










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Cubana de Medicina Tropical
On-line version ISSN 1561-3054
Rev Cubana Med Trop vol.59 no.3 Ciudad de la Habana Sept.-Dec. 2007
ARTÍCULO ORIGINAL
Resistencia a insecticidas y mecanismos de resistencia en Aedes aegypti (Diptera: Culicidae) de 2 provincias del Perú
Insecticide resistance mechanisms of Aedes aegypti (Diptera: Culicidae) from two Peruvian provinces
Lic. Juan A. BissetI; Lic. M. Magdalena RodríguezII; Téc. Ditter FernándezIII; Lic. Miriam. PalominoIV
I Doctor en Ciencias. Licenciado en Ciencias Biológicas. Investigador Titular. Instituto de Medicina Tropical ¨Pedro Kourí¨(IPK).
II Licenciada en Bioquímica. Investigadora Auxiliar. IPK.
III Técnica en Química. IPK.
IV Licenciada en Biología. Instituto Nacional de Salud. Perú.
RESUMEN
Se realizó un estudio de la resistencia a insecticidas en larvas y adultos de Aedes aegypti de 2 provincias del Perú: Trujillo y Tumbes donde prevalecen altos índices de infestación de este vector y están sometidas a extenso uso de insecticidas por la Estrategia de Vigilancia y Control Vectorial del Ministerio de Salud. Los bioensayos en larvas revelaron susceptibilidad al insecticida organofosforado malation (FR50 ≤ 5x) en la cepa TRUJILLO, moderada a fention y fenitrotion (FR50 entre 5 y 10) y alta resistencia (FR50 ≥ 10x) a clorpirifos y temefos, sin embargo en la cepa TUMBES se observó susceptibilidad para los organofosforados evaluados, excepto para fention, con moderada resistencia. En el estado adulto, a la dosis recomendada, en TRUJILLO se observó resistencia al organoclorado DDT y a los piretroides lambdacialotrina y ciflutrina, la cepa TUMBES fue resistente a DDT y a todos los piretroides evaluados. Ninguna de las 2 cepas mostró resistencia a clorpirifos en el estado adulto. Los resultados, utilizando sinergistas, demostraron que las enzimas esterasas y monooxigenasas desempeñaron un papel importante en la resistencia observada a organofosforados en larvas de la provincia TRUJILLO. A través de ensayos bioquímicos se encontró a elevada frecuencia la actividad incrementada de esterasas en TRUJILLO, al igual que la glutation transferasa (GST) y la acetilcolinesterasa modificada (AchE). Mediante electroforesis en gel de poliacrilamida se observó en TRUJILLO, no en TUMBES, la prevalencia de la amplificada actividad de esterasas A4.
Palabras clave: Aedes aegypti, insecticidas, esterasas, glutation transferasa, acetilcolinesterasa, Perú.
SUMMARY
Insecticide resistance of Aedes aegypti larvae and adults from two Peruvian provinces, that is, Trujillo and Tumbes provinces, was conducted. High infestation indexes and extensive use of insecticides based on the Vector Surveillance and Control Strategy of the Ministry of Public Health prevailed in these places. Larval bioassays revealed susceptibility to organophosphorate insecticide called malathion in TRUJILLO strain, it being moderate to fention and fenitrotion and high to chlorpyriphos and temephos; however, TUMBES strain was susceptible to the evaluated organophosphorate compounds, except for fention, with moderate resistance. In the adult state, at the recommended dose, TRUJILLO strain showed resistance to DDT organochlorate insecticide and to pyrethoids called lambdacyalotrine and cyflutrine whereas TUMBES was resistent to DDT and to all assessed pyrethoids. None of them was resistent to chlorpiriphos in adult stage. By using synergists, the results showed that esterases and monooxigenases played an important role in the detected resistence to organophosphorate in Aedes larvae from TRUJILLO province. Biochemical assays yielded that increased activity of esterases was very frequent in TRUJILLO strain as was the case of glutathion transferase(GST) and modified acetylcholinesterase (AchR). On the other hand, the polyacrylamide gel electrophoresis allowed observing the prevalence of amplified activity of esterases A4 in TRUJILLO strain but not in TUMBES strain.
Key words: Aedes aegypti, insecticides, esterases, glutathion transferase, acetylcholinesterase, Perú.
INTRODUCCIÓN
Aedes aegypti Linnaeus es el vector más importante en la transmisión de fiebre amarilla, dengue y fiebre hemorrágica del dengue (FHD) en la región de América. En los programas de control de Aedes aegypti, el uso de insecticidas es un componente importante para lograr la reducción de las poblaciones larvales y adultas de este vector. Dentro de los insecticidas organofosforados más utilizados se encuentran el temefos como larvicida en el tratamiento focal, fention, fenitrotion y malation en tratamiento perifocal o en rociado intradomiciliario o espacial. Hoy día se están utilizando generalmente para el control de adultos los insecticidas piretroides.
Para el tratamiento focal en las aguas de consumo humano, la Organización Mundial de la Salud ha brindado otras alternativas aparte del temefos, el metopreno a dosis no superiores de 1 mg de ingrediente activo (ia) por litro (1 ppm), el pyriproxyfeno a dosis hasta 0,01 mg ia por litro (0,01 ppm) y B. thuringiensis israelensis también es recomendado como otra alternativa.1
La resistencia a insecticidas limita el éxito de las operaciones de control y Aedes aegypti es una especie que tiene la habilidad de desarrollar resistencia a insecticidas y ya existen reportes de esta.2-6 También han sido reportados algunos mecanismos bioquímicos responsables de la resistencia a estos químicos.7-10
El presente trabajo hace una evaluación de los niveles de resistencia a insecticidas en Aedes aegypti de 2 provincias del Perú, tanto en larvas como en adultos, a los insecticidas que generalmente se utilizan para su control, así como otros que pueden tenerse como alternativos. Además se determinan los posibles mecanismos responsables de la resistencia.
MÉTODOS
Cepas
Se utilizaron 3 cepas:
TRUJILLO y TUMBES: 2 cepas de Aedes aegypti provenientes de 2 provincias del Perú, colectadas en 2002.
ROCKEFELLER: 1 cepa de referencia de Aedes aegypti susceptible a insecticidas, suministrada por el CDC, San Juan, Puerto Rico.
Insecticidas grado técnico utilizados
Organofosforados: malation, clorpirifos, metil-pirimifos, temefos, fenitrotion y fention.
Piretroides: lambdacialotrina, deltametrina, betacipermetrina y ciflutrina
Carbamato: propoxur.
Bioensayos de larvas
En los bioensayos de la OMS,11 se realizaron 5 réplicas de cada concentración del insecticida (20 larvas por réplica), se registró entre 2 y 98 % de mortalidad. Todas las soluciones se ajustaron a un volumen final de 1 mL con acetona. Esta concentración de acetona no causó mortalidad en los controles.
La lectura de las mortalidades se realizó a las 24 h, se halló la CL50 y la CL90 con el programa probit- log.12
Bioensayos de adultos
Los bioensayos se realizaron siguiendo las normas de la OMS, para mosquitos adultos.11 Las hembras alimentadas con sangre de curiel, fueron expuestas a papeles impregnados con insecticidas, a dosis diagnósticas sugeridas por la OMS o por el fabricante. Cada uno de los insecticidas fue evaluado a través de 4 réplicas, cada una con 25 mosquitos; los papeles impregnados se colocaron en los cilindros plásticos de exposición (kits de la OMS) durante 60 min, a este tiempo se pasaron los mosquitos al tubo control, el cual contenía el papel de filtro, impregnado con el solvente de cada insecticida a ensayar. No se observó mortalidad en los controles, que consistían en papeles impregnados con aceite de silicona. Los resultados fueron analizados sobre la base de criterios de resistencia a insecticidas según la OMS.
La acción de 2 sinergistas, S, S, S tributil fosforotritioato (DEF) y piperonil butóxido (PB) fueron investigados exponiendo las larvas de cuarto estadio a 0,008 mg/L de DEF o 5 mg/L de PB durante 4 h previo a la adición de la solución del insecticida y se determinó la mortalidad después de 24 h de exposición, hallándose las concentraciones letales 50 y 90 a través del programa Probit-logaritmo.10 No existió mortalidad a estas concentraciones del sinergista solo.
Ensayos bioquímicos: se determinó la actividad de esterasas en larvas de tercer estadio tardío o cuarto temprano, de acuerdo con el método estandarizado para Aedes aegypti.13 Se homogenizó cada larva en 200 µL de buffer fosfato 0,01 M, pH 7,5. En una placa de microtitulación de ELISA, a 20 µL del homogenato se le añadió 200 µL del sustrato (0,7-mM de ß-naftil acetato). Después de dejar transcurrir la reacción por 10 min, se le añadió 40 µL de fast blue y se leyó la densidad óptica (DO) a 570 nm en lector de placas de ELISA Labsystems iMS, manufacturado en Finlandia.
La actividad de glutation- S-transferasa (GST) fue determinada de acuerdo con el método de Booth y otros,14 y modificado para Aedes aegypti. 13 A 20 µL de cada homogenato de larva se le añadió 250 µL de una mezcla de reacción de 1-cloro-2, 4 dinitrobenzeno 50 mM; y de glutation reducido 20 mM. Se dejó transcurrir la reacción por 3 min y se leyó la DO a 340 nm. Se calculó la actividad específica de GST, después de calcular la concentración de proteínas en cada homogenato y se expresó como µmol/mg.min.
Un estimado de la frecuencia de los mecanismos de esterasas y GST fue calculado a partir del número de individuos susceptibles para cada ensayo, asumiendo que la población se encontraba en equilibrio de Hardy- Weinberg.
Se realizó electroforesis en gel de poliacrilamida (10 %). En tubos Eppendorf (1,5 mL), se adicionaron 10 µL de muestra más 10 µL del indicador xilene cianol (0,02 % en sacarosa a 15 %). Se aplicaron 20 µL de esta mezcla en el gel y se realizó la corrida a 150 V, durante 45 min. Para la tinción de las bandas de esterasas, se sumergieron los geles en 50 mL de buffer fosfato (0,1 M) que contenía 4 mL de cada uno de los sustratos inespecíficos de las esterasas (ß-naftilacetato). Después se añadieron 0,5 g del colorante fast blue RR, disuelto previamente en agua destilada y SDS (sodio duodecil sulfato) 5 %. Para fijar la coloración de las bandas se sumergió el gel en una solución de ácido acético 10 %. A cada una de las bandas se le determinó la movilidad relativa.
RESULTADOS
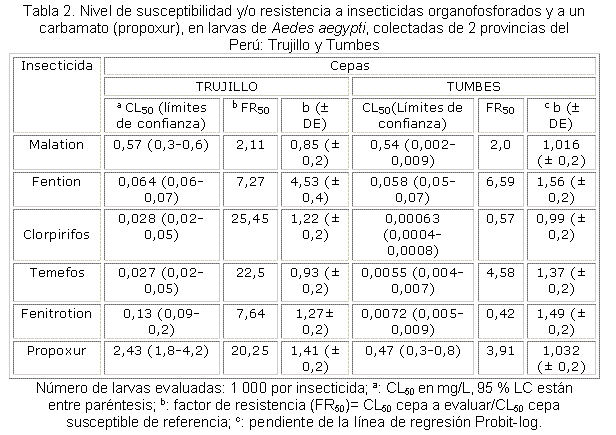
En la tabla 1 se muestran los valores de concentración que causan 50 % (CL50) y 90 % (CL90) de mortalidad en la cepa susceptible de referencia ROCKEFELLER. En la misma tabla se muestran también las pendientes (b) de cada una de las líneas de regresión probit-log. Como se observa, los valores de las pendientes son elevados, lo cual indica que es una cepa homogénea para la susceptibilidad, es decir a mayor pendiente de la recta (% de mortalidad vs. concentración del insecticida) significa que la mayoría de los individuos manifiestan similar respuesta ante la aplicación de las diferentes dosis del insecticida.
Se determinó la susceptibilidad y/o resistencia a insecticidas organofosforados y al carbamato propoxur en las cepas de TRUJILLO y TUMBES, del Perú. Como se muestra en la tabla 2, la cepa TRUJILLO mostró resistencia a los organofosforados clorpirifos y temefos y al carbamato propoxur con valores de factor de resistencia (FR50) de 25,45x, 22,5x y 20,25x, respectivamente. Esta cepa mostró valores moderados (FR50 entre 5 x y 10 x) de resistencia a fention (7,27x) y a fenitrotion (7,64x) y completa susceptibilidad (FR50 ≤ 5x) a malation (2,11x). Sin embargo, la cepa TUMBES mostró susceptibilidad en larvas a todos los organofosforados evaluados, excepto a fention, al cual mostró moderada resistencia (6,59 x).
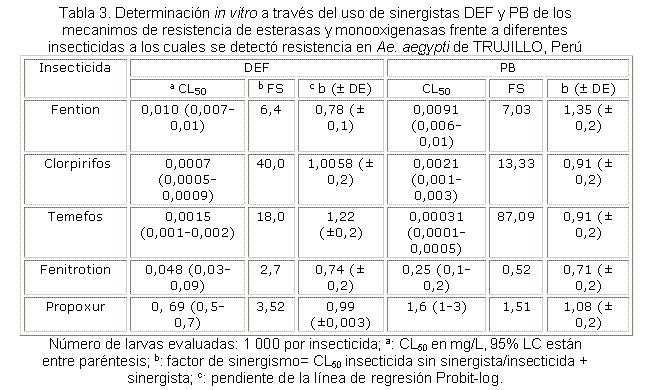
Se determinó in vivo a través del uso de sinergistas (tabla 3) el papel de algunas enzimas en la resistencia a insecticidas. Los sinergistas potencian la acción de los insecticidas inhibiendo enzimas como las esterasas y GST (sinergista DEF), monooxigenasas (sinergista PB). Al comparar la concentración letal media (CL50), obtenida para el clorpirifos antes y después de aplicar el sinergista DEF en la cepa TRUJILLO, el valor de la CL50 disminuyó 40 veces, lo cual se expresa como factor de sinergismo (FS). En el caso del temefos al aplicar DEF el valor del FS fue de 18x, lo cual implica a las esterasas y GST en la resistencia a temefos y clorpirifos detectada en la cepa TRUJILLO. También se observó un valor alto, mayor que 5x, para fention, lo cual implica que estas enzimas pueden ser responsables de la resistencia moderada detectada a fention.
De acuerdo con el valor de FS al aplicar el sinergista PB, las enzimas monooxigenasas desempeñan también un papel importante en la alta resistencia detectada a clorpirifos y temefos, con valores de FS de 13,3 y 87,09, respectivamente; y también fueron estas enzimas responsables de la resistencia moderada a fention (FS= 7,03) en la cepa TRUJILLO. De acuerdo con los resultados obtenidos con los sinergistas, ni las esterasas, ni la GST, ni las monooxigenasas intervienen en la resistencia detectada a fenitrotion y propoxur, dado por los valores de FS menores que 5, con valores de 0,52 y 1,51, respectivamente.
En la figura 1 se muestran los valores de frecuencia en que se encuentra incrementada la actividad de las enzimas esterasas, GST y la frecuencia en que se encontró modificada la enzima acetilcolinesterasa (AchE). Como se puede observar se corresponden muy bien los altos valores de frecuencia de cada una de estas enzimas en la cepa TRUJILLO, con valores de 47,02 %, 38,2 % y 18,7 % para esterasas, GST y AchE, respectivamente. Sin embargo y en correspondencia con la alta susceptibilidad a insecticidas observada en la cepa TUMBES, la frecuencia de los mecanismos de resistencia de esterasas, GST y AchE fue baja con valores de 3,2 %, 5,2 % y 0,4 %, respectivamente.
Se evaluó también el nivel de resistencia en adultos, en las cepas de Aedes aegypti, al insecticida organoclorado DDT, a los piretroides betacipermetrina, deltametrina y lambdacialotrina y al organofosforado clorpirifos. Como se observa en la tabla 4 la cepa TRUJILLO solo es susceptible (% de mortalidad mayor que 98 %) a los insecticidas betacipermetrina (98,24 %) y deltametrina (98,16 %) y al organofosforado clorpirifos (98,06 %), sin embargo, es resistente (% de mortalidad menor que 80 %) a DDT (38,5 %) y lambdacialotrina (56,34 %). La cepa TUMBES mostró resistencia a DDT (22,4 %) y a todos los piretroides, betacipermetrina (31,03 %), deltametrina (45,9 %), y lambdacialotrina (33,9 %). Esta cepa solo fue susceptible a clorpirifos (99,01 %).
Los resultados de la electroforesis en gel de poliacrilamida se muestran en la figura 2. Se observó la presencia de la banda de esterasa A4 incrementada en la cepa TRUJILLO, pero no se observó ni en la cepa TUMBES, ni en la cepa susceptible.
DISCUSIÓN
Con el presente estudio llevado a cabo utilizando Aedes aegypti procedente de dos provincias del Perú (TRUJILLO y TUMBES), se determinó el nivel de susceptibilidad y/o resistencia en larvas a insecticidas organofosforados (malation, fention, clorpirifos, temefos y fenitrotion), y al carbamato propoxur (BAYGON). En adultos se determinó la resistencia a piretroides (betacipermetrina, deltametrina, lambdacialotrina) y al organofosforado clorpirifos. Las larvas de la cepa TRUJILLO resultaron susceptibles a malation y pirimifos metil; sin embargo, la resistencia en esta cepa a temefos y clorpirifos fue alta y moderada a fention y fenitrotion. A pesar de que malation ha sido ampliamente utilizado tanto por los programas de control de salud pública, como por la agricultura, existen recientes reportes de que esta especie es aún susceptible a este insecticida.5,6,15-17 Sin embargo sí existen reportes de resistencia a temefos.5,6,14-20 Durante la epidemia de dengue ocurrida en Ciudad de La Habana 2001-2002 se evaluaron los municipios Playa5 y Guanabacoa,6 los cuales fueron sometidos a aplicación de insecticidas (cipermetrina y clorpirifos en ciclos alternos) y las larvas de estos municipios resultaron ser susceptibles a los organofosforados malation, clorpirifos y pirimifos metil, sin embargo mostraron alta resistencia a temefos y fention. En un estudio de susceptibilidad en México13 se demostró, en larvas, completa susceptibilidad a malation y clorpirifos. En un estudio realizado en 2 localidades de Panamá se encontró que las larvas mostraron resistencia a pirimifos metil, y fueron susceptibles a temefos, malation, fention, fenitrotion y clorpirifos.21
A través de estudios con sinergistas se demostró que las enzimas esterasas y monooxigenasas intervienen en la alta resistencia a temefos y clorpirifos y moderada a fention en la cepa TRUJILLO; sin embargo, no son responsables de la moderada resistencia detectada a fenitrotion. Está bien documentado en la literatura el papel de las enzimas de acción metabólica basadas en la alta actividad de esterasas, glutation transferasa y monooxigenasas en la resistencia a insecticidas organofosforados como temefos y clorpirifos.6,8,22 Algunos autores asocian el mecanismo de alta actividad de esterasa como principal mecanismo de resistencia a temefos en Aedes aegypti.5,6,20,23
En este trabajo se observó que la frecuencia de los mecanismos de resistencia de esterasas GST, se encontró elevada en la cepa TRUJILLO y no en TUMBES, lo cual se corresponde bien con los resultados obtenidos en esta última cepa, que fue completamente susceptible a todos los insecticidas evaluados en larvas, excepto la moderada resistencia que mostró a fention.
Los mosquitos adultos de la cepa TRUJILLO resultaron resistentes a DDT y a los piretroides lambdacialotrina y ciflutrina, pero no a betacipermetrina y deltametrina, ni al organofosforado clorpirifos. La cepa TUMBES, mostró diferente comportamiento en adultos, porque resultó ser resistente a DDT y piretroides, pero no al organofosforado clorpirifos. Es de destacar que la provincia TUMBES ha estado sometida a una fuerte aplicación de insecticidas, sobre todo adulticidas para el control de la malaria, entre ellos DDT, el cual fue aplicado desde 1957 a 1960, 1980 a 1992, y a partir de 1993 fue sustituido por piretroides.
La resistencia a los piretroides en estado adulto es atribuible a que la mayoría de estos insecticidas han sido utilizado, bajo condiciones de epidemia o incremento de las poblaciones de Aedes aegypti, y siempre como adulticidas. Se ha asociado la resistencia a piretroides por el efecto de resistencia cruzada entre DDT y piretroides a través del mecanismo de knockdown tipo Kdr, pero también esta resistencia se ha asociado a mecanismos de resistencia de acción metabólica.24,25 En Cuba, durante la epidemia ocurrida en Ciudad de La Habana en los años 2001-2002 se utilizaron los insecticidas: clorpirifos (Terfos) y cipermetrina (Galgotrin) y las pruebas de efectividad dieron 100 % de mortalidad para ambos insecticidas, utilizándolos en tratamientos térmicos.26,27 Se logró bajar las infestaciones del vector y controlar la epidemia satisfactoriamente.
La electroforesis en gel de poliacrilamida reveló la presencia de la esterasa A4 reportada en Aedes aegypti de Santiago de Cuba (Rodríguez y otros), la cual no se observó amplificada, ni en TUMBES, ni en la cepa susceptible de referencia ROCKEFELLER.
Sobre la base de los resultados obtenidos, los Aedes aegypti, procedentes de 2 provincias del Perú muestran alta resistencia a DDT y piretroides, de ahí que el gen Kdr, debe estar presente en estas poblaciones, por lo que se recomienda establecer estrategias de control, utilizando los piretroides en esquemas de rotación o alternando con insecticidas que posean diferentes mecanismos de resistencia; como fue en este caso el organofosforado clorpirifos, al cual ambas cepas mostraron susceptibilidad. Con respecto al temefos, larvicida que aún se utiliza para el control focal, se recomienda monitorear la resistencia a este, probar su efectividad en el terreno y en caso de resistencia, utilizar las alternativas para el control focal de Aedes aegypti que recomienda la Organización Mundial de la Salud.
REFERENCIAS BIBLIOGRÁFICAS
1. World Health Organization. Pesticides and their application. 6th edition. Geneva:WHO/CDS/WHOPES/GCDPP/; 2006.
2. Georghiou GP, Wirth M, Tran H, Saume F, Knudsen AB. Potential for organophosphate resistance in Aedes aegypti in the Caribbean area and neighbouring countries. J Med Entomol. 1987;24:290-4.
3. Rawlins SC, Ragoonansingh R. Comparative organophosphorous insecticide susceptibility in Caribbean populations of Aedes aegypti and Toxorynchites moctezuma. J Am Mosq Control Assoc. 1990;6:315-7.
4. Ping LT, Yatiman R, Gek LP. Susceptibility of adult field strains of Aedes aegypti and Aedes albopictus in Singapore to pirimiphos-methyl and permethrin. J Am Mosq Control Assoc. 2001;17:144-6.
5. Bisset JA, Rodríguez MM, Fernández D, Pérez O. Estado de la resistencia a insecticidas y mecanismos de resistencia en larvas del municipio Playa, colectadas durante la etapa intensiva contra el Ae. aegypti en Ciudad de La Habana, 2001-2002. Rev Cubana Med Trop. 2004;56:61-6.
6. Rodríguez MM, Bisset JA, Fernández D, Pérez O. Resistencia a insecticidas en larvas y adultos de Aedes aegypti: prevalencia de la esterasa A4 asociada con la resistencia a temefos. Rev Cubana Med Trop. 2004;56:54-60.
7. Hemingway J, Boddington RG, Harris J. Mechanisms of insecticide resistance in Aedes aegypti (L.) (Diptera: Culicidae) from Puerto Rico. Bull Ent Res. 1989;79:123-30.
8. Vaughan A, Chadee DD, Ffrench-Constan, R. Biochemical monitoring of organophosphorous and carbamate insecticide resistance in Aedes aegypti mosquitoes from Trinidad. Med Vet Entomol. 1998;12:318-21.
9. Wirth MC, Georghiou GP. Selection and characterization of temephos resistance in a population of Aedes aegypti from Tortola, British Virgin Islands. J Am Mosq Control Assoc. 1999;15:315-20.
10. Rodríguez MM, Bisset JA, Mila L, Calvo E, Díaz C, Soca LA. Niveles de Resistencia a insecticidas y sus mecanismos en una cepa de Aedes aegypti de Santiago de Cuba. Rev Cubana Med Trop. 1999;51:93-8.
11. World Health Organization. Instructions for determining the susceptibility or resistance of mosquito larvae to insecticides. Geneva:WHO/VBC/81.80; 1981. p. 6.
12. Raymond M. Presentation d'une programme d'analyse log-probit pour microordinateur cahiers. Orstrom Sér Ent Méd Parasitol. 1985;23:117-21.
13. Booth JE, Boyland E, Sims P. An enzyme from the rat liver catalyzing conjugation with glutathione. Biochem J. 1961;79:516-23.
14. Rodríguez MM, Bisset JA, Molina DF, Lauzan L, Soca A. Detection of resistance mechanisms in Aedes aegypti from Cuba and Venezuela. J Med Entomol. 2001;38:623-8.
15. Sames WJ 4th, Bueno R Jr, Hayes J, Olson JK. Insecticide susceptibility of Aedes aegypti and Aedes albopictis in the Lower Río Grande Valley of Texas and Mexico. J Am Mosq Control Assoc. 1996;12:487-90.
16. Rodríguez MM, Bisset J, Díaz C, Soca A. Resistencia cruzada a piretroides en Aedes aegypti de Cuba inducido por la selección con el insecticida organofosforado malation. Rev Cubana Med Trop. 2003;55:105-11.
17. , Sharma SN, Saxena VK, LaI S. Study on susceptibility status in aquatic and adult stages of Aedes aegypti and Ae. albopictus against insecticides at international airports of South India. J Commun Dis. 2004;36:177-81.
18. Ponlawat A, Scott JG, Harrington LC. Insecticide susceptibility of Aedes aegypti and Aedes albopictus across Thailand. J Med Entomol. 2005;42:821-5.
19. Paeporn P, Komalamisra N, Deesin V, Rongsrivam Y, Eshita Y, Thongrungkiat S. Temephos resistance in two forms of Aedes aegypti and its significance for the resistance mechanisms. Southeast Asian J Trop Med Public Health. 2003;34:786-92.
20. Braga IA, Lima JB, Soares Sda, Valle Dom Aedes aegypti resistance to temephos during 2001 in several municipalities in the state of Río de Janeiro, Sergipe, and Alagoas, Brazil. Mem Inst Oswaldo Cruz. 2004;99:199-203.
21. Saelim V, Brogdon WG, Rojanapremsuk J, Suvannadaba S, Pandii W, Jones JW, Sithiprasasna R. Bottle and biochemical assays on temephos resistance in Aedes aegypti in Thailand. Southeast Asian J Trop Med Public Health. 2005;36:417-25.
22. Bisset JA, Rodríguez MM, Lorenzo Cáceres. Niveles de resistencia a insecticidas y sus mecanismos en dos cepas de Aedes aegypti de Panamá. Rev Cubana Med Trop. 2003;55:191-5.
23. Mazarri MB, Georghiou GP. Characterization of resistance to organophosphate, carbamate, and pyrethroid insecticides in field populations of Aedes aegypti from Venezuela. J Am Mosq Control Assoc. 1995;11:315-22.
24. Rodríguez MM, Bisset JA, Ruiz M, Soca A. Cross-resistance to pyrethroid and organophosphate insecticides induced by selection with temephos in Aedes aegypti (Diptera: Culicidae) from Cuba. J Med Entomol. 2002;39:882-8.
25. Vontas JG, Small G, Hemingway J. Glutathione s transferases as antioxidant defence agents confer pyrethroid resistance in Nilaparvata lugens. Biochem J. 2001;357:65-72.
26. Rodríguez MM, Bisset JA, De Armas Y, Ramos F. Pyrethroid insecticide-resistant strain of Aedes aegypti from Cuba induced by deltamethrin selection. J Am Mosq Control Assoc. 2005;21:437-45.
27. Montada D, Zaldívar de Zayas J, Figueredo D, Suárez S, Leyva M. Eficacia de los tratamientos intradomiciliarios con los insecticidas cipermetrina, lambdacialotrina y clorpirifos en una cepa de Aedes aegypti. Rev Cubana Med Trop. 2006;58(2):148-53. Disponible en: http://bvs.sld.cu/revistas/
Recibido: 27 de noviembre de 2006.
Aprobado: 23 de enero de 2007.
Dr. Juan A. Bisset. Instituto de Medicina Tropical "Pedro Kourí". AP 601. Marianao 13. Ciudad de La Habana. Correo electrónico: bisset@ipk.sld.cu
Instituto Nacional de Salud. Perú.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}