










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkPastos y Forrajes
On-line version ISSN 2078-8452
Pastos y Forrajes vol.38 no.1 Matanzas Jan.-Mar. 2015
ARTÍCULO DE INVESTIGACIÓN
Germinación y dormancia de arbustos y trepadoras del bosque siempreverde de la Sierra del Rosario, Cuba
Germination and dormancy of shrubs and climbing plants of the evergreen forest of Sierra del Rosario,Cuba
J. A. Sánchez1, Laura Montejo1, A. Gamboa1, Delhy Albert-Puentes1 y F. Hernández2
1Instituto de Ecología y Sistemática-Ministerio de Ciencia, Tecnología y Medio Ambiente, Carretera de Varona 11835, Calabazar, Boyeros, La Habana 19, CP 11900, Cuba
2 Reserva de la Biosfera Sierra del Rosario, Las Terrazas, Artemisa, Cuba
Correo electrónico: jasanchez@ecologia.cu
RESUMEN
Se determinaron los requerimientos germinativos y las clases de dormancia seminal en 14 especies de arbustos y 16 trepadoras (13 enredaderas y tres lianas), del bosque siempreverde estacional tropical de la Reserva de la Biosfera Sierra del Rosario, Cuba. También se evaluaron las relaciones entre las clases de dormancia y otros rasgos de historia de vida de las plantas. Los experimentos de germinación se realizaron en condiciones controladas de laboratorio (luz y temperatura): una temperatura constante de 25 ºC, tres condiciones de temperatura alterna (25/30 ºC, 25/35 ºC y 25/40 ºC) y dos ambientes lumínicos (luz y oscuridad). En los arbustos, la luz fue el principal factor que reguló la germinación; mientras que en las trepadoras los factores experimentados no afectaron significativamente la germinación en su totalidad, aunque algunas tuvieron una respuesta especie-específica. Solo dos especies fueron totalmente fotoblásticas positivas, y la germinación a la luz disminuyó con la masa fresca de la semilla. La dormancia ocurrió en 23 especies (76,6 %), de ellas 10 eran arbustos y 13, trepadoras. En los arbustos la dormancia fisiológica fue la más abundante, y en las trepadoras predominaron la fisiológica y la física. Entre las trepadoras se hallaron las únicas especies con dormancia morfológica y/o morfofisiológica. El mayor número de semillas dormantes se encontró en la estación seca (87 %). El estudio confirmó la diversidad de mecanismos de dormancia y requerimientos germinativos ya reportados en estas formas de vida para el trópico.
Palabras clave: Enredaderas, semilla, viabilidad.
ABSTRACT
The germination requirements and the seed dormancy classes were determined in 14 species of shrubs and climbing plants (13 vines and three lianas), of the tropical seasonal evergreen forest of the Sierra del Rosario Biosphere Reserve, Cuba. The relations between the dormancy classes and other traits of the life history of the plants were also evaluated. The germination trials were conducted under controlled laboratory conditions (light and temperature): a constant temperature of 25 ºC, three conditions of alternate temperature (25/30 ºC, 25/35 ºC and 25/40 ºC) and two light environments (light and darkness). In the shrubs, light was the main factor that regulated germination; while in the climbing plants the essayed factors did not significantly affect germination completely, although some had a species-specific response. Only two species were totally positive photoblastic, and the germination under light decreased with the fresh seed mass. Dormancy occurred in 23 species (76,6 %), from them 10 were shrubs and 13, climbing plants. In the shrubs the physiological dormancy was the most abundant, and in the climbing plants, the physiological and physical dormancy prevailed. Among the climbing plants the only species with morphological and/or morphophysiological dormancy were found. The highest number of dormant seeds was observed in the dry season (87 %). The study confirmed the diversity of dormancy mechanisms and germination requirements already reported in these life forms for the tropics.
Key words: Vines, seed, viability.
INTRODUCCIÓN
El ciclo natural de regeneración de las poblaciones de plantas es una serie concatenada de procesos demográficos (producción de semilla, dispersión, germinación, emergencia y establecimiento de la nueva plántula), que responde a la variabilidad ambiental actual (factores abióticos y bióticos) y a los cambios climáticos históricos (Marañón et al., 2008). Estos procesos biológicos, dependientes del microhábitat y de los mecanismos evolutivos que minimizan el riesgo de transición de las semillas a plántulas, están bajo la influencia de la presión de selección (Kos, Baskin y Baskin, 2012). Según Baskin y Baskin (2004), la presión de selección natural debe favorecer patrones de germinación, dormancia y longevidad que incrementen el éxito del establecimiento de la nueva plántula en un ambiente determinado. A pesar de esto, existe una gran mortalidad de las plántulas en condiciones naturales y en vivero (Norden et al., 2009); de ahí que el conocimiento de las clases de dormancia y los requerimientos germinativos permitiría incrementar el éxito de los planes de restauración tropical (Baskin y Baskin, 2005). Sin embargo, este conocimiento es muy escaso para las especies nativas de los bosques del Neotrópico, y gran parte de la información que existe se relaciona con las plantas de interés forestal (Sánchez et al., 2011). En este sentido, el primer reporte acerca de las clases de dormancia seminal en comunidades de árboles tropicales de bosques semideciduos de Panamá fue realizado por Sautu et al. (2007).
En Cuba, estos estudios también son muy restringidos; solo se cuenta con una contribución de Sánchez et al. (2012) sobre 60 especies arbóreas del bosque siempreverde estacional de la Reserva de la Biosfera Sierra del Rosario (RBSR), y no se han estudiado los mecanismos de regeneración de otras formas de vida tales como los arbustos, las lianas y las enredaderas. Por tanto, sería relevante estudiar sus requerimientos germinativos, las clases de dormancia y otros rasgos de vida a nivel de comunidad. Este conocimiento ayudaría en la conservación de las plantas nativas cubanas especialmente las que están en peligro de extinción, ya que, según Baskin y Baskin (1998), permitiría predecir los periodos más favorables para el establecimiento de las plántulas en la naturaleza, conocer la tendencia de las especies a conformar bancos de semilla de tipo transitorio o persistente, en función de la existencia o ausencia de mecanismos de dormancia/latencia y la obtención de plántulas en condiciones de vivero; así como realizar ensayos facultativos de germinación previos a la conservación de las semillas en bancos de germoplasma.
De acuerdo con estas premisas, los objetivos del trabajo fueron: 1) determinar la forma en que la temperatura y la luz del sustrato afectan la germinación de las plantas arbustivas y trepadoras de un bosque siempreverde tropical de la RBSR, 2) identificar las clases de dormancia seminal, y 3) determinar las relaciones que se establecen entre las clases de dormancia y otros rasgos de historia de vida.
MATERIALES Y MÉTODOS
Descripción del sitio. La RBSR se localiza en la parte más oriental de la Sierra del Rosario, Artemisa, Cuba (22º 45' N, 82º 50' O) y tiene una extensión de 25 000 ha. En la región, la temperatura media anual del aire es de 24,4 ºC. Los valores de temperatura mínima y máxima para el suelo del sotobosque a 2 cm de profundidad durante el verano (mayo-octubre) son de 22,3 y 24,5 ºC; mientras que para un claro grande en el dosel son de 22,5 y 34,5 ºC, respectivamente (Herrera et al., 1988). La precipitación promedio anual es de 2 300 mm, y, de acuerdo con la distribución de la lluvia y la propuesta de Sánchez et al. (2009), se reconocieron tres estaciones de dispersión/colecta: estación seca (DS, diciembre-marzo), comienzo de la estación lluviosa (ERS, abril-julio) y final de la estación lluviosa (LRS, agosto-noviembre).
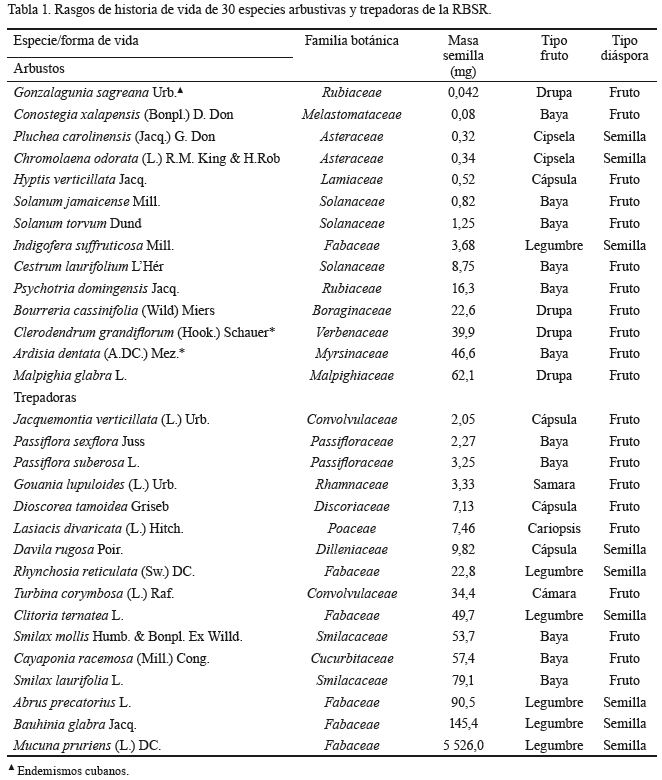
Especies seleccionadas y colecta de semilla. Las plantas se seleccionaron sobre la base de su frecuencia en el sitio de estudio y la disponibilidad de semillas. Según Herrera et al. (1988), las especies pertenecen a la flora primaria de los bosques de la RBSR (tabla 1), excepto Indigofera suffruticosa, que es una especie naturalizada para este sitio. Las especies se ordenaron a partir del valor promedio de la masa fresca seminal dentro de cada forma de vida (Montejo et al., 2014a), la cual mostró un amplio rango de valores (0,042-5 526,0 mg). Los nombres de las plantas se establecieron de acuerdo con lo informado por Acevedo-Rodríguez y Strong (2012) y en el caso de las familias botánicas se siguió el The Angiosperm Phylogeny Group (2003). Los taxones pertenecían a 18 familias botánicas, las más representadas fueron: Fabaceae (6) y Solanaceae (3). Se colectaron semillas frescas de 14 especies arbustivas y 16 trepadoras (13 enredaderas y tres lianas).
El tipo de diáspora (unidad de dispersión) de 21 especies fue el fruto. En este estudio, se consideró como semilla una verdadera semilla y como diáspora, una verdadera semilla más el endocarpo. Las especies cuya diáspora incluyó el endocarpo de las drupas fueron: Gonzalagunia sagreana, Bourreria cassinifolia, Clerodendrum grandiflorum, Malpighia glabra y el endocarpo de la cámara se observó en Turbina corymbosa.
Las semillas se colectaron en la Estación Ecológica El Salón (sección central de la RBSR), cubierta por un bosque siempreverde estacional submontano, con un dosel cerrado de 30 m de altura y un 13 % de árboles que pierden sus hojas durante los meses de menor precipitación. Todos los frutos de cinco o más individuos saludables se tomaron directamente de la planta madre, con una vara telescópica, y se limpiaron inmediatamente. El secado de las semillas se realizó durante 72 h, al aire y a la sombra.
Requerimientos germinativos. Las semillas de cada especie se mezclaron y se sembraron durante los tres primeros días, después de su limpieza. Los estudios de germinación se realizaron en incubadoras, con luz y temperatura controladas. Se utilizó un diseño completamente aleatorizado, con arreglo factorial de los tratamientos, el cual simuló las variaciones de luz y de temperatura que sufre el suelo del bosque desde un claro hasta su interior. Se probaron cuatro condiciones de temperatura y dos de iluminación, para un total de ocho tratamientos por especie. Las respuestas germinativas se obtuvieron a temperatura constante de 25 ºC y en un régimen de temperatura alterna de 25/30 ºC, 25/35 ºC y 25/40 ºC (8 h para la temperatura más alta del termoperiodo y 12 h a 25 ºC, con una transición de 4 h entre estas). Los tratamientos de iluminación fueron: exposición a la luz (8 horas de aproximadamente 40 mmol m-2 s-1, longitud de 400-700 nm), coincidente con el periodo de mayor temperatura dentro de cada termoperiodo; y oscuridad constante. Esta última condición se logró envolviendo las placas en dos capas de papel de aluminio. La localización de las placas en la incubadora se cambió regularmente.
Antes de la siembra, las semillas se esterilizaron con bicloruro de mercurio (1 g L-1), para prevenir la contaminación fúngica. Se utilizaron cinco réplicas de 25 semillas cada una por tratamiento. La siembra se realizó sobre agar hidrostático al 1 %, en placas Petri de 9 o 15 cm de diámetro, en dependencia del tamaño de la semilla. En el caso de las semillas que fueron iluminadas, el conteo de germinación se efectuó diariamente durante 30 a 90 días, en dependencia de la especie. En las semillas expuestas a condiciones continuas de oscuridad total el conteo se realizó tres días después de haber concluido la germinación a la luz. El criterio para considerar las semillas germinadas fue la protrusión de la radícula. En los tratamientos a la luz, el agar se mantuvo húmedo durante el experimento.
Se determinó el porcentaje de germinación final a la luz y a la oscuridad. En las semillas germinadas a la luz se estableció, además, el día de inicio de la germinación (DIG) y el coeficiente de velocidad de germinación (CVG). El CVG se calculó para cada réplica, dividiendo el número final de semillas germinadas (N) entre la sumatoria del número de semillas germinadas en un día particular (ni), multiplicado por el número de días correspondiente (di) desde el comienzo del experimento (Alm, Stoller y Wax, 1993): CVG = N/Sni di.Los valores de CVG se encontraban entre 0 (no germinaci ón) y 1 (rápida velocidad de germinación), y se multiplicaron por 100 para facilitar la interpretación.
El porcentaje de germinación final a la luz y a la oscuridad se empleó para calcular el índice de germinación relativa a la luz (GRL), que expresó los requerimientos de luz para la germinación (Milberg, Andersson y Thompson, 2000). Este índice fue calculado según la fórmula: GRL = GL/(GD + GL), donde GL: porcentaje de germinación a la luz, y GD: porcentaje de germinación a la oscuridad. Para obtener el índice de GRL de cada especie se utilizó el rango de temperatura óptimo (i.e., aquel en el que la especie mostró el mayor porcentaje de germinación a la luz o a la oscuridad). Los valores de GRL variaron entre 0 (semillas que germinan solo a la oscuridad) y 1 (semillas que germinan solo a la luz).
En el caso de las semillas que no germinaron, se determinó la viabilidad mediante la prueba de TZ (ISTA, 1999); o presionando las semillas con una aguja para comprobar si contenían un embrión blanco y firme (semillas vivas) o uno gris y suave (ausencia de viabilidad seminal).
Asignación de las clases de dormancia. Se consideró que una especie poseía semillas dormantes si: a) su germinación comenzó después de 28 días en condiciones óptimas (Baskin y Baskin, 1998), b) las semillas tuvieron embriones subdesarrollados y diferenciados (i.e., el embrión debe crecer dentro de la semilla), independientemente del tiempo para iniciar la germinación; c) al menos el 20 % de las semillas permanecieron vivas y sin germinar. Las semillas se consideraron no dormantes si el embrión se desarrolló completamente y estas requirieron menos de 28 días para germinar (Baskin y Baskin, 1998).
Las especies con semillas dormantes se clasificaron según el sistema de clases de dormancia seminal propuesto por Baskin y Baskin (2004), el cual reconoce cinco clases: 1) dormancia fisiológica (PD), 2) dormancia morfológica (MD), 3) dormancia morfofisiológica (MPD), 4) dormancia física (PY), y 5) dormancia combinada (PY + PD). Las semillas que tienen dormancia combinada necesitan más de 28 días para germinar, después de ser eliminada la PY.
La capacidad de absorción de agua (imbibición) de las semillas frescas se determinó en aquellas especies cuyos porcentajes de germinación fueron bajos, o bien en las que se reporta PY en la familia. Para ello, se utilizaron cinco réplicas de 25 semillas por especie, que se colocaron sobre dos capas de papel de filtro humedecido con agua destilada en placas Petri. Las pruebas se realizaron bajo luz blanca y a temperatura óptima de germinación de cada especie (como se describió anteriormente). A las 24, 48 y 72 horas de imbibición, las semillas se secaron superficialmente y se pesaron. El porcentaje de absorción de agua se determinó en relación con el peso fresco de las semillas.
En los casos en que los porcentajes de germinación fueron bajos y el número de semillas colectadas fue suficiente, se aplicaron además tratamientos pregerminativos, con el objetivo de romper la dormancia seminal. Estos tratamientos fueron: escarificación mecánica total (EMT) en Ardisia dentata, B. cassinifolia, Clerodendrum grandiflorum y Cestrum laurifolium, y ruptura de la testa en Mucuna pruriens. Los ensayos se realizaron en las condiciones óptimas de germinación de cada especie, que fueron establecidas en las pruebas de germinación sin tratamientos. Se determinó el porcentaje de germinación final y el día de inicio de la germinación.
Finalmente, para asignar las clases de dormancia no solo se emplearon las metodologías previamente referidas, sino también la información sobre el tipo de embrión reportado para la especie/familia y las características morfológicas de las semillas, debido a que la dormancia seminal es un rasgo filogenético altamente conservado (Baskin y Baskin, 2005; Kos, Baskin y Baskin, 2012).
Rasgos de historia de vida por clase de dormancia. Las variables seminales comparadas fueron: la masa fresca de la semilla (mg), el contenido de humedad inicial (%), el porcentaje de masa seca de la semilla destinado a las cubiertas seminales, el índice de GRL, el DIG y el porcentaje de germinación final (GF). Los rasgos de las semillas se tomaron de lo informado por Montejo et al. (2014a), y los rasgos germinativos, del presente estudio. También se determinaron las clases de dormancia según el tiempo y la forma de dispersión de las semillas. Para asignar los síndromes de dispersión las especies se agruparon en dos categorías: las que fueron dispersadas por los animales (incluye la dispersión interna o externa) y las no dispersadas por estos animales.
Análisis estadístico. Los rasgos de germinación que se establecieron a nivel de comunidad y por forma de vida no cumplieron las premisas de los supuestos paramétricos; por tanto, para detectar posibles interacciones entre los factores (temperatura por luz), se aplicó un análisis de varianza multivariado sobre la base de permutaciones PERMANOVA (Anderson, 2001). Los PERMANOVA se realizaron con el valor medio de cada especie y por una matriz de distancia euclidiana, después de 9 999 interaciones. Para investigar la relación entre la masa de la semilla (tamaño seminal) y el índice de GRL, se empleó un análisis de regresión lineal simple. Las diferencias entre los tiempos de imbibición y los tratamientos pregerminativos se detectaron por medio de un ANOVA de clasificación simple, debido a que los datos cumplieron las premisas de este análisis. Se utilizó, además, un análisis de similitud (ANOSIM) de una vía, para evaluar la hipótesis Ho de no diferencias estadísticamente significativas entre rasgos de historia de vida por clases de dormancia. Cada ANOSIM se realizó con una matriz de distancia euclidiana y usando 9 999 permutaciones.
RESULTADOS
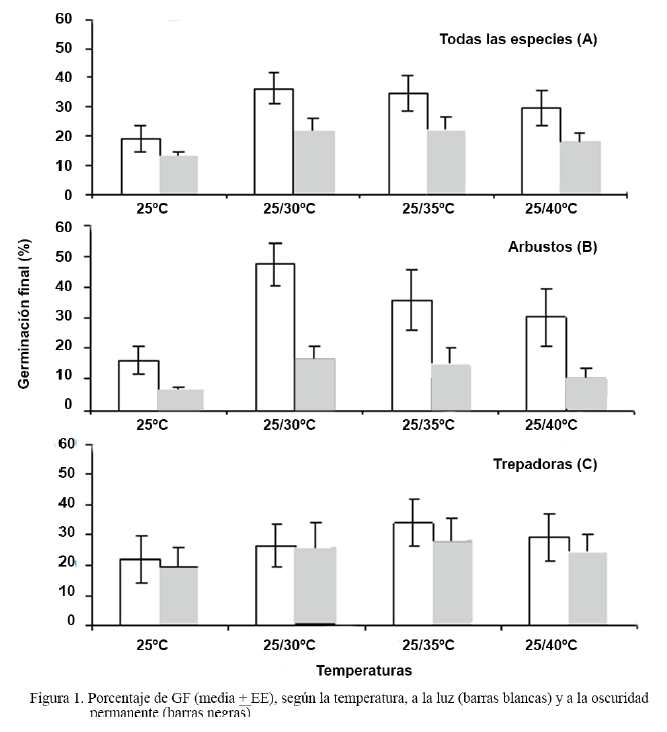
Requerimientos germinativos. La germinación de las 30 especies se afectó significativamente por la temperatura (F (3, 232) = 2,82; p < 0,05) y la luz del sustrato (F (1, 232) = 11,07; p < 0,001), aunque no se observó interacción entre estos factores (p = 0,83). La luz fue el factor más importante, por lo que las condiciones de oscuridad afectaron considerablemente la germinación (fig. 1A). Al considerar el total de especies, el porcentaje de GF a 25/30 ºC y 25/35 ºC fue superior que a temperatura constate (25 ºC), tanto en la luz como en la oscuridad permanente; pero no difirió del obtenido a 25/40 ºC. En los arbustos, este patrón general se repitió (fig. 1B), o sea, la germinación dependió significativamente de la temperatura (F (3, 104) = 4,05; p < 0,01) y de la luz del sustrato (F (1, 104) = 21,90; p < 0,001), pero no de la interacción de estos factores (p = 0,91). El incremento de la germinación en los arbustos por la luz fue más evidente en el rango de 25/30 ºC, aunque no difirió de lo obtenido a 25/35 ºC y 25/40 ºC, pero sí a 25 ºC. En las trepadoras, el ANOVA no mostró diferencias en la germinación final (datos no mostrados), aunque el máximo valor promedio se encontró a 25/35 ºC bajo luz blanca (fig.1C).
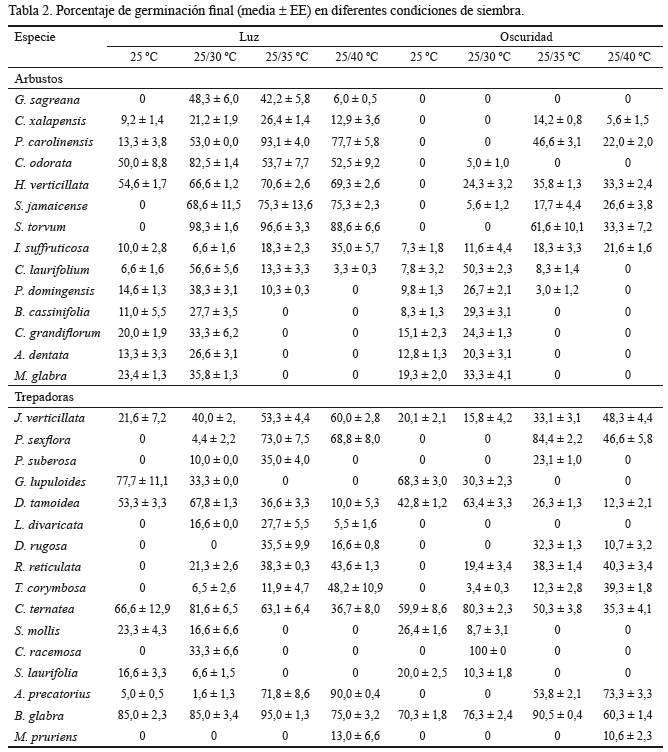
El patrón para todos los taxones también se repitió individualmente en muchas especies de arbustos, así como en las trepadoras (tabla 2). Sin embargo, en siete especies de arbustos (G. sagreana, Conostegia xalapensis, Pluchea carolinensis, Chromolaena odorata, Hyptis verticillata, Solanum jamaicense y Solanum torvum) que provenían de semillas pequeñas (menos de 1,3 mg, tabla 1) la germinación fue nula, cuando la siembra se realizó en condiciones de oscuridad total y a 25 ºC. Asimismo se evidenció, independientemente de la condición de iluminación del sustrato, que la mejor respuesta germinativa en 11 especies (36,6 %) ocurrió a 25/30 ºC; mientras que en 9 especies (30,0 %) se manifestó a 25/35 ºC. A 25 ºC, la mejor germinación (10,0 %) solo se alcanzó en 3 especies trepadoras (Gouania lupuloides, Smilax mollis y Smilax laurifolia) y a 25/40 ºC en 7 especies (23,3 %), de ellas dos arbustos (S. jamaicense e I. suffruticosa) y 5 trepadoras (Jacquemontia verticillata, Rhynchosia reticulata, T. corymbosa, Abrus precatorius y M. pruriens).
No existió un patrón claro entre la masa fresca de la semilla (tamaño) y el porcentaje de GF, aunque esta última variable tendió a ser mínima cuando se incrementó la alternancia de temperatura del sustrato y el tamaño de las semillas. Este efecto fue mayor en los arbustos, tanto en condiciones de luz como en oscuridad total (tabla 2).
El CVG y el DIG no mostraron diferencias significativas (p = 0,45 y p = 0,79, respectivamente) en las distintas temperaturas, según los resultados del ANOVA, pero estas no evidenciaron las verdaderas diferencias provocadas por las temperaturas del sustrato en la velocidad del proceso germinativo (tabla 3). Solo se registraron el CVG y el DIG para 11 especies de las 30 estudiadas. Los mayores valores del CVG (rápida velocidad de germinación) y los menores del DIG (menor tiempo para iniciar la germinación) se obtuvieron a la temperatura óptima de germinación de cada especie.
En las condiciones óptimas de siembra, el porcentaje de GF entre las especies varió desde 13,0 % en M. pruriens hasta 96,6 % en S. torvum; 13 especies (que representaron el 43,3 %) alcanzaron más de 50 % de germinación y solo 4 (13,3 %) lograron menos del 30 % en esta variable. El CVG (%) osciló de 0,7 en M. pruriens hasta 26,9 en C. odorata, y el DIG abarcó desde 2,2 días en C. odorata hasta 97,3 días en D. rugosa (tabla 3). En los arbustos, el CVG y el DIG mostraron valores medios de 7,2 ± 1,8 % y 23,3 ± 6,5 días, respectivamente; mientras que en las trepadoras estas variables fueron de 3,5 ± 0,7 % y 34,8 ± 2,8 días, respectivamente.
El porcentaje de semillas vivas no germinadas (o dormantes), en todas las especies, solo se afectó significativamente por la temperatura del sustrato (F (3, 231) = 2,93, p < 0,05); y cuando se realizó el análisis por forma de vida ninguno de los factores probados ni su interacción afectaron significativamente dicha variable (datos no mostrados).
El porcentaje de semilla dormante, en el rango de temperatura óptimo para la germinación de cada especie, también varió considerablemente, desde 0 % en diversas especies hasta 83 % en S. laurifolia (tabla 4). Sin embargo, los valores nulos en algunas especies (e.g., G. sagreana, Psychotria domingensis, B. cassinifolia, C. grandiflorum, A. dentata, M. glabra, Passiflora suberosa y Lasiacis divaricata) y muy bajos en otras no representaron verdaderamente el grado de dormancia; en estas especies las condiciones de siembra afectaron la viabilidad de las semillas que no lograron germinar.
El GRL, que representó la relación entre el porcentaje de germinación a la luz y a la oscuridad, varió desde 0,24 hasta 1. Este último valor lo alcanzaron solo dos especies (G. sagreana y L. divaricata), que se podrían considerar como dependientes de la luz para germinar o fotoblásticas positivas; y también pueden incluirse en este grupo las semillas de C. odorata y S. jamaicense por sus altos valores de GRL (0,94 y 0,73, respectivamente). En general, estas especies presentaron semillas pequeñas (con un peso menor que 2,5 mg). Asimismo, aparecieron 9 especies (C. xalapensis, P. carolinensis, H. verticillata, S. torvum, I. suffruticosa, P. domingensis, C. grandiflorum, A. dentata y P. suberosa) que germinaron mejor a la luz que a la oscuridad, pero los valores de GRL no fueron tan altos (0,56-0,67). Por su parte, la única especie que presentó un valor de GRL menos o igual que 0,40 fue Cayaponia racemosa (0,24), taxón que prefiere la oscuridad para germinar; aunque curiosamente las otras dos lianas estudiadas (S. mollis y S. laurifolia: 0,46 y 0,45, respectivamente) y la enredadera Pasiflora sexflora (0,46) también presentaron valores bajos del índice de GRL, que se ubicaron dentro del rango de especies indiferentes a la luz, pero tendieron a germinar mejor en la oscuridad (tabla 2).
El resto de las especies (13-43 % del total) germinaron de forma similar en condiciones de luz o de oscuridad (indiferentes a la luz); los valores de GRL variaron desde 0,48 hasta 0,55, y la masa seminal, desde 2,0 hasta 5 226,0 mg; aunque, como promedio, las que integraron este grupo presentaron una masa fresca seminal de 382,5 mg. En todas las especies, el índice de GRL se relacionó negativamente con la masa fresca de la semilla (fig. 2; R2= 0,339, p < 0,001), es decir, la respuesta germinativa a la luz disminuyó con el aumento de la masa seminal.
Asignación de las clases de dormancia. De las 30 especies estudiadas, 23 (76,6 %) poseían semillas dormantes. De estas, 10 eran arbustos (43,47 %) y 13 trepadoras (56,53 %). Los taxones con semillas no dormantes (P. carolinensis, C. odorata, S. jamaicense, S. torvum, G. lupuloides, Clitoria ternatea y Bauhinia glabra) presentaron como promedio 85,9 ± 8,7 % de GF en el rango de temperatura óptimo para cada especie, y este proceso comenzó, por lo general, en menos de ocho días (tablas 2 y 3). De las plantas con semillas dormantes, 15 especies (65,2 %, en relación con el total de dormantes) G. sagreana, P. domingensis, B. cassinifolia, C. grandiflorum, A. dentata, M. glabra, P. sexflora, P. suberosa, Dioscorea tamoidea, D. rugosa, S. mollis, S. laurifolia, C. racemosa, A. precatorius y M. pruriens necesitaron más de 28 días para iniciar la germinación, y 7 (30,4 %) C. xalapensis, H. verticillata, I. suffruticosa, C. laurifolium, J. verticillata, R. reticulata y T. corymbosa finalizaron el experimento con más de 20 % de semillas vivas no germinadas. Las semillas de D. tamoidea, S. mollis y S. laurifolia mostraron embriones subdesarrollados, y las de L. divaricata alcanzaron su máxima germinación (27,7 %) en un rango de temperatura (25/35 ºC) que no existía en el suelo cuando se dispersaron, por lo que se consideraron dormantes.
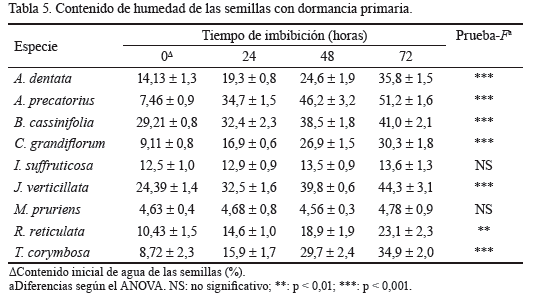
Las pruebas de imbibición de 9 especies con dormancia primaria demostraron que las semillas de todas, excepto I. suffruticosa y M. pruriens, se hidrataron después que estuvieron 24 h en contacto con el agua (tabla 5). Sin embargo, las pruebas de corte de semillas frescas de J. verticillata, R. reticulata, T. corymbosa y A. precatorius mostraron que al menos 30, 40, 20 y 10 %, respectivamente, no se hidrataron al final del experimento de germinación.
El tratamiento de EMT fue efectivo para incrementar el porcentaje de GF en 3 de las 4 especies que se probaron (tabla 6); aunque en C. laurifolium aumentó la GF, pero no la velocidad de dicho proceso. En las semillas de A. dentata este tratamiento pregerminativo no fue efectivo, y en semillas frescas de M. pruriens la ruptura de la testa incrementó y aceleró la germinación.
Se puede resumir que 14 especies presentaron dormancia fisiológica como único impedimento para su germinación; 6, dormancia física (en al menos un fracción de las semillas del lote), y 3, dormancia morfológica y/o morfofisiológica. No se identificó ninguna especie con combinación de dormancia. En los arbustos predominó la PD (9 especies) y en las trepadoras PD y PY se distribuyeron de manera equitativa, es decir, 5 especies presentaron PD y 5, PY. En las trepadoras también se encontraron las únicas especies con embriones subdesarrollados y, por tanto, con MD + MPD o MPD.
Rasgos de historia de vida por clases de dormancia. El porcentaje de GF y el DIG mostraron variabilidad significativa según la clase de dormancia (tabla 7). Las semillas con DM y/o MPD presentaron los menores valores de GF y los máximos del DIG, y las semillas no dormantes, los máximos de GF y los menores del DIG. En los otros rasgos seminales no se detectaron diferencias significativas por clase de dormancia, debido, posiblemente, a la gran variabilidad que tuvieron.
Por otra parte, el porcentaje de especies con semillas dormantes varió con el tiempo de dispersión (tabla 8). Las semillas de 15, 13 y 2 especies se colectaron durante DS, ERS y LRS, respectivamente. Las especies dispersadas en la DS presentaron semillas más dormantes (87 %, en relación con el total colectado en la estación) que aquellas colectadas al ERS (77 %), y no tuvieron semillas dormantes las colectadas en LRS. La mayoría de las especies dispersadas en la ERS y la DS presentaron PD (38,4 y 60,0 %, respectivamente) y de las 6 especies con PY, 4 se encontraron en la DS. De las 15 especies que fueron dispersadas por los animales, 13 tuvieron dormancia (86,6 %), 11 con PD (73,3 %) y 2 con MD o MPD (13,3 %). En cambio, en las no dispersadas por los animales, la dormancia estuvo presente en 10 especies de 15, y predominó la dormancia física (40 %).
DISCUSIÓN
La temperatura y la luz del sustrato afectaron la germinación en todas las especies (por comunidad), y la iluminación fue el factor principal que reguló dicho proceso. Según Pons (2000) y Pearson et al. (2002), en el dosel del bosque, el principal factor limitante de la germinación suele ser la luz que llega al suelo, ya que esta disminuye en cantidad y calidad por la vegetación. Tal resultado fue más evidente cuando se analizó la germinación por forma de vida. En la mayoría de los arbustos, esta variable fue estimulada por la luz, fundamentalmente en el rango de 25/30 ºC, pero también a 25/35 ºC y 25/40 ºC. Dicho comportamiento se ha informado para un gran número de especies arbóreas pioneras del Neotrópico (Vázquez-Yanes y Orozco-Segovia, 1994; Pearson et al. 2002), así como en Cuba, para este mismo grupo ecológico del área de estudio (Sánchez et al., 2003; Muñoz et al., 2012). Las observaciones en el sitio de colecta evidenciaron que muchas de las plántulas de arbustos se encontraron indistintamente en sitios abiertos o semiabiertos.
La respuesta germinativa de los arbustos en los rangos de temperatura ensayados demostró que la germinación estuvo condicionada también por la alternancia de temperatura del sustrato, como sucede en los claros del bosque (Vázquez-Yanes y Orozco-Segovia, 1994). De hecho, la gran mayoría de los arbustos (estudiados o no en las pruebas de germinación; Sánchez, J.A., inédito) dispersaron sus semillas durante el comienzo de las lluvias o estación de verano para Cuba (48,8 %); por lo que se evidenció que ello está condicionado por una variabilidad espacial (estructura abierta de la vegetación) y temporal (concentración de lluvia e incremento de las temperatura), probablemente para maximizar el establecimiento y la supervivencia de los individuos (Baskin y Baskin, 1998).
En las trepadoras se apreció una gran variabilidad interespecífica en relación con la temperatura, por lo que no se detectó preferencia germinativa por un rango de temperatura en particular (fig. 1C); aunque es necesario señalar que en las especies con PY (I. suffruticosa, J. verticillata, R. reticulata, T. corymbosa, A. precatorius y M. pruriens) la mejor respuesta germinativa se obtuvo a 25/40 ºC (tabla 2). Las condiciones de oscuridad total incrementaron significativamente la germinación en cuatro trepadoras (P. sexflora, S. mollis, S. laurifolia y C. racemosa); la mejor respuesta en S. mollis y S. laurifolia a 25 ºC y en condiciones de oscuridad evidenció la adaptación de estos taxones para germinar en el interior del bosque, donde la temperatura del aire se mantiene relativamente constante durante casi todo el año 25 ± 2 ºC (Herrera et al., 1988) y la iluminación que llega al suelo es pobre en rojo y rica en rojo lejano (Vázquez-Yanes y Orozco-Segovia, 1990). De estas cuatro especies, la única que tendió a ser fotoblástica negativa fue C. racemosa, pues alcanzó los valores más bajos de GRL (0,24). Probablemente este comportamiento sea normal, ya que dicha planta crece en sitios semicerrados del bosque; sin embargo, su mejor respuesta germinativa a 25/30 ºC no apoya totalmente esta hipótesis.
La respuesta germinativa de las semillas frescas de las plantas trepadoras mostró gran plasticidad, ya que no se evidenció preferencia germinativa marcada por los factores probados. Sanches y Válio (2002) también encontraron un resultado similar en 13 trepadoras, en un bosque tropical de Brasil, las cuales germinaron y se establecieron en claros, en sitios semisombreados o en el interior del bosque.
A través de todas las especies se evidenció que la luz, como señal para la germinación, fue menos importante en aquellas con semillas grandes que en las que poseían semillas pequeñas (fig. 2). Este patrón fue detectado por primera vez, a nivel de comunidad, por Grime et al. (1981) en plantas de regiones templadas. Milberg, Andersson y Thompson (2000) observaron un comportamiento similar en 54 especies herbáceas, con o sin control filogenético, por lo que concluyeron que los requerimientos de luz para la germinación han coevolucionado con la masa seminal. Recientemente Flores et al. (2011) obtuvieron, en semillas de cactáceas de siete países, una relación negativa entre el índice de GRL y la masa de la semilla; ello coincide con lo obtenido por Sánchez et al. (2012) en 60 árboles pertenecientes al sitio de estudio. Es importante señalar que el coeficiente de determinación (R2) entre el índice de germinación a la luz y el logaritmo de la masa fresca de la semilla en el presente estudio fue bajo (R2 = 0,339), y similar al encontrado por Milberg, Andersson y Thompson (2000) R2 = 0,277 y Sánchez et al. (2012) R2 = 0,280, lo que demostró que en ambas floras la respuesta germinativa a la luz varió grandemente, y no toda la variación se debió a la masa de las semillas. De hecho, esta variable solo explicó el 33 % de la variación del índice de GRL. Diversos autores han planteado que la fotosensibilidad es un fenómeno complejo, en el que se involucran rasgos intrínsecos de cada semilla y su relación con el ambiente edafoclimático (Baskin y Baskin, 1998; Pons, 2000).
En el pequeño grupo de arbustos y trepadoras se encontró un alto porcentaje de especies con semillas dormantes, tanto en la estación seca (87 %) como al comienzo de las lluvias (77 %), el cual sobrepasa los valores de dormancia informados para estas formas de vida en los bosques húmedos estacionales de los trópicos (Baskin y Baskin, 1998). De hecho, la poca información que existe sobre la dormancia de las trepadoras y los arbustos indica que la mayoría de estas especies presentan semillas no dormantes cuando ocupan sitios húmedos, y en el caso de que sean dormantes, en las trepadoras predominan la PD, la PY y la MPD, y en los arbustos la PD y la PY (Baskin y Baskin, 1998). Sin embargo, de los 14 arbustos, 10 especies presentaron dormancia (71,4 %); con predominio de la clase fisiológica. Según Baskin y Baskin (1998), esta clase de dormancia es la más frecuente para todos los tipos de vegetación y formas de vida (árboles, arbustos, enredaderas y hierbas) de las zonas tropicales y subtropicales del mundo. También se ha informado que es la más importante para todos los clados de las angiospermas (Baskin y Baskin, 2003; Silveira et al., 2012).
Este resultado se contrapone a los modelos teóricos que se han propuesto a escala global, y que presuponen que el número de especies con estrategias de aplazamiento de la germinación (i. e., dormancia seminal) aumenta en los ambientes no favorables respecto a aquellas que ocupan hábitats favorables (Jurado y Moles, 2003; Baskin y Baskin, 2005; Jurado y Flores, 2005); aunque también se conoce que estos estudios no permiten concluir con precisión acerca de los procesos que intervienen en el ensamblaje (o matriz de especies) de las comunidades locales (Bu et al., 2008; Kos, Baskin y Baskin, 2012). De hecho, muchas de las especies que dispersan sus semillas en la llamada época óptima de crecimiento (ERS) de estos bosques estacionales tienen dormancia seminal (Sautu et al., 2007; Sánchez et al., 2012); por tanto, no parece ser la dormancia, en esta estación del año, una estrategia de aplazamiento para esperar la llegada de condiciones ambientales más adecuadas (incremento de la temperatura y la precipitación) para germinar. Más bien parece ser una estrategia para evitar la depredación del cohorte de plántulas, aunque también podría evitar procesos de sequía dentro de la estación lluviosa. El efecto de este último escenario ecológico sobre la mortalidad de las plántulas en los bosques húmedos de la RBSR fue documentado por Herrera et al. (1988), y por Engelbrecht et al. (2006) para especies pioneras de bosques de Panamá.
También los resultados demostraron que las semillas de las trepadoras tuvieron mayor retardo para iniciar la germinación respecto a las de los arbustos, lo que quizás se deba a que las primeras presentaron el mayor número de especies con PY y las tres únicas especies con MD y/o MPD (Baskin y Baskin, 2004). Igualmente, la mayoría de estas especies se dispersaron durante la estación seca; por tanto, la dormancia en este grupo de plantas podría ser una estrategia de aplazamiento hasta la llegada de las condiciones óptimas para germinar y establecerse las plántulas (Jurado y Moles, 2003; Salazar et al., 2011). Este comportamiento germinativo se corresponde con el síndrome de germinación encontrado por Garwood (1983) y Sautu et al. (2007) para las especies arbóreas de los bosques estacionales de Panamá y para los árboles del sitio de estudio (Sánchez et al., 2012).
De acuerdo con los resultados de las pruebas de imbibición, corte y germinación se confirmó que las semillas frescas de A. precatorius, I. suffruticosa, J. verticillata, M. pruriens, R. reticulata y T. corymbosa presentaron PY como principal impedimento para la germinación, tal como se ha informado previamente para las familias de estas especies Fabaceae y Convolvulaceae (Baskin, Baskin y Li, 2000; Gehan et al., 2012) y para algunas de las especies estudiadas (Brancalion et al., 2010).
En las semillas intactas de A. precatorius, M. pruriens, R. reticulata y T. corymbosa los mayores porcentajes de GF se obtuvieron a temperaturas alternas de 25/40 ºC y 25/35 ºC. Sin embargo, estas temperaturas no son las que encuentran las semillas cuando caen al suelo en la estación de seca (invierno para Cuba), pues dichos rangos solo se pueden alcanzar en la estación lluviosa. Así, fue evidente que la escarificación en caliente aceleró la ruptura de la dormancia de, al menos, una porción de las semillas; este tratamiento es ampliamente reconocido para eliminar la PY (Baskin y Baskin, 2004). En semillas de M. pruriens, con el tratamiento de ruptura de la testa y siembra a 25/40 ºC, se logró incrementar la germinación hasta 100 %, resultado que también se obtuvo para esta especie a 25/30 ºC y 25/35 ºC (datos no mostrados) y coincide con lo expresado por Baskin y Baskin (1998) en relación con la amplia capacidad germinativa que muestran las semillas con PY después que son sometidas a escarificación. Las semillas de A. precatorius presentaron la mejor respuesta germinativa al incrementarse la temperatura del sustrato, y lograron un 90 % de GF a 25/40 ºC. Sin embargo, a temperatura constante de 25 ºC y a 25/30 ºC más del 90 % de ellas no se hidrataron al final del experimento. Esta especie presenta mimetismo simulado y la PY le permite sobrevivir al deterioro seminal que sufren las semillas durante su paso por el tracto digestivo de los dispersores (Brancalion et al., 2010). Sin embargo, su mecanismo de dispersión primario es la autocoria, por tanto, la PY también podría servirle para permanecer viable en el suelo.
En las semillas de I. suffruticosa (Fabaceae) y J. verticillata (Convolvulaceae), los porcentajes de germinación que se alcanzaron a 25/40 ºC no fueron tan altos (35 y 60 %, respectivamente); por tanto, hubo una fracción importante del lote que al parecer tuvo una mayor profundidad de la PY, posiblemente como mecanismo para extender la germinación durante las lluvias, pues las semillas llegaron al suelo al comienzo de la temporada lluviosa (abril). Sin embargo, el mecanismo propuesto podría no funcionar para I. suffruticosa, ya que la especie no es nativa de la RBSR.
Las pruebas de imbibición indicaron que la PY no constituyó el principal obstáculo para la germinación en las semillas de A. dentata, B. cassinifolia y C. grandiflorum. En las familias de estas especies tampoco se ha reportado la presencia de embriones subdesarrollados (Montejo et al., 2014b), por lo que se asume que no presentaron MD o MPD ni la combinación de ninguna de ellas con la PY; entonces solo pueden tener PD, que es la dormancia más común en algunas de dichas familias (Baskin y Baskin, 1998). Un análisis similar se puede hacer para las semillas de G. sagreana, C. xalapensis, H. verticillata, C. laurifolium, P. domingensis, M. glabra, P. sexflora, P. suberosa, L. divaricata, D. rugosa y C. racemosa, aunque no se realizaron pruebas de imbibición. En las familias de estas especies los embriones están desarrollados y nunca se ha reportado PY, excepto en C. racemosa Cucurbitaceae (Baskin, Baskin y Li, 2000); sin embargo, sus semillas se hidrataron desde el comienzo de las pruebas de germinación. También se ha sugerido que podría existir PY en Passifloraceae por la anatomía de sus cubiertas (Baskin, Baskin y Li, 2000), pero las semillas de P. sexflora y P. suberosa se hidrataron muy rápidamente; por tanto, para estas últimas 11 especies igualmente se asume que presentaron PD, lo cual se debe a mecanismos de inhibición fisiológica en el embrión (Baskin y Baskin, 1998).
Las semillas frescas de los arbustos endémicos cubanos (A. dentata, C. grandiflorum y G. sagreana) llegaron al suelo en la estación seca y mostraron PD, como ya se comentó; sin embargo, la profundidad de esta dormancia cambió con las especies. Las de C. grandiflorum incrementaron y aceleraron significativamente la germinación final con la EMT; lo que sugiere que deben presentar PD no profunda, como se reporta para la familia Verbenaceae (Baskin y Baskin, 1998). Según Baskin y Baskin (2004), el tratamiento pregerminativo de EMT es adecuado para eliminar este tipo de dormancia. Sin embargo, este no fue efectivo en A. dentata: una fracción del lote de semillas (30 %) debió presentar PD no profunda porque germinaron después de 28 días, y la otra fracción que no germinó (70 %) aun con el tratamiento pregerminativo es probable que presentara PD intermedia o profunda. Estas semillas de A. dentata llegan al suelo a finales de la estación seca (marzo) y son ingeridas por los animales, por lo que es muy probable que la PD les permita sobrevivir hasta la llegada de las lluvias, o bien durante su paso por el tracto digestivo, tal como se ha informado para un gran número de especies que son dispersadas por los animales (Bu et al., 2008).
Las semillas de G. sagreana iniciaron la germinación después de los 36 días y no llegaron a alcanzar más de 50 % de germinación final a la temperatura óptima (25/30 ºC), por lo que presentaron PD, como se ha informado para la familia Rubiaceae (Baskin y Baskin, 1998). Los resultados evidenciaron además que al menos una fracción del lote (50 %) debió presentar PD intermedia o bien profunda, pues al cabo de los 80 días de su siembra aún esa fracción no había germinado. Esta porción de semillas probablemente permanezca viable en el suelo hasta el comienzo de las lluvias, pero tal vez podrían formar un banco de semilla más estable, pues fueron las más pequeñas (0,042 mg), fotoblásticas positivas, y destinaron más de 40 % de su masa seca a las estructuras de defensa física (testa/endocarpo); rasgos morfofisiológicos que permiten formar un banco de semillas (Thompson et al., 2003) y, por tanto, responder a los disturbios. Estas características se presentan en especies arbóreas pioneras tempranas de los bosques cubanos (Muñoz et al., 2012). De hecho, G. sangreana se considera una planta nativa expansiva (Ricardo y Herrera, 2010).
Por su parte, las semillas de H. verticillata y L. divaricata germinaron en menos de 28 días, pero alcanzaron su máxima germinación final (70,6 y 27,7 %, respectivamente) en un rango de temperatura que no encuentran cuando llegan al suelo (25/35 ºC), durante la estación de seca; por tanto, este rango aceleró la posmaduración de al menos una fracción del lote. Al parecer la PD fue más profunda en la segunda especie, pues la germinación resultó muy baja. Las semillas de L. divaricata fueron además fotoblásticas positivas, lo que sugiere que podrían permanecer viables en el suelo por un tiempo, como previamente se discutió para G. sagreana. También las semillas frescas de C. xalapensis (Melastomataceae) y C. laurifolium (Solanaceae) germinaron en menos de 28 días, pero dejaron un alto porcentaje de dormantes en la mejor temperatura de siembra (73,6 y 26,6 %); por lo que al menos esas fracciones presentaron PD, que es la dormancia informada en sus familias (Baskin y Baskin, 1998; Silveira et al., 2012). Probablemente, la PD en estas especies sea un mecanismo para sobrevivir al tracto digestivo de sus dispersores (aves y murciélagos), pues llegan al suelo en plena estación de lluvias (mayo). La PD en al menos una fracción de las semillas de C. laurifolium parece ser de tipo intermedio, ya que la EMT solo fue efectiva en una fracción del lote (23 %) y tampoco aceleró la germinación; igualmente, el tratamiento pregerminativo con ácido giberélico no promovió la germinación (Sánchez, J.A., inédito) más de lo que lo hizo el de EMT empleado.
Las semillas de B. cassinifolia, P. sexflora, P. suberosa, M. glabra, P. domingensis y D. rugosa necesitaron más de 28 días para iniciar la germinación (entre 39,3 y 97,3 días), mostraron embriones desarrollados y resultaron permeables al agua, por lo que no hay dudas de que presentaron PD. Todas ellas, excepto D. rugosa, son dispersadas por los animales y llegan al suelo en la estación seca, lo que justifica la PD. En las semillas de B. cassinifolia el tratamiento pregerminativo fue efectivo para mejorar la germinación y por tal motivo estas presentaron PD no profunda, aunque llegaron al suelo al comienzo de la estación lluviosa. Algunas especies arbóreas de Psychotria (Rubiaceae) son descritas como dormantes (Garwood 1983; Baskin y Baskin, 1998; Sánchez et al., 2012). Un 38,3 % de las semillas de P. domingensis al parecer tuvieron PD no profunda, pero para el resto no hubo evidencias suficientes que permitieran precisar el nivel de PD. Esta situación fue similar para P. sexflora, P. suberosa y M. glabra.
Finalmente, las semillas de D. tamoidea, S. mollis y S. laurifolia mostraron embriones subdesarrollados y por consiguiente dormancia MD o MPD, tal como se ha informado para las familias Smilacaceae y Discoriaceae (Baskin y Baskin, 1998). Una fracción importante de las de D. tamoidea (67,8 %) comenzaron la germinación después de los 30 días (35,7 días); por consiguiente, dicha fracción debió presentar MD, aunque después de los 60 días de la siembra más del 20 % permanecieron vivas sin germinar, lo que sugiere que al menos ese porcentaje de semillas tuvo MPD. Las semillas de S. mollis y S. laurifolia presentaron MPD, porque la germinación comenzó después de los 40 días y más de un 70 % no germinaron. La otra liana estudiada (C. racemosa) también mostró dormancia, pero de clase fisiológica, probablemente no profunda, ya que alcanzó el 100 % de germinación en la oscuridad, y bajo estas condiciones la germinación comenzó a los 32 días (datos no mostrados). La dormancia en esta forma de vida parece ser bastante común, pues las dos especies de lianas no estudiadas (Forteronia corymbosa y Smilax havanensis ) pertenecen a familias con dormancia (Baskin y Baskin, 1998). También Gehan et al. (2012) encontraron PD y PY en lianas tropicales de la familia Fabaceae.
CONSIDERACIONES FINALES
Los resultados demostraron que los factores abióticos (luz y temperatura) afectaron considerablemente la germinación de las especies arbustivas. En las trepadoras, cuando se analizó la respuesta germinativa en su totalidad, ninguno de los factores ensayados influyó significativamente en esta variable, lo que sugiere que estas plantas pueden ocupar muchos micrositios; sin embargo, para algunas de ellas aparece una respuesta especie-específica. Por su parte, la respuesta al rango de temperatura de 25/40 ºC evidenció que muchas de las especies arbustivas pueden ser muy sensibles al incremento de temperatura del aire, como se propone en los posibles escenarios del cambio climático (IPCC, 2007), que en el caso de Cuba se estima que varíe entre 1,6 ºC y 2,5 ºC en 2100 (Capote, Mitrani y Suárez, 2011). Por tanto, el cambio climático podría alterar la composición y la estructura de la vegetación y, de este modo, inducir una sustitución progresiva de las clases de dormancia que componen las especies de la comunidad vegetal, o sea, se favorecerían aquellas especies con dormancia física. Sin embargo, de acuerdo con los resultados, este tipo de dormancia no es muy común en los arbustos que componen esta formación boscosa, por lo que el estrés hídrico y el calórico (sequía) podrían favorecer otro tipo de estrategia para asegurar el establecimiento de la comunidad por ejemplo, el incremento de especies no dormantes, o con dormancia fisiológica profunda, o bien beneficiar a aquellas especies con proporciones altas de recursos destinados a las estructuras de defensa testa/endocarpo (Garwood, 1983; Salazar et al., 2011).
AGRADECIMIENTOS
Esta investigación fue financiada por la Fundación Internacional para la Ciencia (IFS) mediante el donativo D/3536-2 dado a J. A. Sánchez. También los estudios se realizaron en el marco del proyecto «Ecofisiología de semillas y plántulas de árboles y arbustos de la Sierra del Rosario», del Programa Ramal de Diversidad Biológica de Cuba.
REFERENCIAS BIBLIOGRÁFICAS
1. Acevedo-Rodríguez, P. & Strong, M. T. Catalogue of seed plants of the West Indies. Washington, D.C.: Smithsonian Institution Scholarly Press, 2012.
2. Alm, D. M.; Stoller, E. W. & Wax, L. M. An index model for predicting seed germination and emergence rates. Weed Technol. 7:560-769, 1993.
3. Anderson, M. J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 26:32-46, 2001.
4. Baskin, Carol C. & Baskin, J. M. Seeds: ecology, biogeography and evolution of dormancy and germination. San Diego, USA: Academic Press, 1998.
5. Baskin, Carol C. & Baskin, J. M. Seed dormancy in trees of climax tropical vegetation types. Trop. Ecol. 46:17-28, 2005.
6. Baskin, J. M. & Baskin, Carol C. Classification, biogeography, and phylogenetic relationships of seed dormancy. In: R. D. Smith, J. B. Dickie, S. H. Linington, H. W. Pritchard and R. J. Probert, eds. Seed conservation turning science into practice. London: Kew Royal Botanical Gardens. p. 518-544, 2003.
7. Baskin, J. M. & Baskin, Carol C. A classification system for seed dormancy. Seed Sci. Res. 14:1-163, 2004.
8. Baskin, J. M.; Baskin, Carol C. & Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Spec. Biol. 15:139-152, 2000.
9. Brancalion, P. H. S.; Novembre, Ana D. L. C.; Rodrigues, R. R. & Marcos-Filho, J. Dormancy as exaptation to protect mimetic seeds against deterioration before dispersal. Ann. Bot. 105:991-998, 2010.
10. Bu, H.; Chen, X.; Xu, X.; Liu, K. & S. Wen, S. Community-wide germination strategies in an alpine meadow on the eastern Qinghai-Tibet plateau: phylogenetic and life-history correlates. Plant Ecol. 195:87-98, 2008.
11. Capote, R. P.; Mitrani, Ida & Suárez, A. Conservación de la biodiversidad cubana y cambio climático en el archipiélago cubano. Anales de la Academia de Ciencias de Cuba. 1:1-25, 2011.
12. Engelbrecht, B. M. J.; Dalling, J. W.; Pearson, T. R. H.; Wolf, R. L.; Galvez, D. A.; Koehler, T. et al. Short dry spells in the wet season increase mortality of tropical pioneer seedlings. Oecologia. 148:258-269, 2006.
13. Flores, J.; Jurado, E.; Chapa-Vargas, L.; Ceroni-Stuva, A.; Dávila-Aranda, P.; Galíndez; G. et al. Seeds photoblastism and its relationship with some plant traits in 136 cacti taxa. Environ. Exp. Bot. 71:79-88, 2011.
14. Garwood, Nancy. Seed germination in a seasonal tropical forest in Panama: a community study. Ecol. Monogr. 53:159-181, 1983.
15. Gehan, K. M. G.; Baskin, J. M.; Baskin, Carol C. & Fernando, M. T. Variation in seed dormancy and storage behavior of three liana species of Derris (Fabaceae, Faboideae) in Sri Lanka and ecological implications. Res. J. Seed Sci. 5 (1):1-18, 2012.
16. Grime, J. P.; Mason, G.; Curtis, A. A.; Rodman, J.; Band, S. R.; Mowforth, M. A. G. et al. A comparative study of germination characteristics in a local flora. J. Ecol. 69:1017-1059, 1981.
17. Herrera, R. A.; Menéndez, L.; Rodríguez, M. E. & García, E. E., Eds. et al. Ecología de los bosques siempreverdes de la Sierra del Rosario, Cuba. Proyecto MAB No. 1, 1974-1987. Montevideo: ROSTLAC, 1988.
18. IPCC. Cambio climático 2007: Informe de síntesis. Contribución de los Grupos de trabajo I, II y III al Cuarto Informe de evaluación del Grupo Intergubernamental de Expertos sobre el Cambio Climático. Ginebra: OMM, PNUMA, 2007.
19. ISTA. International Rules for Seed Testing. Seed Sci. & Technol. 27 (suppl):1-333, 1999.
20. Jurado, E. & Moles, Angela. Germination deferment strategies. In: G. Nicolás, K. J. Bradford, D. Côme, M. Curie and H. W. Pritchard, H. W., eds. The biology of seed: Recent research advances. Wallingford, UK: CAB International. p. 381-388, 2003.
21. Jurado, E. & Flores, J. Is seed dormancy under environmental control or bound to plant traits. J. Veg. Sci. 16: 559, 2005.
22. Kos, M.; Baskin, Carol C. & Baskin, J. M. Relationship of kinds of seed dormancy with habitat and life history in the Southern Kalahari flora. J. Veg. Sci. 23:869-879, 2012.
23. Marañón, T.; Camarero, J. J.; Castro, J.; Díaz, M.; Espelta, J. M.; Hampe, A. et al. Heterogeneidad ambiental y nicho de regeneración. En: F. Valladares, ed. Ecología del bosque mediterráneo en un mundo cambiante. Madrid: EGRAF. p. 69-99, 2008.
24. Milberg, P.; Andersson, L. & Thompson, K. Large-seeded species are less dependent on light for germination than small-seeded ones. Seed Sci. Res. 10:99-104, 2000.
25. Montejo, Laura et al. Caracterización seminal de un bosque tropical del oeste de Cuba. Correlaciones ecológicas entre rasgos. Bosque (En prensa), 2014.
26. Montejo, Laura; Muñoz, Bárbara; Sánchez, J. A. & Gamboa, A. Variabilidad seminal entre las especies de un bosque siempreverde tropical de Sierra del Rosario, Cuba. Bosque. 35:37-47, 2014b.
27. Muñoz, Bárbara et al. Guía técnica para la reproducción de especies arbóreas pioneras. La Habana: Ministerio de Ciencia, Tecnología y Medio Ambiente, Instituto de Ecología y Sistemática, 2012.
28. Norden, N.; Daws, M. I.; Antoine, C.; González, M. A.; Garwood, N. C. & Chave, J. The relationship between seed mass and mean time to germination for 1037 tree species across five tropical forests. Funct. Ecol. 23:203-210, 2009.
29. Pearson, T. R. H.; Burslem, D. F. R. P.; Mullins, C. E. & Dalling, J. W. Germination ecology of neotropical pioneers: interacting effects of environmental conditions and seed size. Ecology. 83:2798-2807, 2002.
30. Pons, T. L. Seed responses to light. In: M. Fenner, ed. Seeds: The ecology of regeneration in plant communities. Wallingford, UK: CAB International. p. 237-260, 2000.
31. Ricardo, Nancy. & Herrera, P. Las plantas expansivas nativas de Cuba. Apófitos. Acta Bot. Cub. 208:17-32, 2010.
32. Salazar, Ana; Goldstein, G. Franco, A. C. & Miralles-Wilhelm, F. Timing of seed dispersal and dormancy, rather that persistence in soil seed-banks, control recruitment of woody plants in Neotropical savanna. Seed Sci. Res. 21:103-116, 2011.
33. Sanches, M. C. & Válio, I. F. M. Seed and seedling survival of some climber species in a southeast brazilian tropical forest. Biotropica. 34:323-327, 2002.
34. Sánchez, J. A.; Montejo, Laura & Muñoz, Bárbara. Efectos de tratamientos robustecedores de semillas sobre la germinación y establecimiento de árboles pioneros bajo condiciones de estrés. Ecotrópicos. 16:91-112, 2003.
35. Sánchez, J. A.; Muñoz, Barbara; Montejo, Laura & Herrera, R. Ecological grouping of tropical trees in an evergreen forest of the Sierra del Rosario, Cuba. Acta Bot. Cub. 204:14-23, 2009.
36. Sánchez, J. A. et al. El cambio climático y las semillas de las plantas nativas cubanas. Acta Bot. Cub. 214:38, 2011.
37. Sánchez, J. A. et al. Ecofisiología de semillas y plántulas de árboles y arbustos de la Sierra del Rosario. Informe Final de Proyecto, Programa Diversidad Biológica (DB-032). La Habana: Instituto de Ecología y Sistemática, Agencia de Medio Ambiente, 2012.
38. Sautu, Adriana; Baskin, J. M.; Baskin, Carol C.; Deago, J. & Condit, R. Classification and ecological relationships of seed dormancy in a seasonal moist tropical forest, Panama, Central America. Seed Sci. Res. 17:127-140, 2007.
39. Silveira, F. A. O.; Ribeiro, Rafaella C.; Oliveira, Denise M. T.; Wilson Fernandes, G. & Lemos-Filho, J. P. Evolution of physiology dormancy multiple times in Melastomataceae from Neotropical montane vegetation. Seed Sci. Res. 22:37-44, 2012.
40. The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 141:399-436, 2003.
41. Thompson, K.; Ceriani, Roberta; Bakker, J. P. & Bekker, Renée M. Are seed dormancy and persistence in soil related? Seed Sci. Res. 13:97-100, 2003.
42. Vázquez-Yanes, C. & Orozco-Segovia, Alma. Signals for seeds to sense and respond to gaps. In: M.M. Caldwell and R.W. Pearcy, eds. Exploitation of environmental heterogeneity by plants. San Diego, USA: Academic Press. p. 209-236, 1994.
43. Vázquez-Yanes, C. & Orozco-Segovia, Alma. Seed dormancy in the tropical rain forest. In: K. S. Bawa and M. Hadley, eds. Reproductive ecology of tropical forest plants. New Jersey: Parthenon. p. 247-260, 1990.
Recibido el 16 de diciembre de 2013
Aceptado el 2 de diciembre de 2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}