










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Cubana de Medicina Tropical
On-line version ISSN 1561-3054
Rev Cubana Med Trop vol.60 no.1 Ciudad de la Habana Jan.-Apr. 2008
ARTÍCULOS ORIGINALES
Obtención de clones recombinantes que expresan las proteínas prM-E-NS1 del virus dengue serotipo 2
Obtaining recombinant clones expressing dengue virus serotype 2 prM-E-NS1 proteins
Dra. Mayra Muné JiménezI; Dr. Donald HarnII; Lic. Yudira SotoIII; Lic. Rosa RamírezIV; Téc. Melkis AlfonsoV; Dr. Akran D´daraVI
I Doctora en Ciencias de la Salud. Licenciada en Bioquímica. Investigadora Titular. Instructora. Departamento de Virología. Instituto de Medicina Tropical "Pedro Kourí" (IPK).
II Doctor en Ciencias de la Salud. Doctor en Medicina. Investigador Titular. Escuela de Salud Pública de Harvard. Universidad de Harvard.
III Máster en Virología. Licenciada en Microbiología. Investigadora Agregada. Departamento de Virología. IPK.
IV Máster en Biotecnología. Licenciada en Bioquímica. Investigadora Agregada. Asistente. Departamento de Virología. IPK.
V Técnica en Farmacia Industrial. Departamento de Virología. IPK.
VI Doctor en Ciencias de la Salud. Doctor en Medicina. Investigador Titular. Escuela de Salud Pública de Harvard. Universidad de Harvard.
RESUMEN
OBJETIVO: obtener clones recombinantes que expresen diferentes proteínas del virus dengue 2 en un vector de expresión en células eucariotas.
MÉTODOS: se realizó el clonaje de los genes prM, envoltura (E) y 65 aminoácidos (aa) de la proteína NS1 del virus dengue serotipo 2 (cepa Nueva Guinea C) en un vector de expresión en células eucariotas pcDNA (3.1). La obtención de los genes correspondientes a la zona prM/E/NS1 y prM/E truncada en 100 aa se realizó mediante reacción en cadena de la polimerasa (RCP). La detección de los posibles clones recombinantes se llevó a cabo mediante las técnicas de RCP, análisis de restricción enzimática y secuenciación nucleotídica. Se realizó la transfección de la línea celular CHO con cada plásmido recombinante. Para determinar la expresión transciente de los genes clonados se empleó la técnica de inmunofluorescencia indirecta (IFI) y trascripción reversa-RCP (TR-RCP).
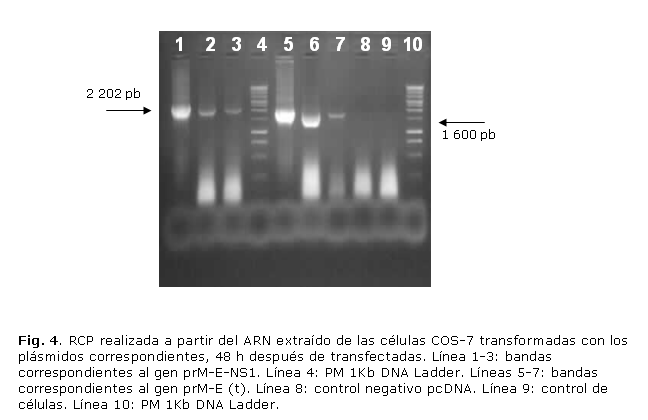
RESULTADOS: se obtuvieron bandas de 2 202 y 1 600 pares de bases (pb), respectivamente. Se estudiaron 20 posibles colonias recombinantes, de las cuales, 7 resultaron positivas para prM-E-NS1 y 5 para prM/E truncada. Se obtuvieron células fluorescentes 48 h después de transfectadas, además una RCP positiva a ese mismo tiempo, lo que indicó la presencia de las proteínas en las células transfectadas.
CONCLUSIONES: el vector empleado fue eficiente para el clonaje y la expresión de las proteínas seleccionadas, por lo que las construcciones genéticas obtenidas pudieran ser evaluadas en animales como posibles candidatos vacunales para la obtención de una vacuna de ADN contra el dengue.
Palabras clave: Clonaje, genes, dengue 2, transfección, vacuna de ADN.
SUMMARY
OBJECTIVE: To obtain recombinant clones expressing different dengue virus 2 proteins in an expression vector of eukaryote cells.
METHODS: Cloning of prM genes, E envelope and 65 amino-acids (aa) of dengue virus serotype 2 NS1 proteins (Nueva Guinea strain) in an expression vector of pcDNA eukaryote cells (3.1). The prM/E/NS1 zone and the truncated prM/E zone at 100 aa genes were obtained by polymerase chain reaction (PCR). Possible recombinant clones were detected using PCR, enzyme restriction analysis and nucleotide sequencing. Transfection of the CHO cell line with each recombinant plasmid was performed. Indirect immunofluorescence and reverse transcription-polymerase chain reaction (RT-PCR) determined the transient expression of cloned genes.
RESULTS: Bands of 2 202 and 1 600 base pairs (bp) respectively were obtained. Twenty possible recombinant colonies were studied, 7 of them were pr-E-NSI-positive and 5 truncated prM/E-positive. Fluorescent cells emerged 48 hours after being transfected in addition to positive PCR, all of which indicated that the studied proteins were present in transfected cells.
CONCLUSIONS: The used vector proved to be efficient for cloning and expression of the selected proteins; therefore, the obtained genetic constructions could be evaluated in animals as likely vaccinal candidates for a dengue virus DNA vaccine.
Key words: Cloning, genes, dengue 2, transfection, DNA vaccine.
INTRODUCCIÓN
Hoy día el dengue es considerada la enfermedad viral transmitida por artrópodos de mayor importancia en términos de morbilidad y mortalidad humana.1 Millones de personas son infectadas anualmente y la enfermedad se reporta en un gran número de países del Sudeste asiático, Pacífico occidental, Mediterráneo, Centroamérica y Sudamérica.2 Los virus del dengue pertenecen a la familia Flaviviridae género Flavivirus. El complejo dengue está formado por 4 serotipos: Dengue 1 (D1), Dengue 2 (D2); Dengue 3 (D3) y Dengue 4 (D4) los que se transmiten en la naturaleza en un ciclo mosquito-hombre-mosquito, y el principal vector es el mosquito Aedes aegypti. La infección por cualquier serotipo puede producir una infección asintomática, una enfermedad benigna, la fiebre del dengue (FD) o la forma severa de la enfermedad, la fiebre hemorrágica del dengue/síndrome de choque por dengue (FHD/SCD) responsable de un elevado número de fallecidos, principalmente en el Sudeste asiático, el Pacífico occidental y las Américas.3
El desarrollo de una vacuna contra el dengue se considera de máxima prioridad tanto para las poblaciones de los países endémicos como para los viajeros. Esta vacuna podría ser utilizada también para prevenir las epidemias que pudieran desarrollarse en el futuro en los países no endémicos. Una vacuna contra el dengue debe brindar protección de larga duración frente a la infección por cualquiera de los 4 serotipos del dengue, o debe prevenir la enfermedad. Adicionalmente, una vacuna contra el dengue deberá evitar el riesgo teórico del fenómeno de inmunoamplificación y debe ser segura y eficaz en niños y adultos. Por último, debe ser barata y accesible para los países pobres.4
Varias estrategias vacunales están siendo evaluadas tanto en estudios preclínicos como en voluntarios, una de estas estrategias consiste en la inmunización con ADN,5-7 la cual consiste en la inoculación a un organismo de un ADN plasmídico que lleva un gen que codifica para un determinado antígeno. Este ADN es captado por células del tejido inoculado, donde se expresa el antígeno e induce una respuesta inmune frente a este. Este tipo de inmunización ofrece algunas ventajas como son: el ADN contiene la secuencia codificante de interés y los elementos reguladores esenciales para que se exprese; ocasiona la generación de una fuerte respuesta humoral y celular, contra el antígeno codificado por el plásmido debido a su presentación por vía MHCI y MHCII; el proceso de obtención del ADN plasmídico es relativamente barato y fácil. Además el ADN plasmídico es estable frente al calor, lo que permitiría la liofilización de potenciales vacunas, esto hace que se convierta en un candidato muy atractivo para su uso en países subdesarrollados donde la economía y los factores climáticos, entre otros, son factores limitantes para la obtención de candidatos vacunales.8 Los autores de este trabajo se propusieron la obtención de clones recombinantes que expresen diferentes proteínas del virus Dengue 2 en un vector de expresión en células eucariotas, los que serán posteriormente evaluados como vacunas de ADN para determinar la respuesta inmune inducida en animales de experimentación.
MÉTODOS
Clonaje de los genes. PrM-E-NS1 en el vector pcDNA 3.1
Obtención del inserto
Para la obtención del inserto correspondiente a los genes prM-E-NS1 se realizó una reacción en cadena de la polimerasa (RCP) empleando como molde el plásmido NGC que contiene el genoma completo del virus dengue 2, gentilmente donado por la doctora Irene Bosch (Universidad de Massachussets). Para ello se utilizaron cebadores específicos para esta zona cuya secuencia es la siguiente:
5´GCTCGAGATGGTGATTATTATGTT 3´ 5´GTCTAGATTAATTTTCCAGTCTTGTTACTGAGCG 3´
Estos oligos dan un producto de amplificación de 2,238 pb con los sitios XhoI y XbaI en sus extremos. Las condiciones de la RCP fueron las siguientes:
95 °C 5 min. Desnaturalización 95 °C 1 min., Hibridación 55 °C 1 min., Extensión 42 °C 2 min. El fragmento obtenido fue empleado posteriormente para el clonaje en el vector pcDNA 3.1 (Invitrogen).
Preparación del vector pcDNA 3.1
El vector pcDNA 3.1 fue digerido con las enzimas XhoI y XbaI (Promega). Para ello se añadieron 5 mL (5 mg) del vector a una mezcla que contenía 1 mL de cada enzima, 5mL del tampón 10X, así como 38 mL de agua bidestilada estéril. La mezcla fue incubada durante 2 h a 37 °C. Transcurrido este tiempo se tomaron 10 mL de la digestión y fueron mezclados con 2 mL de bromofenol azul y glicerol 0,1 %. El ADN digerido fue analizado mediante una corrida electroforética en gel de agarosa 0,8 % en Tris Borato EDTA (TBE 1X). Para la inactivación de las enzimas la mezcla fue incubada a 65 °C durante 15min. Posteriormente se procedió a la purificación del vector, aplicándose los 25 mL restantes en un gel de agarosa de bajo punto de fusión (LGT). Posteriormente el ADN fue extraído por el método del fenol.9
Ligazón
Para la reacción de ligazón se empleó 1 mL del vector digerido (10 ng) y 1 mL de banda (100 ng), 1 mL de la enzima T4 ligaza (Promega), 2 mL del tampón de la enzima 10X y 15 mL de agua para un volumen final de 20 mL. Además fueron incluidas 2 ligazones, una de control de banda y otra de control de vector.
Transformación
Se tomaron 100 mL de células TOP 10 (Promega) previamente descongeladas y se le adicionaron 10 mL de cada ligazón, se incubaron en hielo durante 20 min. Después se realizó un choque térmico a 42 °C durante 1 min, se colocaron de inmediato en hielo durante 2 min. Pasado ese tiempo se le añadió 1 mL de medio Luria-Bertani (LB) constituido por triptona 1 %, extracto de levadura 0,5 % y NaCl 1 %, se dejaron en incubación a 37 °C durante 1 h. Una vez concluido este paso se procedió a la siembra de 100 mL de células transformadas, en placas Petri que contenían medio LB sólido con ampicilina (LBA) a una concentración de 25 mg/mL. A continuación las placas fueron incubadas durante toda la noche a 37 °C hasta la aparición de las colonias.
Para realizar la selección de los transformantes se escogieron 10 colonias al azar, que fueron sembradas simultáneamente en placas con medio LBA sólido y tubos con 5 mL de medio líquido LBA, los cuales se incubaron toda la noche a 37 °C a una velocidad de agitación de 250 rpm. Después se procedió a la extracción del ADN plasmídico siguiendo la metodología descrita por el estuche comercial Wizar Plus Minipreps DNA Purification System, Promega.
Selección de los recombinantes
EL ADN extraído se chequeó mediante una corrida electroforética en gel de agarosa 0,8 % y se visualizó en un transiluminador de luz UV. La selección de los recombinantes se realizó mediante un análisis de restricción, digiriéndose los plásmidos con las enzimas XhoI (Promega) y XbaI (Promega). Para ello se emplearon 5 mL de ADN (1 mg/mL), 1 mL de cada enzima (10 U/mL), 3 mL de tampón D 10X (Promega) y 21 mL de agua, para un volumen final de 30 mL. La reacción se dejó en incubación durante 1 h a 37 °C. Transcurrido este tiempo se tomaron 10 mL de cada digestión y se mezclaron con 2 mL de bromofenol azul y glicerol 0,1 %. Estas mezclas se aplicaron a un gel de agarosa 0,8 % y se visualizó en un transiluminador.
Secuenciación del ácido nucleico
Se seleccionaron 2 clones positivos al azar para la secuenciación del ácido nucleico, se empleó el estuche comercial Thermo Sequenase Cy5 Dye Terminator Cycle Sequencing Kit, Promega, según instrucciones del fabricante. Para la secuenciación se utilizaron los mismos oligonucleótidos empleados en la RCP.
Clonaje de los genes. PrM-E-truncada en 100 aa del extremo C terminal en el vector pcDNA 3.1
Obtención del inserto
Para la obtención del inserto correspondiente al gen prM-E (truncado) se realizó una RCP para la amplificación de este fragmento que se corresponde con una talla de 1,6 Kb, el cual fue empleado posteriormente para el clonaje en el vector pcDNA 3.1. Como molde se empleó el plásmido NGC que contiene el genoma completo del virus dengue 2, cepa Nueva Guinea C. Para ello se utilizaron cebadores específicos para esta zona cuya secuencia es la siguiente:
5´GCTCGAGATGGTGATTATTATGTT3´
5´ GTCTAGAATGGTGATTATTATGTTGATTCCAACAGCG 3´
Estos oligos dan un producto de amplificación de 1,6 (Kb) con los sitios XhoI y XbaI en sus extremos, las condiciones de la RCP son las siguientes:
95 °C 5 min, desnaturalización 95 °C 1 min, Hibridación 55 °C 1 min, Extensión 42 °C 2 min.
La preparación del vector, ligazón, transformación y selección de los clones recombinantes se realizó como ha sido descrito anteriormente para la obtención del gen prM-E-NS1.
Expresión transciente de las proteínas en las células CHO
Transfección
Se emplearon 2 mg/mL de cada plásmido para transfectar las células CHO. Así mismo se transfectaron 2 mg/mL del vector pcDNA 3.1, el cual fue utilizado como control negativo. Se prepararon 2 mezclas para cada plásmido. La mezcla A compuesta por 500 mL de medio RPMI sin suero de ternera fetal y 40 mL del plásmido, y la mezcla B compuesta por 500 mL de medio y 4 mL de Transfectan (Promega) o reactivo de transfección a una concentración de 1 mL/mg.
Para la transfección se utilizaron 2 placas de 6 pozos, sembradas con células CHO a 2,5 x 105 células por pozo y con 70 % de confluencia. Se eliminó el medio de las células y fueron lavadas 2 veces con medio sin suero de ternera y posteriormente se le añadieron 500 mL de medio sin suero, a cada pocillo. A continuación se mezclaron las soluciones A y B y se añadieron 500 mL de esta mezcla a cada pozo. Se incubaron las placas durante 2 h a 37 °C en atmósfera de CO2. Trascurrido este tiempo se añadieron 3 mL de medio completo (RPMI + 10 % de suero de ternera) y se dejó en incubación a 37 °C en atmósfera de CO2. Posteriormente se chequeó la expresión por inmunofluorescenica indirecta (IFI) a las 48 y 72 h postransfección. A las 24 h se eliminó el medio de las células, y fueron desprendidas mediante raspado con un policía de goma. Se colocaron en un tubo eppendorff y se centrifugaron durante 5 min a 10 000 rpm. El precipitado fue lavado 2 veces con solución fosfato tamponada (PBS) 1X y finalmente se resuspendieron en 50 mL de PBS 1X. La lámina se dejó secar durante 20 min y después se introdujo en acetona, para fijar las células. A continuación se añadió una dilución 1/40 de líquido ascítico hiperinmune anti-D2 y se incubó durante 1 h a 37 °C. Después se realizaron 3 lavados de 5 min con PBS 1X y se añadió el conjugado (IgG de carnero anti IgG de ratón conjugado con fluoresceína), se incubaron durante 30 min a 37 °C. Se repitieron los lavados con PBS1X, las láminas fueron observadas en un microscopio de fluorescencia (Leitz).
RCP a partir de las células transfectadas
Después de incubar las células durante 48 y 72 h, las monocapas fueron desprendidas con un policía de goma y recuperadas por centrifugación a 10 000 rpm durante 10 min. Seguidamente se realizó la extracción de ARN mediante el protocolo de extracción por TRIZOL (Sigma). Se utilizaron 300mL de cultivo celular transfectado, y se le adicionaron 500 mL de Trizol, se incubaron durante 5 min a temperatura ambiente. A continuación se realizó la extracción por cloroformo. La fase acuosa fue transferida a un nuevo tubo y se le adicionó 400 µL de isopropanol, se mezcló mediante agitación manual y se incubó durante 10 min a temperatura ambiente. EL ARN fue precipitado al centrifugarse a 10 000 rpm durante 30 min a 4 °C. El precipitado obtenido fue lavado mediante la adición de 500 µL de etanol 75 %, se centrifugó otra vez, de la misma forma. Una vez seco se resuspendió en 20 µL de agua estéril tratada con dietilpirocarbonato 0,01 % (Sigma). A este precipitado se le realizó un tratamiento con la enzima ADNasa (RQ1 RNase-free DNase, Promega) 1 h a 37 °C. La metodología para la RCP se realizó siguiendo el protocolo antes descrito para la obtención de los insertos.
RESULTADOS
Se realizaron 2 construcciones genéticas en el vector de expresión en células eucariotas pcDNA 3.1 que contenían los genes prM, envoltura y 65 aminoácidos de la proteína NS1 del virus Dengue, serotipo 2. Para la obtención de los genes correspondientes se realizó una RCP con los oligos específicos, se obtuvo en cada caso una banda a la talla adecuada (2,2Kb y 1,6Kb), las cuales fueron clonadas en el vector previamente digerido con las enzimas XhoI y XbaI (fig. 1).
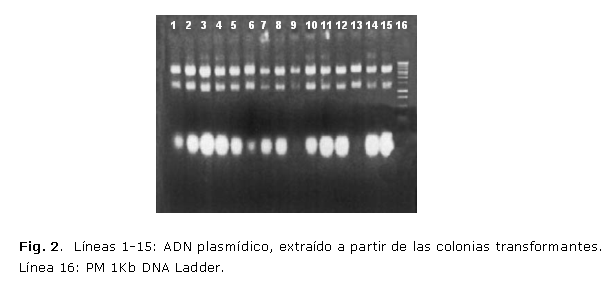
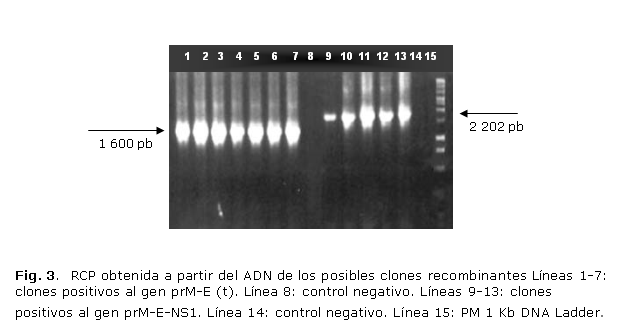
El ADN obtenido a partir de los posibles plásmidos recombinantes tuvo la calidad y concentración adecuadas para ser utilizados en las técnicas de RCP y el análisis de restricción enzimática (fig. 2). De los 10 clones analizados 5 resultaron recombinantes para prM-E-NS1 y 7 para prM/E truncada lo cual fue determinado por las técnicas de RCP y análisis de restricción enzimática. En ambas técnicas se obtuvieron bandas específicas a la talla adecuada, que fue comprobado en la corrida electroforética de los productos de la RCP y los productos digeridos (fig. 3). La inserción correcta de los genes clonados fue igualmente corroborada mediante secuenciación nucleotídica del ADN de los plásmidos correspondientes. Para ser secuenciados se seleccionaron al azar 2 de los clones positivos para su secuenciación. En ambos se pudo comprobar la inserción correcta del fragmento clonado cuya secuencia coincidía con la reportada en la literatura para los genes seleccionados.
Concluidos los experimentos de clonaje se determinó la expresión transciente de los genes en las células eucariotas (CHO) mediante la transfección de estas células con los plásmidos correspondientes previamente purificados por la técnica de "Minipreps". Se determinó la expresión de los genes mediante IFI de las células transfectadas, 48 y 72 h postransfección, se obtuvo una inmunofluorescencia positiva 48 y 72 h después de ser transfectadas. Aproximadamente 80 % de las células fueron transfectadas, se observó una fuerte señal fluorescente, los experimentos realizados con el plásmido control (plásmido sin inserto) fueron negativos.
Igualmente se realizó la RCP a partir del ARN extraído de las células transfectadas, se obtuvieron bandas específicas a las 48 y 72 h después de la transfección con cada plásmido (fig. 4).
DISCUSIÓN
En 1990 surge una nueva estrategia de vacunación, la cual consiste en la inoculación de un ADN plasmídico in vivo, que codifica para determinados antígenos de interés, con la inoculación intradérmica o intramuscular del ADN desnudo. El proceso de transcripción se produce dentro de las células del hospedero así como la presentación al sistema inmune de la proteína codificada. La inmunización con ADN desnudo supera muchas de las desventajas que se presentan en las estrategias para el desarrollo de vacunas convencionales.10
Las vacunas de ADN contra Flavivirus han evolucionado en los últimos años, proporcionando un acercamiento prometedor para la comprensión de algunos factores clave involucrados en la inducción de una respuesta inmune eficiente. Sin embargo, a pesar de los esfuerzos para desarrollar vacunas de ADN, vacunas de subunidad, vacunas vivas atenuadas, o vacunas quiméricas, no existe en la actualidad una vacuna disponible contra el dengue segura y efectiva.11 El mayor obstáculo para desarrollar una vacuna contra el dengue es la observación epidemiológica de que la infección por el virus dengue solo produce inmunidad protectora contra el serotipo infectante y que la infección secundaria por un serotipo heterólogo incrementa el riesgo de fiebre hemorrágica del dengue, síndrome de choque por dengue (FHD/SCD).12
Las vacunas de ADN ofrecen una estrategia alternativa esperanzadora para el desarrollo de una vacuna contra el dengue. Esta metodología en la cual el plásmido de ADN que expresa el antígeno de interés es utilizado como vacuna ha mostrado inducir una respuesta inmune contra diferentes virus en modelos animales incluidos varios Flavivirus.13-16 Una vacuna de ADN contra el virus del Oeste del Nilo ha sido recién aprobada para el uso en caballos, resulta la primera vacuna de ADN comercializada en el mercado. Previamente, por otros autores ha sido reportada la factibilidad de una vacuna de ADN contra el virus del dengue.17-23
Los resultados alentadores publicados por varios autores incitaron al desarrollo, en nuestro laboratorio, de varias construcciones genéticas que contenían proteínas del virus dengue 2, las cuales una vez inoculadas en ratones y primates no humanos permitirán estudiar la respuesta inmune inducida en estos animales de experimentación.
En este trabajo se realizó el clonaje de las proteínas PrM_E y 65 aa de la proteína no estructural NS1 en el vector de expresión en células eucariotas pcDNA 3.1. La glicoproteína E (55-60 kDa) constituye el componente proteico principal en la superficie de la partícula viral. Se ha propuesto que en el dominio III está presente el receptor viral que se une a los receptores celulares.24,25 En la proteína E radican las principales propiedades biológicas del virus como enlace al receptor, la fusión, la inducción de anticuerpos neutralizantes de importancia en la respuesta inmune protectora, y el desarrollo de amplificación dependiente de anticuerpos (ADA). Esta proteína es también diana para los linfocitos T auxiliadores.26 Además ha sido evaluado el papel protector de las 2 formas de la proteína de membrana, PrM presente en los viriones inmaduros y M presente en el virión maduro.27 La proteína PrM es el precursor de la proteína M. El co-transporte de la PrM y del heterodímero de la envoltura a través de la vía exocítica es esencial en el proceso de maduración viral. La síntesis de E requiere de la cosíntesis de la PrM. La proteína M también desempeña un papel esencial en el control de la actividad de fusión de la proteína E. La proteína prM es necesaria para que la proteína E logre su conformación adecuada por lo que se ha sugerido que tiene un papel de proteína "chaperona"de la proteína E.28,29 Una síntesis eficiente y correcta de cualquier proteína de subunidad depende de su expresión en su conformación nativa, la cual es indispensable para el desarrollo de una vacuna de ADN efectiva.12
Aunque varias de las proteínas estructurales y no estructurales estimulan la inmunidad mediada por células T citotóxicas, las proteínas NS1 y NS3 constituyen la fuente principal de epítopes de células T CD4 + y CD8+.29,30
En el presente trabajo se realizó el clonaje eficiente de las proteínas PrM-E y NS1 del virus dengue 2 en el vector seleccionado como se evidenció por las técnicas de análisis de restricción y RCP. Además de los métodos antes utilizados, se realizó la secuenciación del ADN de los clones positivos. Este aspecto antes descrito es de vital importancia para los posteriores procesos de expresión de la proteína recombinante, de forma tal que la proteína sea traducida y expresada de forma correcta.
Las técnicas empleadas para determinar la expresión correcta de los plásmidos en las células transfectadas fueron la RCP y la inmunofluorescencia indirecta. Una vez concluida la extracción del ARN de las células, se realizó el tratamiento con la enzima ADNasa I para evitar posibles resultados falsos positivos por amplificación del ADN plasmídico. Como se pudo evidenciar se obtuvo una banda a la talla específica en las células transfectadas con ambos plásmidos, lo cual pudiera sugerir una correcta transcripción del ADN plasmídico en la células hospederas. Sin embargo, estos resultados no son del todo concluyentes para demostrar la correcta expresión de los plásmidos in vitro. Con este objetivo se empleó la técnica de inmunofluorescencia indirecta para la detección de las proteínas recombinantes expresadas en las células transfectadas. Los 2 plásmidos seleccionados fueron correctamente expresados en las células transfectadas como se evidenció por el reconocimiento de los antígenos específicos. Estos resultados coinciden con los reportados por otros autores que han utilizado plásmidos que contienen las proteínas PrM-E del virus dengue 2.31
Koechel y otros, en 1997, desarrollaron una vacuna de ácido nucleico contra el virus dengue 2.32 Los plásmidos construidos, que contenían los genes de PrM y 92 % de E, fueron capaces de inducir una respuesta de anticuerpos neutralizantes en 100 % de los ratones inmunizados; sin embargo, no se obtuvo protección significativa con posterioridad al reto con el virus salvaje. En esta misma vacuna, se evaluó también el uso de secuencias inmunoestimuladoras (CpG) en relación con el desarrollo de anticuerpos y de protección viral frente al reto. En este estudio, 60 % de los ratones inmunizados con la vacuna de ADN de dengue 2 sobrevivió al reto, comparado con 10 % de supervivencia en el grupo control. Esto sugiere
que la inmunización con vacunas de ADN puede constituir otra estrategia para el desarrollo de una vacuna contra el dengue.20,32-33
El presente trabajo demostró la factibilidad del clonaje de los genes prM /E y NS1 del virus dengue 2 en el vector de expresión en células eucariotas pcDNA 3.1. Para demostrar la inmunogenicidad de las vacunas de ADN en animales de experimentación serán necesarias investigaciones futuras centradas en el estudio y caracterización de la respuesta inmune humoral y celular inducida. Estos estudios serían de gran utilidad para corroborar la eficacia, seguridad y niveles de protección de estas formulaciones vacunales en modelos animales y su posible aplicación en humanos.
REFERENCIAS BIBLIOGRÁFICAS
1. Guzman MG, Kouri G: Dengue: an update. Lancet Infect Dis. 2002;2:33-42.
2. Halstead SB. Pathogenesis of dengue: challenges to molecular biology. Science. 1988;239(4839):476-81.
3. Guirakhoo F, Arroyo J, Pugachev KV, Miller C, Zhang, ZX, Weltzin R, et al. Construction, safety, and immunogenicity in nonhuman primates of a chimeric yellow fever-dengue virus tetravalent vaccine. J Virol. 2001;75(16):7290-304.
4. Mota J, Acosta M, Argotte R, Figueroa R, Mendez A, Ramos C. Induction of protective antibodies against dengue virus by tetravalent DNA immunization of mice with domain III of the envelope protein. Vaccine. 2005;23:3469-76.
5. Blair PJ, Kochel TJ, Raviprakash K, Guevara C, Salazar M, WuS , Olson JG, Porter KR. Evaluation of immunity and protective efficacy of a dengue-3 premembrane and envelope DNA vaccine in Aotus nancymae monkeys. Vaccine. 2006;24:1427-32.
6. Raviprakash K, Apt D, Brinkman A, Skinner C, Yang S, Dawes G, et al. A chimeric tetravalent dengue DNA vaccine elicits neutralizing antibody to all four virus serotypes in rhesus macaques. Virology. 2006;3:781-9.
7. Liu MA, Wahren B, Kasrlsson GB. DNA vaccines: recent development and future possibilities. Hum Gene. 2006;17(11):1051-61.
8. Maniatis T. Molecular cloning a laboratory manual 2nd ed. New York:Cold Spring Harbord Laboratory; 1989.
9. Jechlinger W. Optimization and delivery of plasmid DNA for vaccination. Expert Rev Vaccines. 2006;5(6):803-25.
10. Mota. J, Acosta M, Argotte R, Figueroa R, Mendez A, Ramos C. Induction of protective antibodies against dengue virus by tetravalent DNA immunization of mice with domain III of the envelope protein. Vaccine. 2005;23:3469-76.
11. Raviprakash K, Apt D, Brinkman A, Skinner C, Yang S, Dawes G, et al. A chimeric tetravalent dengue DNA vaccine elicits neutralizing antibody to all four virus serotypes in rhesus macaques. Virology. 2006;4:889-905.
12. Colombage G, Hall R, Pavy M, Lobigs M. DNA-based alphavirus-vectored immunisation with prM and E proteins elicits long-lived and protective immunity against the flavivirus, Murray Valley encephalitis virus. Virology. 1998;250(1):151-63.
13. Konishi E, Yamaoka M, Khin S, Kurane W, Mason I. Induction of protective immunity against Japanese encephalitis in mice immunization with a plasmid encoding Japanese encephalitis virus premembrane and envelope genes. J. Virol. 1998;72(6):4925-30.
14. Phillpotts R J, Venugopal K, Brooks T. Immunisation with DNA polynucleotides protects mice against lethal challenge with St. Louis encephalitis virus. Arch. Virol. 1996;141(3-4):743-9.
15. Schmaljohn C, Vanderzanden L, Bray M, Custer D, Meyer B, Rossi C, et al. Naked vaccines expressing the prM and E genes of Russian spring summer encephalitis virus and Central European encephalitis virus protect mice homologous and heterologous challenge. J Virol. 1997;71(12):9563-9.
16. Kochel T, Wu SJ, Raviprakash K, Hobart P, Hoffman S, Porter C. Inoculation of plasmids expressing the dengue-2 gene elicit neutralizing antibodies in mice. Vaccine. 1997;15(5):547-52.
17. Kochel TJ, Raviprakash K, Hayes CG, Watts DM, Russell KL, Phillips AS, et al. Dengue virus serotype-1 DNA vaccine induces virus neutralizing antibodies and provides protection from viral challenge in Aotus monkeys. Vaccine. 1998;3(27):3166-73.
18. Raviprakash K, Liu K, Matteucci M, Wagner R, Riffenburgh R, Carl B. Inhibition of dengue virus by novel, modified antisense oligonucleotides. J Virol. 1995;69(1):69-74.
19. Raviprakash K, Kochel TJ, Ewing D, Simmons M, Phillips I, Porter KR. Immunogenicity of dengue virus type 1 vaccines expressing truncated and full length envelope protein. Vaccine. 2000;22:2426-34.
20. Raviprakash K, Porter KR, Kochel TJ, Ewing D, Simmons PI, Murphy GS, et al. Dengue virus type 1 DNA vaccine induces protective immune responses in macaques. J Gen Virol. 2000;81(7):1659-67.
21. Raviprakash K, Marques E, Ewing D, Lu Y, Phillips I., Porter K, et al. Synergistic neutralizing antibody response to dengue virus type 2 DNA vaccine incorporation of lysosome-associated membrane protein sequences and of plasmid expressing GM-CSF. Virology. 2001;290(1):74-82.
22. Raviprakash K, Ewing D, Simmons M, Porter KR, Jones TR., Stout CG, et al. Needle-free Biojector injection dengue virus type 1 DNA vaccine with human immunostimulatory sequences and the GM-CSF gene increases immunogenicity protection from virus challenge in Aotus monkeys. Virology. 2003;315:345-52.
23. Zhang Y, Corver J, Chipman PR et al. Structures of immature flavivirus particles. Eur Mol Biol Organ J. 2003;22:2604-13.
24. Allison SL, Schalich J, Stiasny K, Mandl CW, Heinz FX. Mutational evidence internal fusion peptide in flavivirus envelope protein E. J Virol. 2001;75:4268-75.
25. Beasley DM, Aaswkov JG. Epitopes dengue 1 virus envelope protein recognized by neutralizing IgM monoclonal antibodies. Virology. 2001;279:447-58.
26. Roehrig JT. Immunochemistry of dengue viruses. In: Dengue and Dengue Hemorrhagic Fever. Gubler DJ, editor. London, UK: CAB International; 1997. p. 199-219.
27. Vazquez S, Guzman MG, Guillen G. Immune response to synthetic peptide dengue prM protein. Vaccine. 2002;20:1823-30.
28. Lorenz IC, Allison SL, Heinz FX, Helenius A. Folding and dimerisation of tick-borne encephalitis virus envelope proteins prM and E in the endoplasmic reticulum. J Virol. 2002;76:5480-91.
29. Mathew A, Kurane I, Green S. Predominance of HLA-restricted cytotoxic T-lymphocyte responses to serotypecrossreactive epitopes on non-structural proteins following natural secondary virus infection. J Virol. 1998;72:3999-4004.
30. Kurane I, Zeng L, Brinton MA, Ennis FA. Definition of an epitope on NS3 recognized by human CD4+ cytotoxic T-lymphocyte clones crossreactive for dengue virus types 2, 3 and 4. Virology. 1999;15:169-174.
31. Kochel T, Wu SJ, Raviprakash K, Hobart P, Hoffman S, Porter K, et al. Inoculation of plasmids expressing the dengue-2 envelope gene elicit neutralizing antibodies in mice. Vaccine. 1997;15(5):547-52.
32. Kurane I, Brinton MA, Samson AL, Ennis FA. Dengue virus-specific, human CD4+ CD8- cytotoxic T-cell clones: multiple patterns of virus cross-reactivity recognized by NS3-specific T-cell clones. J Virol. 1991;65(4):1823-8.
33. Kochel TJ, Raviprakash K, Hayes CG, Watts DM, Russell KL, Gozalo AS, et al. A dengue virus serotype-1 DNA vaccine induces virus neutralizing antibodies and provides protection from viral challenge in Aotus monkeys. Vaccine. 2000;18(27):3166-73.
Recibido: 29 de octubre de 2007.
Aprobado: 3 de diciembre de 2007.
Dra. Mayra Muné Jiménez. Instituto de Medicina Tropical Pedro Kourí. Subdirección de Microbilogía. Departamento de Virologia. Autopista Novia del Mediodía Km 6 ½ entre Carretera Central y Autopista Nacional. CP 17 100. Teléf.: 202 0450. Correo electrónico: mayra@ipk.sld.cu
Instituto de Medicina Tropical ¨Pedro Kourí¨


{kind=link}
{kind=link}
{kind=link}
{kind=link}