










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkCultivos Tropicales
Print version ISSN 0258-5936
cultrop vol.33 no.1 La Habana Jan.-Mar. 2012
EFECTOS DEL ALUMINIO EN LA DIVISIÓN Y EL ALARGAMIENTOCELULAR EN PLÁNTULAS DE ARROZ (Oryza sativa L.)
Aluminium effects on cell division and elongation in rice seedlings
(Oryza sativa L.)
Dra.C. Idioleidys Alvarez Bello,I Dra.C. Inés Reynaldo Escobar,II Dra.C. Pilar Sánchez Testillano,III Dra.C. María del C. Risueño IV
I Investigador Agregado del Instituto Nacional de Ciencias Agrícolas, gavetapostal 1, CP 32700, San José de Las Lajas, Mayabeque, Cuba.
II Investigador Titular del departamento de Fisiologíay Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas, gavetapostal 1, CP 32700, San José de Las Lajas, Mayabeque, Cuba
III Investigador Titular
IV Profesora de Investigación e Investigador Titular del departamentode Desarrollo de Plantas y Organización Nuclear, Centro de Investigaciones Biológicas del CSIC «Ramiro de Maestzu, 109, 28040, Madrid, España.
RESUMEN
Teniendo en cuenta que la toxicidad por aluminio es el efecto más importante en los suelos ácidos y constituye el mayor factor limitante del crecimiento y la producción vegetal, se desarrolló una investigación que tuvo como objetivo fundamental determinar el efecto del aluminio en los procesos de división y alargamiento celular en dos cultivares comerciales de arroz, así como la relación de éstos con la inhibición del crecimiento radical. Se emplearon diferentes niveles de aluminio para imponer la condición de estrés y los ápices radiculares fueron sometidos al procesamiento tradicional para microscopía óptica. Las secciones teñidas se observaron al microscopio óptico (Zeiss) con cámara digital acoplada y las dimensiones celulares se cuantificaron mediante el programa morfométrico ImageJ. Se demostró que el aluminio inhibió el crecimiento radical de las plántulas de arroz, como consecuencia del efecto tóxico del metal. Se observó una reducción del índice mitótico para ambos cultivares con el incremento de la concentración de aluminio y el estudio del alargamiento celular evidenció que las células radicales se acortaron en longitud y aumentaron su grosor. Los resultados demostraron que los desórdenes que provoca el metal en los dos procesos celulares evaluados son los responsables fisiológicos de la inhibición del crecimiento radical.
Palabras clave: aluminio, procesos celulares, arroz.
ABSTRACT
Aluminium toxicity is a very important factor limiting growing and crop productivity in acid soils. The aim of this paper was to study the aluminium effects on division and elongation cell process and their relation with root growing in roots of two rice commercial cultivars. Different AlCl3 levels were employed to impose stress condition. Root apical segments were processed by Traditional Procedure to Optic Microscopy. The stained sections were viewed with Optic Microscopy (Zeiss) and photographed with Digital Camera Coupled to microscopy. Morfometric Program ImajeJ was employed to determine cell dimensions. Root elongation was reduced in rice seedlings like a consequence to aluminium toxic effect. A mitotic index inhibition was observed with aluminium level increasing and reduced length accompanied by radial expansion was observed in meristematic cells. The obtained results evidence some changes provokes by aluminium toxic levels in both cellular process which are responsible to cell growing.
Key words: aluminium, cell process, rice.
INTRODUCCIÓN
La acidificación de los suelos se ha intensificado en los últimos años, estimándose que aproximadamente el 30% de la superficie terrestre y el 50% de las tierras cultivadas están afectadas por este fenómeno y de estas últimas cerca de un 60% se localiza en las zonas tropicales y subtropicales (1).
Los suelos de Cuba no han sido de los más afectados por la acidez (2), no obstante, diversos factores naturales y antrópicos han generado alrededor de 557 400 hectáreas de suelos alíticos con el transcurso de los años, los que se encuentran distribuidos en casi toda la isla, con una mayor incidencia en las provincias de Pinar del Río, Camagüey y Granma (3).
La toxicidad por aluminio es el efecto más importante en los suelos ácidos y constituye el mayor factor limitante del crecimiento y la producción vegetal (1, 4, 5). Para la mayoría de las plantas, la raíz es el órgano más afectado por este ión (6), por lo que se han planteado diversas hipótesis acerca de los mecanismos de toxicidad en esta parte de la planta.
En este sentido, se conoce que diferentes componentes y procesos celulares pueden ser afectados por el aluminio (7) y entre los más importantes se encuentran, el núcleo y la división celular (8, 9) y la dinámica del citoesqueleto (10). En un estudio en el cultivo del maíz (11) la reducción del número de células proliferativas inducidas por aluminio estuvo acompañada por un acortamiento en la región de división celular. Varios años después se observó la rápida entrada del ión a las células y su localización cercana a los núcleos celulares, en células radicales de soya y suspensiones celulares de Catharanthus roseaus (12). A partir de estos resultados las investigaciones centraron su atención en el núcleo celular, teniendo en cuenta que el aluminio podía inhibir la división celular mediante su interacción con algunos componentes nucleares y esto permitió sugerir que la toxicidad letal podía estar asociada con esta interacción (13).
En un estudio de suspensiones celulares de dos líneas de tabaco ante una exposición prolongada al aluminio, se evidenció una desorientación de los elementos del citoesqueleto microtubular (4). En este caso los cambios observados estuvieron precedidos por una disminución en la viabilidad celular y un incremento en los niveles de ?-tubulina, hallazgos que permitieron concluir que el citoesqueleto microtubular es un sitio clave ante la toxicidad por aluminio, aunque otras evidencias más convincentes deben obtenerse al respecto. (14).
Aún cuando lo anterior muestra varios estudios sobre el tema en algunas gramíneas, no se han realizado investigaciones para cuantificar estos dos procesos celulares en la raíz de arroz, lo que ha limitado el estudio integrado de los cambios morfológicos y fisiológicos que puede provocar el aluminio en esta planta.
Por las razones antes expuestas, el objetivo de la investigación fue determinar el efecto del aluminio en los procesos de división y alargamiento celular en dos cultivares comerciales de arroz, así como la relación de éstos con la inhibición del crecimiento radical.
Materiales y Métodos
Material vegetal
Los experimentos se realizaron en el Laboratorio de Organización y desarrollo Nuclear del Centro de Investigaciones Biológicas (CIB), España y en el Departamento de Fisiología y Bioquímica Vegetal del Instituto Nacional de Ciencias Agrícolas (INCA). Para ello se emplearon plántulas de arroz (Oryza sativa L) de dos cultivares comerciales cubanos.
El cultivar Jucarito-104 (J-104), que es empleada como patrón comercial por sus potencialidades en el rendimiento (15) y la variedad INCA LP- 7, obtenida en el Departamento de Genética y Mejoramiento de las Plantas del Instituto Nacional de Ciencias Agrícolas, la cual exhibe cierto grado de tolerancia al estrés iónico (16).
Condiciones experimentales
Se desarrolló un experimento en el que se definieron 4 tratamientos, en los cuales el nivel de aluminio en el medio de germinación fue de 0 (Control), 30, 65 y 125 µmol.L-1 de cloruro de aluminio (AlCl3), respectivamente. Para ello las semillas de arroz se desinfectaron con una solución de hipoclorito de sodio comercial al 10% durante 15 minutos y se lavaron tres veces con agua esterilizada. Posteriormente se colocaron en placas de Petri (90x15 mm) con papel de filtro, el que se humedeció diariamente con 1ml de la solución de AlCl3 empleada para la germinación de las semillas, en cada uno de los tratamientos, y con agua, para el control.
Se colocaron 25 semillas en cada placa de Petri, con 12 horas de luz natural y 12 horas de oscuridad y una temperatura noche/día de 21/26 o C.
Se realizaron 3 réplicas del experimento y las plántulas se evaluaron después de siete días de germinadas las semillas. Se muestrearon 20 plántulas por placa y un total de 60 por tratamiento, a las cuales se les tomó la radícula para realizar las evaluaciones correspondientes.
Procesamiento para microscopía
Para el estudio de las secciones longitudinales de la radícula, las muestras se sometieron al procesamiento tradicional para microscopía óptica (2) y se incluyeron en resina Lowicryl K4M. Los bloques de resina fueron seccionados en un ultramicrótomo y se obtuvieron secciones longitudinales de la radícula de una micra. Para lograr un adecuado contraste de las muestras se utilizó el azul de toluidina y las observaciones se realizaron en un microscopio óptico de luz (Zeiss) con una magnificación de 1000x.
Evaluación del efecto del Al en la orientación del crecimiento celular
Para evaluar el alargamiento celular, se procedió a determinar el largo y ancho de las células del meristemo fundamental en cada uno de los tratamientos. Para ello se obtuvieron imágenes digitales de las células del meristemo fundamental de esta zona, con una ampliación de 1000x, las que se exportaron al Programa Morfométrico ImageJ, donde se utilizó el instrumento de “medición lineal”. El largo de las células corticales se determinó en todos los casos, de forma paralela al borde de la radícula y para el grosor celular de forma perpendicular, siempre por el centro de las células, lo que permitió homogenizar las mediciones. Se tomaron tantas imágenes como fue necesario para medir 200 células por tratamiento.
Evaluación del efecto del Al en el proceso de división celular
Se cuantificó el índice de proliferación celular a través del cálculo del índice mitótico y para ello se empleó la técnica de tinción con orceína acético-clorhídrica (17), con una modificación, ya que para lograr la penetración de la orceína en los ápices radiculares fueron necesarios tres calentamientos en vez de uno.
El número de células estadísticamente representativo se estimó de acuerdo a la fórmula empleada por Matos y Molina (1997): i = Z2a/2 x S2/e2, donde Z tiene un valor de 1,96 según la estadística tabulada de la normal estándar y S2, que significa la varianza muestral, es de 13,66. El error del muestreo (e2) es de 0,25. De acuerdo a esta fórmula, 1462 células por tratamiento es el valor mínimo de células que se deben analizar para obtener resultados confiables y se evaluaron tantos campos como fue necesario para contar 2000 células por tratamiento. Teniendo en cuenta que dentro de cada campo se contaron las células que se encontraban en mitosis (profase, metafase, anafase y telofase) y en interfase.
Los cálculos del índice mitótico se realizaron a través de la ecuación:
IM = M x 100/T, donde:
IM: Índice Mitótico,
M: Células en mitosis (profase, metafase, anafase y telofase) y
T: Total de células contadas (Mitosis + Interfase).
Análisis estadísticos
Se utilizó un diseño completamente aleatorizado. Los resultados experimentales del largo y ancho de las células y el índice mitótico fueron sometidos a un análisis de varianza con arreglo bifactorial, donde los factores fueron las variedades y las concentraciones de Al. En los casos que se encontraron diferencias significativas, se compararon las medias según la Dócima de Tukey, para el 5% de probabilidad del error. Para el análisis estadístico fue utilizado el paquete estadístico STATGRAPHICS Versión 4.1 en ambiente Windows.
RESULTADOS Y DISCUSIÓN
Dentro de los factores abióticos que más influyen en el desarrollo del sistema radical, se pueden mencionar, la temperatura, el contenido de agua y de oxígeno, así como una elevada concentración de elementos químicos, entre los que se encuentran el aluminio (18).
La radícula fue el centro de atención de esta investigación dentro del sistema radical, en primer lugar por la importancia que reviste en la fase de plántula y además porque garantiza una adecuada absorción de agua e iones en esta fase del desarrollo de la planta. Es importante destacar que no se produjo interacción entre las variedades y las concentraciones de aluminio para ninguna de las variables estudiadas.
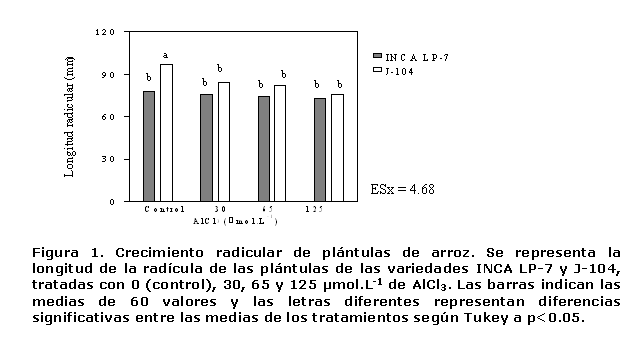
Las plántulas del control en la variedad J-104 exhibieron valores de longitud de la radícula, superiores a las de la INCA LP-7, como se puede apreciar en la Figura 1. Sin embargo, fueron las más afectadas por el ión Al3+, ya que a 30 µmol.L-1 se observó una disminución significativa de este indicador en relación con el control. Esta alteración se considera el síntoma más frecuente ante la presencia de este ión, ya que una reducción similar del crecimiento ha sido observada en diferentes especies de plantas (12, 19, 20) como consecuencia del efecto tóxico del aluminio (21).
Este comportamiento indicó que 30 µmol.L-1 es suficiente para afectar la longitud radicular en la variedad J-104 y que a partir de esta concentración se mantiene el efecto inhibitorio del Al en el crecimiento de la radícula, de manera tal que en las plántulas expuestas a 125 µmol.L-1 se observó una reducción del 23% en relación con las plántulas no tratadas.
En la INCA LP-7 la longitud radicular no difirió entre ninguno de los tratamientos, por lo que esta variedad fue capaz de soportar dosis de hasta 125 µmol.L-1 de AlCl3, sin tener afectaciones en la longitud de su radícula (Figura 1).
El comportamiento diferente que exhibió la longitud de la radícula en las variedades de arroz, puede estar relacionado con el grado de tolerancia al metal de cada una de ellas, aún más, cuando se ha visto que la intensidad de la inhibición puede diferir entre especies y entre variedades (12).El crecimiento radical es el resultado de la división y el alargamiento de las células radicales, razón por la que se evaluaron las afectaciones provocadas por el ión Al3+ en ambos procesos celulares.
Al evaluar el efecto provocado por el aluminio en el alargamiento celular, se pudo apreciar que en las plántulas del control, las células del meristemo fundamental se caracterizaron por tener una estructura poliédrica alargada (Figura 2A y B), sin embargo en las expuestas al metal, éstas se deformaron y exhibieron paredes muy irregulares, así como una expansión radial en su estructura (Figura 2C-H).
A través de un estudio morfométrico se pudieron cuantificar las alteraciones observadas en la estructura de estas células y se demostró que la toxicidad del ión Al3+ afecta de forma significativa el alargamiento celular.
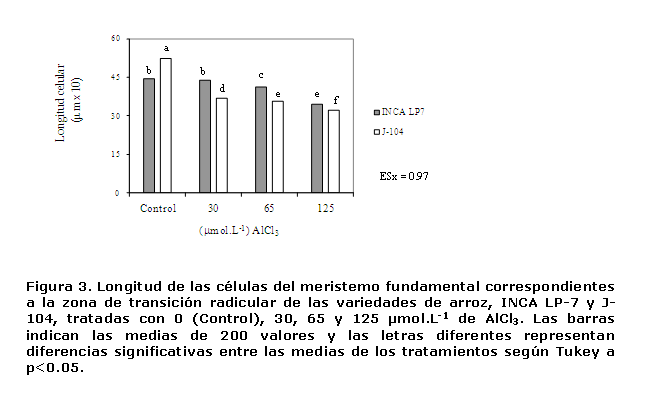
Las plántulas de la variedad INCA LP-7 no exhibieron cambios en la longitud de las células cuando fueron expuestas a 30 µmol.L-1 de AlCl3, sin embargo, en la J-104 a ese mismo valor, se observó una disminución en la misma (Figura 3). Esto indica que la concentración de 30 µmol.L-1 resulta tóxica para esta última variedad, resultados que apoyan los obtenidos al analizar la longitud de la radícula (Figura 1).
Además, cuando las plántulas de la variedad J-104 fueron expuestas a la mayor dosis de aluminio (125 µmol.L-1), la longitud celular se redujo casi un 40 % en relación con el control, mientras en que en la INCA LP-7 la reducción fue de solo un 25.5% (Figura 3), lo que evidenció que la longitud de sus células, fue menos afectada ante la toxicidad del aluminio.
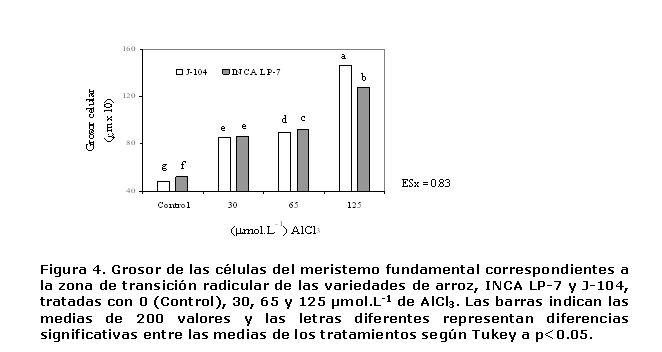
El grosor de las células del meristemo fundamental determinado a través del estudio morfométrico, arrojó un incremento importante ante la presencia del ión Al3+ (Figura 4). Para este indicador, las dos variedades estudiadas exhibieron un nivel de tolerancia similar, ya que ambas mostraron un aumento significativo del grosor celular cuando fueron expuestas a 30 µmol.L-1 del metal. No obstante, los resultado apuntan a que la J-104 fue la más afectada por el estrés iónico, pues el grosor celular se incrementó en un 70%, mientras que para la otra variedad el incremento fue de un 57% (Figura 4).
La evaluación integral del estudio morfométrico, indicó que el aluminio provocó disminución en la longitud de las células del meristemo fundamental e incremento en el grosor de las mismas, de manera que dichas células se hicieron más cortas y anchas en la medida en que se incrementaron los niveles de la exposición al ión metálico, sobre todo en la variedad J-104, la que evidenció éstas modificaciones a menores dosis del metal en relación con la INCA LP-7. Sin lugar a dudas estas modificaciones contribuyen a la reducción detectada en la longitud y al engrosamiento observado en la radícula de las plantas expuestas al metal.
Por otro lado, la respuesta significativa observada tanto en el largo como en el grosor celular de las plántulas expuestas a la menor dosis de aluminio respecto a las del control, sugiere que 30 µmol.L-1 del metal pueden ser suficientes para romper la sensible dinámica de ensamble y desensamble de la red de microtúbulos y microfilamentos. Además, se evidencia la posibilidad de que el citoesqueleto pueda ser un blanco de acción importante ante la toxicidad provocada por el aluminio, como ha sido informado para otras especies (22, 23). Aún más, si se tiene en cuenta que el citoesqueleto es la red tridimensional de proteínas que conforman el citosol, la cual cumple un papel esencial en el mantenimiento de la forma celular entre otras funciones (24).
A partir de estos elementos, la expansión celular que arrojó el estudio morfométrico pudiera ser explicada mediante la interacción del ión Al3+ con factores que influyen en la organización del citoesqueleto, similar a como sucede en Arabidopsis thalianna (22), como los niveles de Mg2+ y Ca2+ en el citosol.
La división celular no es por sí misma un mecanismo de crecimiento, ya que no conduce necesariamente a un incremento en el tamaño; no obstante es un proceso que acompaña al crecimiento en los tejidos meristemáticos. Por este motivo el estudio del efecto del ión Al3+ en la división celular estuvo confinado a la zona de división de la radícula, según la Figura 5 y se evaluó mediante la determinación del índice mitótico (IM), como indicador cuantitativo de este proceso celular.
La caracterización de los núcleos celulares, que se observaron de color rojo, permitió determinar con facilidad en ambas variedades las células que se encontraban en interfase, así como en las diferentes fases de la mitosis (profase, metafase, telofase y anafase) para su posterior evaluación. La misma se realizó a través de la figura mitótica característica de cada fase, como se muestra en la Figura 5 y fue posible por la modificación realizada a la técnica de tinción con orceína, la que permitió un adecuado contraste del material nuclear.
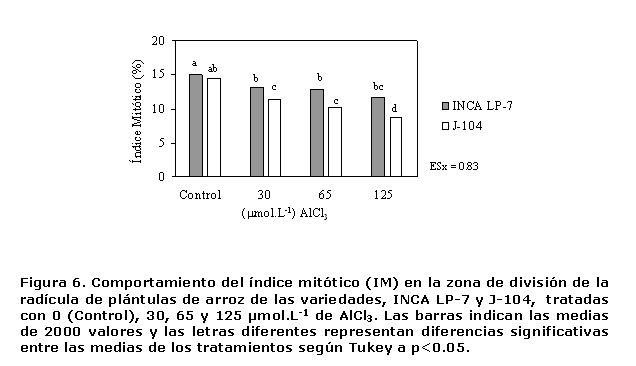
Al evaluar el índice mitótico como indicador del proceso de división celular, se pudo apreciar una disminución progresiva del mismo con el aumento en los niveles de aluminio para la variedad J-104, (Figura 6), lo que se corresponde con la reducción en longitud observada en esta variedad y evidencia las afectaciones que puede provocar este ión en el crecimiento celular, debido a sus efectos tóxicos. Las dos variedades mostraron una reducción de este indicador ante la concentración de 30 µmol.L-1 de AlCl3, sin embargo para los demás tratamientos exhibieron un comportamiento diferente (Figura 6). Lo anterior indica que esta dosis es suficiente para afectar la división de éstas células.
En la variedad INCA LP-7, las dosis de 65 y 125 µmol.L-1 de AlCl3 no afectaron de forma significativa el índice mitótico, lo que indica que después de una reducción en el mismo, se alcanza una estabilidad en el proceso de división celular.
El comportamiento de la variedad J-104 fue diferente, ya que no se apreciaron diferencias significativas entre las plántulas tratadas con 30 y 65 µmol.L-1 de AlCl3, sin embargo las expuestas a 125 µmol.L-1 evidenciaron una reducción del índice mitótico de un 40.2% en relación con las del control, mientras que en la INCA LP-7 lo hizo solo en un 21.7% (Figura 6). Esto está en total concordancia con la inhibición del alargamiento radical observado en la radícula de las plantas tratadas con aluminio y evidencian el efecto inhibitorio que ejerce este elemento sobre los procesos celulares de alargamiento y división celular.
Los resultados obtenidos evidenciaron que el aluminio provocó una reducción en la longitud radicular y que esta fue causada en primer lugar por la inhibición de la división celular. La evaluación del índice mitótico, como indicador de crecimiento, demostró una afectación importante en este proceso, ya que el porciento de células en la fase mitótica disminuyó ante la presencia del ión, sobre todo, en la variedad J-104. Este comportamiento sugirió la posibilidad de que el aluminio pudiera afectar algún elemento en la interfase que impidiera la entrada de la célula a la fase mitótica.
Por otra parte se comprobaron alteraciones en la orientación del crecimiento celular, fenómeno que estuvo dado por una reducción en longitud y una expansión en sentido radial de las células del meristemo fundamental. Se evidenció la posibilidad de que el aluminio pudiera afectar elementos del citoesqueleto y alterar así, la polaridad celular, aspectos que deben ser abordados en estudios posteriores.
REFERENCIAS
1. Pereira, L.M.; Tabaldi, L.A.; Gonçalves, J.F.; Jucoski, G.O.; Pauletto, M.M.; Weis, S.N.; Nicoloso, F.T.; Borher, D.; Rocha, J.B.T.; Schetinger, M.R.C. Effect of aluminium on d-aminolevulinic acid dehydratase (ALA-D) and the development of cucumber (Cucumis sativus).Environ. Exp. Bot. 2006, 57,106–115.
2. Alvarez, I.; Sam, O. y Reynaldo, I. Cambios inducidos por el aluminio en la morfología radicular del arroz. Cultivos Tropicales. 2005, 26, 21-25.
3. Cuba. MINAGRI. CAI Arrocero “Los Palacios”. Informe de los resultados de la campaña de arroz de frío 2008-2009. 2009, 18 p.
4. Schwarzerová, K.; Zlenkova, S.; Nick, P.; Opatrn, Z. Aluminium induced rapid changes in the microtubular cytoskeleton of tobacco cell lines. Plant and Cell Physiol. 2002,43, 207-216.
5. Liu, Q.; Yang, J.L.; He, L.S.; Li, Y.; Zheng, S.J. Effect of aluminium on cell wall, plasma membrane, antioxidants and root elongation in triticale. Biologia Plantarum. 2008, 52, 87-92.
6. Wang, J.W.; Kao, C.H. Effect of aluminium on endosperm reserve mobilization in germinating rice grains. Biologia Plantarum. 2007, 49, 405-409.
7. Morita, A.; Yanagisawa, O.; Takatsu, S.; Maeda, S.; Hiradate, S. Mechanism for the detoxication of aluminum in roots of tea plant (Camellia sinensis (L.) Kuntze). Phytochemistry. 2008, 69, 147–153.
8. Panda, S.K.; Matsumoto, H. Molecular physiology of aluminum toxicity and tolerance in plants. Bot Rev. 2007, 73, 326–347.
9. Panda, S.K.; Baluska, F.; Matsumoto, H. Aluminum stress signaling in plants. Plant Signal Behav. 2009, 4, 592–597.
10. Shahk, J., Nongkynrih, J.M. Metal hyperaccumulation and bioremediation. Biologia Plantarum. 2007, 51, 618-634.
11. Ryan, P.R.; Delhaize, E.; Randall, P.J. Malate efflux from root apices: evidence for a general mechanism of Al-tolerance in wheat. Aust. J. Plant Physiol. 1995, 22, 531-536.
12. Matsumoto, H. Cell biology of aluminium toxicity and tolerance in higher plants. Int. Rev. Cytol. 2000, 200, 1-46.
13. Amenós, M.; Corrales, I.; Poschenrieder, C.; Illéš, P.; Baluška, F.; Barceló, J. Different effects of aluminium on the actin cytoskeleton and brefeldin A-sensitive vesicle recycling in root apex cells of two maize varieties differing in root elongation rate and Al tolerance. Plant Cell Physiol. 2009, 50, 528–540.
14. Zheng, S.J.; Yang, J.L. Target sites of aluminium phytotoxicity. Biologia Plantarum. 2005, 49, 321-331.
15. Díaz, S.; Morejón, R. Comportamiento de variedades de arroz de diferente procedencia en la localidad de Los Palacios. Cultivos Tropicales. 2002, 23, 63-67.
16. González, M.C.; Cristo, E.; Pérez, N.; Delgado, P. INCA LP-7, nueva variedad de arroz para suelos afectados por la salinidad. Cultivos Tropicales. 2002, 23, 89.
17. Matos, A.; Molina, J. Estudio citogenético de células radicales de Aloe vera L. Rev. Fac. Agron. 1997, 14, 173-182.
18. Liao, H.; Wan, H.; Shaff, J.; Wang, X.; Yan, X.; Kochian, L.V. Phosphorus and Aluminum Interactions in Soybean in Relation to Aluminum Tolerance. Exudation of Specific Organic Acids from Different Regions of the Intact Root System. Plant Physiology. 2006, 141, 674–684.
19. Ciamporová, M. Morphological and structural responses of plant roots to aluminium at organ, tissue and cellular levels. Biol. Plant. 2002, 45, 161-171.
20. Tahara, K.; Norisada, M.; Yamanoshita, T.; Kojima, K. Role of aluminum-binding ligands in aluminium resistance of Eucalyptus camaldulensis and Melaleuca cajuputi. Plant Soil. 2008,302, 175–187.
21. Wang, J.W.; Kao, C.H. Protective effect of ascorbic acid and glutathione on AlCl3-inhibited growth of rice roots. Biologia Plantarum. 2007, 51, 493-500.
22. Baskin, T.I., Wilson, J.E. Inhibitor of protein kinase and phosphatases alter root morphology and disorganize cortical microtubules. Plant Physiol. 1997,113, 493-502.
23. Tsai, T.M.; Huang, H.J. Effects of iron excess on cell viability and mitogen-activated protein kinase activation in rice roots. Physiologia Plantarum. 2006, 127, 583–592.
24. Taiz, L.; Zeiger, E. Plant physiology. (Eds) Sinaver Associates, 2006. 672 p.
Recibido 12/10/2010, aceptado 13/12/2011.
Dra.C. Idioleidys Alvarez Bello, Investigador Agregado del Instituto Nacional de Ciencias Agrícolas, gavetapostal 1, CP 32700, San José de Las Lajas, Mayabeque, Cuba. Email: ialvare@inca.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}