la conservación está basado en que 1) indica potenciales sitios donde intervenir en el caso de necesitar hacer exploraciones directas, 2) entrega lineamientos para determinar el comportamiento de las especies frente a alteraciones exógenas, principalmente antrópicas y 3) entrega información sobre los sitios a conservar (Hernández et al., 2008 y Arribas et al., 2012).
Los algoritmos de modelado de distribución de especies toman las ubicaciones de recolección de una especie e identifican los valores de un conjunto de variables ambientales explicativas que inciden en esos sitios (Eliht et al., 2011) y devuelven las probabilidades proyectadas de presencia para esa especie a través de un área de estudio predefinida, dependiente de las variables de conocimiento. Siempre que la distribución geográfica de la especie esté bien representada por los sitios muestreados, los modelos deben identificar con precisión el nicho ecológico para la especie de interés.
Magnolia cubensis Urb. subsp. cubensis (marañón de Sierra Alta), es un taxón endémico del bosque pluvisilva de montaña en el oriente de Cuba, se desarrolla sobre los 800 m s.n.m de la Sierra Maestra y Sierra de Gran Piedra (Testé et al., 2019). Dentro del género Magnolia es uno de los taxones con más amplia distribución (Palmarola et al., 2015; Palmarola et al.,2016), pero con poblaciones fragmentadas dentro del macizo montañoso, es considerada dentro de las especies con madera preciosas cubanas (Molina-Pelegrín et al., 2014). En los últimos años, sus poblaciones naturales se han visto seriamente afectadas por el aprovechamiento maderero, otras actividades antrópicas y los eventos meteorológicos (Palmarola et al. 2015) por lo que la especie está categorizada como vulnerable (González-Torres et al., 2016). En la actualidad, se han iniciado una serie de actividades encaminadas a fortalecer su conservación y utilización sostenible, así como estudios básicos de su estructura poblacional en la Sierra de Gran Piedra (Testé et al., 2019).
Esta subespecie se encuentra sujeta a niveles de riesgo por experimentar reducciones potenciales en sus poblaciones, a causa de la falta de espacios naturales y escaso manejo. De ahí, el importante papel que juegan las áreas protegidas ya que gran parte de sus poblaciones se encuentran en estas, las acciones a realizar abarcan desde el conocimiento de los recursos del área y el manejo de especies amenazadas, hasta la educación ambiental e integración comunitaria a las tareas de conservación.
Todo lo anterior indica la importancia de realizar estudios de modelación de la distribución espacial de M. cubensis subsp. cubensis que contribuyan a la conservación de este taxón en la región oriental de Cuba. La presente investigación tiene por objetivo determinar la distribución potencial actual de M. cubensis subsp cubensis en el oriente de Cuba, como base para estudios de impacto de cambio climático e identificación de sitios para la conservación.
MATERIALES Y MÉTODOS
Caracterización del área de estudio
La subespecie tiene su distribución natural en la Sierra Maestra comprende cinco áreas protegidas: Parque Nacional La Bayamesa (PNLB) y Parque Nacional Turquino (PNT), áreas protegidas de significación nacional, Paisaje Natural Protegido Gran Piedra, así como, las Reservas Ecológicas Pico Caracas y El Gigante. Conforman el bloque montañoso por encima de los 1 200 m s.n.m. con mayor área en Cuba.
El clima del territorio es poco variado, aunque existen diferencias entre la parte norte y el borde meridional. La precipitación es elevada, pues oscila entre 1 800 y 2 300 mm anuales. La evaporación media anual es baja. La temperatura media anual varía entre 16°C y 20°C. Las temperaturas mínimas absolutas varían entre 12°C y 16°C, la media de las temperaturas máximas absolutas es de 26°C a 30°C, mientras que la media de las mínimas absolutas es de 4°C a 8°C (Montenegro, 1991).
En la Sierra Maestra, se encuentra una gran diversidad de formaciones vegetales: bosque nublado, matorral nublado, pluvisilva de montaña, pinares naturales, bosque siempreverde mesófilo, herbazal de galería y vegetación antrópica. Estas dependen fundamentalmente de las diferencias ecológicas, las que están condicionadas principalmente por la altitud y en ocasiones el edátopo (Maceira et al., 2005).
Magnolia cubensis subsp. cubensis, habita en la formación vegetal de pluvisilva de montaña, que predomina en sitios entre los 800 y los 1 400 m s.n.m. En las áreas superiores transiciona con el bosque nublado, y en las inferiores con el bosque siempreverde mesófilo. El relieve es abrupto, frecuentemente con pendientes de más de 30 grados sobre suelos ferralíticos rojos lixiviados muy pobres y ácidos (Testé et al., 2019).
Distribución geográfica
Distribución real
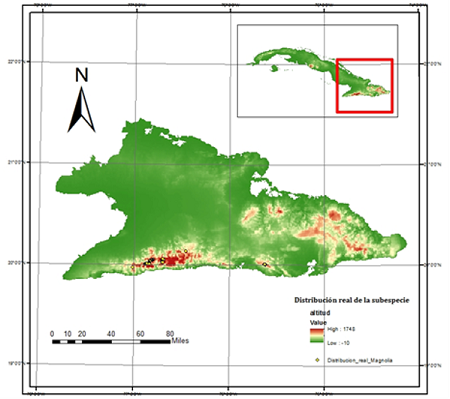
El área de distribución real de M. cubensis subsp. cubensis se determinó a partir de las coordenadas de cada individuo registrados con GPS Gramin. Los individuos adultos censados cuentan con una edad que oscila entre los 12 y los 60 años de edad estimada de acuerdo al fenotipo. El período de recopilación de los puntos de presencia comprende los años entre 2008 y 2017. Con estos datos se generó el mapa de distribución real de la subespecie mediante el empleo del programa ArcGis versión 10.3. Se calculó el área de ocupación real de la población, multiplicando la cantidad de celdas con presencia de al menos un individuo por el área de la celda (400 m²).
Transformación y conversión de variables bioclimáticas y topográficas
Para el estudio se emplearon 19 variables bioclimáticas: (bio01) temperatura media anual, (bio02) rango medio diurno de temperatura, (bio03) isotermalidad, (bio04) estacionalidad de la temperatura, (bio05) temperatura máxima del período más cálido, (bio6) temperatura mínima del período más frío, (bio07) variación anual de temperatura (bio05-bio06), (bio08) temperatura media del trimestre más lluvioso, (bio09) temperatura media del trimestre más seco, (bio10) temperatura media del trimestre más cálido, (bio11) temperatura media del trimestre más frío, (bio12) precipitación anual, (bio13) precipitación del período más lluvioso, (bio14) precipitación del período más seco, (bio15) estacionalidad de las precipitaciones, (bio16) precipitación en el trimestre más húmedo, (bio17) precipitación del trimestre más seco, (bio18) precipitación del trimestre más cálido y (bio19) precipitación del trimestre más frío, obtenidas de la base de datos global de superficies climáticas WorldClim, (http://www.worldclim.org). Además, se utilizó la altitud como variable, a partir del modelo digital de elevación disponible en la colección WordClim basada en los registros del radar SRTM (Shuttle Radar Topography Mission).
Primeramente, los archivos raster de las variables bioclimáticas fueron recortados para tomar el área de estudio. Para esto se utilizó una máscara proveniente de un shapefile de la región oriental de Cuba, el shapefile tuvo que reproyectarse al sistema de coordenadas WGS 84 UTM Zone 18N, una vez reproyectado, constituyó el soporte para la transformación del resto de las variables bioclimáticas y altitud.
La configuración de los raster ambientales se llevó a cabo con la aplicación de la herramienta Entornos Ambientales, que permite que todas las capas ambientales tengan la misma extensión, tamaño de celda y sistema de coordenadas; estos son requisitos indispensables para que los archivos sean procesados por el programa MaxEnt. Una vez configurados los raster fueron convertidos a formato ASCII para poder ser ejecutados por MaxEnt, el proceso de conversión se realizó con el empleo del programa ArcGis 10.3.
Modelo de la distribución
Modelo de la distribución potencial actual
Se modeló la distribución de M. cubensis subsp. cubensis utilizando el programa MaxEnt v. 3.3.3 (Phillips et al., 2006). El resultado de la modelación con MaxEnt, revela la probabilidad relativa de distribución de una especie en todas las cuadrículas o celdas en el espacio geográfico definido, en el cual un valor de alta probabilidad asociado a una cuadrícula particular indica la probabilidad de la misma, de tener condiciones ambientales favorables para la especie modelada (Elith y Leathwick, 2009).
Para la modelación se partió de las coordenadas de los 589 puntos de presencia, puntos generados durante el censo a los individuos de M. cubensis subsp. cubensis, que comprenden todo el área de distribución natural. Las variables utilizadas en esta primera modelación fueron las 19 variables ambientales y la altitud como variable topográfica.
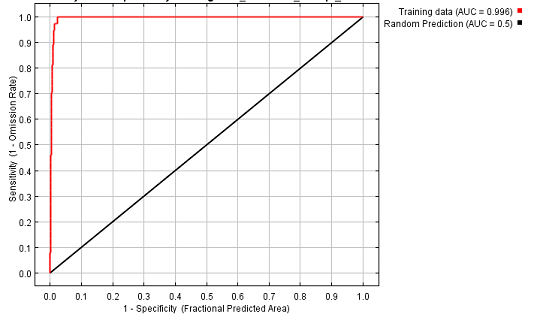
La robustez de predicción del modelo se determinó a través de la técnica de evaluación de la curva operacional (ROC) a través del área bajo la curva ROC (AUC) generando un análisis de sensibilidad vs. Especificidad (Phillips et al., 2006), para esto se utilizó un 75 % de las accesiones de entrenamiento y un 25 % de prueba en la opción random test percentage, la cual le indica al programa que debe escoger de manera aleatoria y apartar el 25 % de los registros de muestras para usarlos en la prueba para la comprobación de los modelos resultantes. El AUC se puede entonces interpretar como la mayor probabilidad de que un punto de presencia seleccionado aleatoriamente esté ubicado en una celda del raster con un alto valor de probabilidad para la presencia de la especie en comparación al de un punto generado aleatoriamente (Elith et al., 2011 y Ávila-Coria et al., 2014). La mayor capacidad de predecir de un modelo generado por MaxEnt se alcanza cuando el AUC tiene un valor de 1 (Araújo y Guisan, 2006).
En la configuración, se especificó un máximo de 500 interacciones con un límite de convergencia prefijado en 0,00001 que garantiza la convergencia del algoritmo, es decir que mantiene una estabilidad en sus análisis (Elith et al., 2011). La definición de estos parámetros del límite de convergencia son usualmente los empleados (Ávila et al., 2014). Con el fin de evitar datos de presencia con registros duplicados se activó borrar duplicados, lo que permitió reducir los sesgos del muestreo.
Para validar y entrenar al mismo tiempo sin que se produzca sobreajuste en la red neural se utilizó la opción crossvalidate. Se estableció como formato de salida la función logística, por ser la más sencilla de conceptualizar, ya que proporciona un estimado entre cero y uno de probabilidad de presencia (Phillips y Dudik, 2010). Esta representa la probabilidad estimada de presencia de la especie, dadas las restricciones impuestas por las variables predictivas. Con este formato, las cuadrículas con valores bajos (cercanos a cero) se consideran inapropiadas para la especie dado su nicho ecológico (Phillips y Dudik, 2010; Elith et al., 2011). El modelo final de la distribución potencial se tomó a partir del mapa en formato ASCII generado por el programa, este mapa fue convertido a formato raster mediante el empleo del programa ArcGis 10.3. Se determinó a su vez, el área que abarca la distribución potencial actual, mediante la creación de polígonos y determinando el área que ocupa dentro del territorio, para ello se utilizaron las herramientas de ArcMap.
Para evaluar la contribución de las variables bioclimáticas dentro del modelo se utilizó el método de remuestreo Jackknife. Esta prueba analiza la contribución de cada variable de manera aislada y adicionalmente genera otro con todas las variables, en donde indica la ganancia de cada una de estas. Para realizar el análisis de la influencia de cada variable climática en la presencia de la especie fueron generadas las curvas de respuesta, ya que estas expresan cómo afectan cada una de estas la predicción de MaxEnt.
Evaluación de zonas de conservación de la subespecie
La determinación de las zonas potenciales y críticas para la conservación de la subespecie, se propusieron mediante la unión de los mapas de distribución geográfica generados por el programa MaxEnt (puntos de presencia y distribución potencial) con la zonificación del Sistema Nacional de Áreas Protegidas (SNAP) de acuerdo al CNAP (2014). Para ello, se utilizó la versión digital del mapa del Centro Nacional de Áreas Protegidas (CNAP) para el período 2014-2020. Se cuantificó el área de las zonas de alta idoneidad para la presencia de la subespecie en cada una de las áreas protegidas objeto de estudio con el programa ArGis 10.3.
RESULTADOS Y DISCUSIÓN
Distribución geográfica
Distribución real
Magnolia cubensis subsp. cubensis ocupa un área de 0,02 km², de ellos 0,0184 km² corresponden a la subpoblación localizada en la provincia Granma y 0,0016 km² a la subpoblación de la provincia Santiago de Cuba (Figura 1). Desde el punto de vista biológico, la distribución pudiera estar relacionada con condiciones ecológicas favorables para su desarrollo solo en las montañas de la Sierra Maestra (Testé et al., 2019).
La disposición actual de su distribución en áreas pequeñas y tan separadas, desde punto de vista geográfico, puede deberse a la fragmentación del hábitat como consecuencia del aprovechamiento del taxón, la poca disponibilidad de individuos reproductores y problemas relacionados con el banco de semillas; sin embargo, estudios que se concentren en estos aspectos son muy escasos (Molina-Pelegrín, 2014). Hasta el momento solo se dispone de un estudio del impacto de la fragmentación del hábitat en la estructura poblacional y genética de M. cubensis subsp. acunae a través de análisis espacial y molecular, que permitió concluir que esta es la causa principal de la reducción de su distribución natural y que, además, incrementa el riesgo ante los escenarios de cambio climático (Hernández et al., 2020). Por lo que estudios moleculares y de autoecología en la subespecie cubensis son necesarios.
Este patrón de distribución en M. cubensis subsp. cubensis y la estructura poblacional de clases diamétricas y de altura evaluada por Molina-Pelegrín et al., (2014) y por Testé et al., (2019) para Gran Piedra, revelaron el envejecimiento de estas poblaciones, lo cual podría explicar la reducida área de ocupación actual.
La distribución geográfica de los individuos de la subespecie comprende desde los 19°58' 10,2'' N y 77°00'29,6'' W hasta los 20°0'46,0'' N y 75°37'50,1'' W. El rango altitudinal se encuentra entre los 702 y los 1 396 m s.n.m, aunque mayor parte de los individuos se encuentran en el rango de 1 000 hasta 1 200 m s.n.m.
Modelo de distribución potencial actual
Existe un buen ajuste para predecir la presencia de la especie, ya que el valor del área bajo la curva ROC fue de 0,996 (Figura 2), valores por encima de 0,9 son considerados como indicadores de buena robustez de predicción del modelo (Phillips et al., 2008 y Elith et al., 2011). Este resultado fue similar en estudios de predicción de hábitat potencial de coníferas (Contreras-Medina et al., 2010; García-Aranda et al., 2012, Ávila-Coria et al., 2014 y Miranda-Sierra 2017) y otras especies de interés forestal (Smith et al., 2012) . Sin embargo, en especies de animales se reportan valores inferiores de AUC en la modelación con variables climáticas, al parecer por la influencia de otras variables ambientales en la definición de su nicho climático (Azor-Hernández y Barro-Cañamero 2014; Cobos-Cobos, 2016).
 Nota: en rojo la curva ROC promedio para los datos de entrenamiento. La línea negra representa la predicción aleatoria (AUC=0,5)
Nota: en rojo la curva ROC promedio para los datos de entrenamiento. La línea negra representa la predicción aleatoria (AUC=0,5)Fig. 2. - Curva de respuesta operativa (ROC) del modelo de distribución potencial actual de Magnolia cubensis subsp. cubensis en el oriente de Cuba
El mapa de probabilidad de presencia potencial, según el umbral elegido, indicó ausencias de la subespecie por debajo de 0,15. El modelo destacó dos núcleos geográficos principales: Sierra Maestra (provincia Granma) y Gran Piedra, también perteneciente al mismo macizo montañoso, pero este último ubicado en la provincia de Santiago de Cuba. Aunque se muestran seis pequeños núcleos poblacionales en el macizo montañoso Sagua Nipe Baracoa, con posibilidades de presencia del taxón. De forma general, se manifiesta un área con probabilidad media (0,38-0,69) de presencia y se reflejó un patrón disyunto en la distribución de la subespecie (Figura 3). La región de Sierra Maestra, en la parte oeste (provincia Granma), comprende la mayor área de ocupación potencial con 54 081,48 ha (92,80 %), mientras Gran Piedra (este) comprende 4 168, 36 ha (7,15 %), para un total de 58 249, 83 ha.
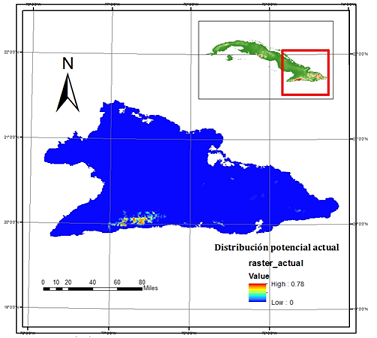
Fig. 3. - Mapa de distribución potencial actual de Magnolia cubensis subsp. cubensis en el oriente de Cuba
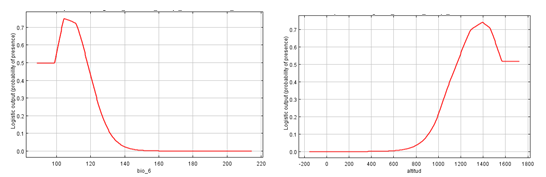
El análisis de la importancia de cada variable mediante la prueba de jackknife (Figura 4), muestra que las variables que más influyen en la presencia de la especie, son las relacionadas con las temperaturas (bio01, bio05, bio06, bio08, bio09 y bio10) y la altitud. Estos resultados permiten identificar las exigencias climáticas de las especies y sus rangos que determinan la probabilidad de presencia. Por lo tanto, este constituye un primer acercamiento a la evaluación de la influencia de las variables climáticas en la distribución de la especie. Asimismo, esta ha sido una alternativa empleada por varios autores para algunas especies endémicas o en peligro de extinción y de esta forma facilitar la toma de decisiones en la conservación como: Cedrela odorata(Gómez-Díaz et al., 2007); Taxus globosa(García-Aranda et al., 2012); Pinus strobus(Joyce y Rehfeldt, 2013); Pinus herrerae(Ávila-Coria et al., 2014); Magnolia schiedeana (Vásquez-Morales et al., 2014) y otras (ver revisión Gray y Hamann 2012).
En este caso, se explica que las variables relacionadas con el régimen térmico son las que afectan, en primer orden, la distribución potencial del taxón. Por tanto, si hay cambios bruscos y prolongados en los rangos para una o varias variables climáticas entonces se vería afectada las condiciones climáticas favorables para la presencia del taxón. Al respecto, McLean (2015) proyecta un clima más seco e incrementos de las temperaturas en el área del Caribe, en particular al oriente de Cuba; esto podría suponer afectaciones en la especie relacionados con sus procesos fisiológicos, la producción de biomasa e inclusive la floración y producción de semilla (Álvarez-Brito, 2017 y Sáenz-Romero, 2014). Esto hará que estas sean sensibles como resultado de alteraciones en su nicho climático.
Estudios de la influencia de los cambios climáticos en procesos fisiológicos y reproductivos ha sido estudiado en varias coníferas y especies latifolias con amplia distribución (Aitken et al., 2008), pero en especies tropicales e insulares son muy escasos (Álvarez-Brito et al., 2014 y Álvarez-Brito, 2018) y estos apoyado además en estudios genéticos de la especie y sus poblaciones son la base para promover la resiliencia y el manejo adaptativo. En Cuba, dichos estudios, aunque incipientes, se dispone de información de alteraciones de las fenofases vegetativas y reproductivas en tres especies forestales (Hechavarría-Kindelán et al., 2008; Hechavarría-Kindelán 2012 y Álvarez-Brito y Mercadet-Portillo, 2014). Por lo que, investigaciones sobre la fenología y biología reproductivas en especies de árboles y sus cambios por efecto de las variaciones en el clima, junto a estos, pero a partir de la simulación de cambios en las variables climáticas en condiciones de viveros, áreas de conservación in situ son cruciales en la comprensión del impacto de los cambios en la estructura demográfica de las especies.
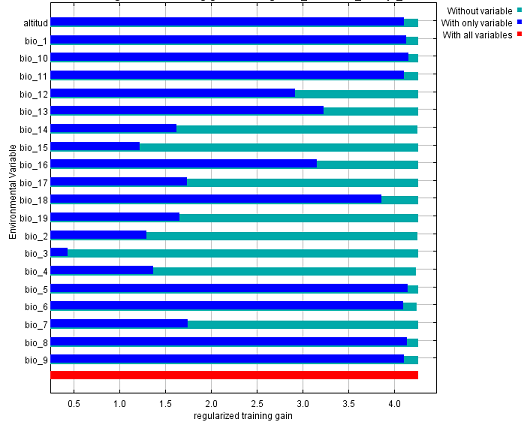
Fig. 4. - Contribución de las variables ambientales al modelo de distribución potencial actual de Magnolia cubensis subsp. cubensis en el oriente de Cuba. Las barras azules representan la ganancia promedio del modelo usando solo una variable y las verdes la ganancia cuando cada variable fue excluida
En el análisis de la respuesta de las variables que más contribuyen al modelo reveló que, en especial, la curva de respuesta de la variable temperatura mínima del período más frío (°C) tuvo una forma exponencial inversa, a menores de temperatura correspondió a aumentos en la probabilidad de presencia de la especie, es aumentos de la idoneidad para la presencia de la subespecie (Figura 5)A. Para el caso de la variable altitud, se muestra que la probabilidad de presencia del taxón aumentó desde los 700 hasta alrededor de los 1 400 m s.n.m, a partir del cual comenzó a perder relevancia (Figura 5)B. La implicación de la altitud como una de las variables más explicativas en los modelos concuerda con lo reportado por González-Torres et al., (2013) y Palmarola et al., (2018) para la subespecie M. cubensis subsp. acunae en el macizo de Guamuhaya y con los estudios de M. schiedeana en México (Vásquez-Morales et al., 2014).
Zonas potenciales y críticas para la conservación de la subespecie
El 78,58 % de la distribución potencial actual de M. cubensis subsp. cubensis se encuentra bajo objeto de conservación de áreas pertenecientes al Sistema Nacional de Áreas Protegidas de Cuba (SNAP), con una superficie de 45 773, 38 ha. A continuación, se muestran los mapas de las zonas potenciales para la conservación in situ del taxón (Figura 6, Figura 7, Figura 8, Figura 9 y Figura 10) y que corresponden a cada una de las áreas protegidas.

Fig. 6. - Mapa de zonas potenciales para la conservación in situ de Magnolia cubensis subsp. cubensis en el Parque Nacional La Bayamesa, basado en la distribución potencial
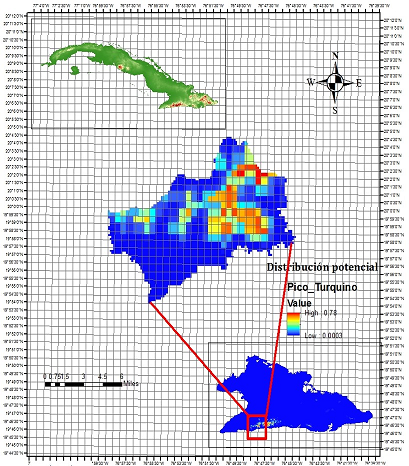
Fig. 7. - Mapa de zonas potenciales para la conservación in situ de Magnolia cubensis subsp. cubensis en el Parque Nacional Turquino, basado en la distribución potencial

Fig. 8. - Mapa de zonas potenciales para la conservación in situ de Magnolia cubensis subsp. cubensis en la Reserva Ecológica El Gigante, basado en la distribución potencial
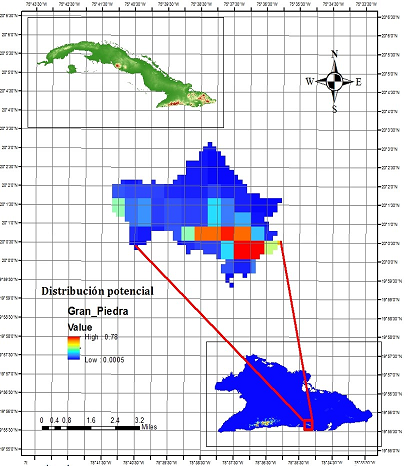
Fig. 9. - Mapa de zonas potenciales para la conservación in situ de Magnolia cubensis subsp. cubensis en el Paisaje Natural Protegido Gran Piedra, basado en la distribución potencial
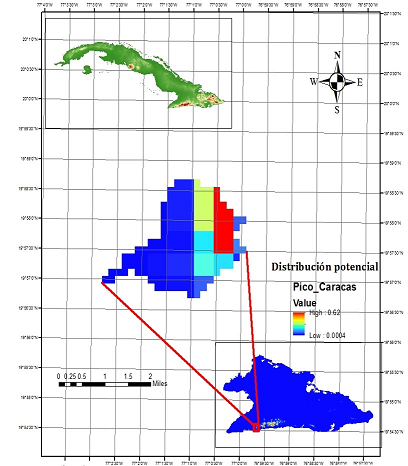
Fig. 10. - Mapa de zonas potenciales para la conservación in situ de Magnolia cubensis subsp. cubensis en la Reserva Ecológica Pico Caracas, basado en la distribución potencial
Se muestra una baja correspondencia de la distribución potencial de M. cubensis subsp. cubensis en el SNAP que contrasta con una alta representatividad real de la subespecie en el estudio desarrollado.
Se muestra una baja correspondencia en la distribución de taxón censada por el SNAP y la presentada en este estudio como distribución actual real, lo que demuestra que no toda la especie está únicamente dentro de las áreas protegidas, y que, por tanto, quedan aún individuos formando parte de espacios donde no existe protección. En este sentido, Hernández y Estrada (2004) plantean que la delimitación de las áreas protegidas se ha establecido en base a razones logísticas, a partir de criterios antropocéntricos o geográficos y sin tomar en cuenta los requerimientos de las especies a proteger. De igual manera, ocurre con relación al área potencial y su correspondiente distribución real, en casi todas las áreas protegidas hay hábitats potenciales donde la especie podría desarrollarse de hacerse acciones para su propagación.
Las zonas críticas de conservación de la subespecie, principalmente se encuentran en la zona de amortiguamiento de las áreas protegidas; esto refuerza la importancia de estas zonas dentro de la estrategia de conservación del taxón y la necesidad de aunar acciones más interdisciplinarias e interadministrativas de las áreas protegidas. Se determinó la existencia de 12 428,62 ha que no forman parte del objeto de conservación de la subespecie y que muestran sitios con probabilidades de presencia del taxón (Figura 11). Algunos de estos sitios están contenidos dentro de la zona de amortiguamiento del Parque Nacional Pico La Bayamesa, lugar donde además se encuentran los mayores asentamientos poblacionales rurales, factor que constituye un riesgo para la conservación in situ de la subespecie.

CONCLUSIONES
Magnolia cubensis subsp. cubensis se distribuye de forma fragmentada en el macizo montañoso Sierra Maestra. Los individuos mayoritariamente se distribuyen en el rango altitudinal comprendido desde los 1 000 hasta los 1 200 m s.n.m.
El área de distribución potencial actual, en el oriente de Cuba, presenta un patrón disyunto con un área de idoneidad total en el macizo montañoso Sierra Maestra, condicionada fundamentalmente por las variables: temperatura mínima del período más frío y elevación sobre el nivel del mar.
El 78,58 % de la distribución potencial actual de Magnolia cubensis subsp. cubensis se encuentra bajo objeto de conservación por parte del Sistema Nacional de Áreas Protegidas de Cuba (SNAP).

