










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkPastos y Forrajes
On-line version ISSN 2078-8452
Pastos y Forrajes vol.38 no.1 Matanzas Jan.-Mar. 2015
ARTÍCULO DE INVESTIGACIÓN
Perfil cualitativo de metabolitos secundarios en la fracción comestible de especies leñosas seleccionadas por vacunos en un bosque semicaducifolio
Qualitative profile of secondary metabolites in the edible fraction of woody species selected by cattle in a semi-deciduous forest
Á. Ojeda1, N. Obispo2, J. L. Gil2 e Irana Matute2
1Instituto de Producción Animal, Facultad de Agronomía, Universidad Central de Venezuela. Maracay 2101, Venezuela. Correo electrónico: ajojeda99@yahoo.com
2Instituto Nacional de Investigaciones Agrícolas. Centro Nacional de Investigaciones Agropecuarias. Maracay 2101, Aragua. Venezuela.
RESUMEN
Con el fin de caracterizar cualitativamente el perfil de metabolitos secundarios de la fracción comestible de plantas leñosas seleccionadas por vacunos en silvopastoreo, las cuales se encuentran en un bosque tropófilo semicaducifolio de Venezuela, se utilizaron 14 vacunos machos Brahman x Holstein de 390,4 ± 18,0 kg de PV. Estos fueron manejados en pastoreo continuo en una superficie de 81 ha, de las cuales 50 ha correspondían a un bosque semicaducifolio y el resto, a vegetación herbácea natural (Cynodon nlemfuensis, Sporobolus indicus, Axonopus sp., Mimosa pudica e Hyptis suaveolens). La composición de la biomasa leñosa seleccionada por los animales se estudió mediante la evaluación de los fragmentos epidérmicos presentes en las heces colectadas por vía transrectal, cada 15 días. A partir de estos resultados, se tomaron muestras de cinco plantas por especie leñosa, en los periodos seco y lluvioso. Se determinaron 15 grupos de metabolitos: alcaloides, á-aminos, carbohidratos solubles, cardenólidos, cianógenos, esteroles, fenoles totales, flavonoides, lectinas, mucílagos, quinonas, saponinas, taninos extractables (totales y condensados) y terpenoides. La mayoría de la biomasa correspondió a especies de la familia Fabaceae, las cuales presentaron todos los metabolitos evaluados. En ambos periodos se detectaron niveles de esteroles, terpenoides, fenoles totales y taninos extractables que variaron desde leves hasta cuantiosos. Con independencia del periodo, el 66,7 % de las especies presentó entre 10-11 metabolitos secundarios en su biomasa comestible. Se recomienda realizar estudios para la cuantificación de los metabolitos secundarios presentes en plantas leñosas de valor forrajero, para este sistema silvopastoril.
Palabras clave: Fabaceae, sistemas silvopascícolas.
ABSTRACT
In order to characterize qualitatively the profile of secondary metabolites of the edible fraction of woody plants selected by cattle in a silvopastoral system of a tropophilous semi-deciduous forest of Venezuela, 14 Brahman x Holstein bulls of 390,4 ± 18,0 kg of LW were used. They were managed under continuous grazing on a surface of 81 ha, from which 50 ha corresponded to a semi-deciduous forest and the rest, to natural herbaceous vegetation (Cynodon nlemfuensis, Sporolobus indicus, Axonopus sp., Mimosa pudica and Hyptis suaveolens). The composition of the woody biomass selected by the animals was studied through the evaluation of the epidermal fragments present in the feces collected from the rectum, every 15 days. From these results, samples were taken from five plants per woody species, in the dry and rainy seasons. Fifteen groups of metabolites were determined: alkaloids, á-amines, soluble carbohydrates, cardenolides, cyanogens, sterols, total phenols, flavonoids, lectins, mucilages, quinones, saponins, extractable (total and condensed) tannins and terpenoids. Most of the biomass corresponded to species from the Fabaceae family, which showed all the evaluated metabolites. In both seasons levels of sterols, terpenoids, total phenols and extractable tannins which varied from slight to abundant, were detected. Independently from the season, 66,7 % of the species showed between 10 and 11 secondary metabolites in their edible biomass. It is recommended to conduct studies for the quantification of the secondary metabolites present in woody plants of forage value, for this silvopastoral system.
Key words: Fabaceae, silvopastoral systems.
INTRODUCCIÓN
Se estima que en América existen 519.597 km2 de sabanas, de las cuales el 51 % se ubica en el sur del continente. De esta superficie, Venezuela cuenta con 29.396 km2, asociados en un 80 % a formaciones de bosque seco tropical, donde los productores promueven el ingreso de los vacunos a las áreas boscosas a medida que avanza el periodo seco, en una modalidad desarrollada de modo empírico, con el objetivo de garantizar el incremento de la cantidad y calidad de la oferta forrajera (Baldizán y Chacón, 2007; Portillo y Sánchez, 2010).
Para diseñar estrategias de manejo sostenible del silvopastoreo en bosques tropicales es necesario profundizar en los estudios de caracterización de los recursos vegetales, con particular énfasis en su valoración química y nutricional. Aunque a nivel local se han realizado estudios sobre algunas plantas leñosas de valor forrajero, así como evaluaciones de sistemas silvopastoriles en formaciones de bosque seco caducifolio (Benezra et al., 2003; Baldizán et al., 2006; García et al., 2006), no existe información disponible acerca de los perfiles de los metabolitos secundarios presentes en la biomasa de las leñosas que son seleccionadas por los vacunos al pastar en los bosques secos semicaducifolios.
El término «metabolitos secundarios» se emplea para identificar sustancias ecológicamente eficaces, de amplia diversidad química y que pueden ejercer efectos contrarios en la nutrición óptima de los animales, debido a la afectación de los procesos digestivos y metabólicos (Kumar, 1992). Los metabolitos secundarios superan los 80.000 compuestos, entre los que se destacan: 29.000 terpenoides, 12.000 alcaloides, 8.000 polifenoles, 600 aminoácidos no proteínicos, 100 aminas y 100 cianógenos. Estas sustancias tienen importancia funcional para los tejidos vegetales, ya que actúan como agentes alelopáticos y contra las invasiones de hongos; ofrecen protección contra la radiación ultravioleta y la deshidratación, con reserva de nitrógeno; y establecen relaciones de mutualismo y toxicidad con los depredadores, entre otros beneficios (Mazid et al., 2011).
La ingestión de biomasa vegetal que contenga estos metabolitos puede generar efectos adversos en el consumo voluntario, la digestión y la absorción de nutrientes, el metabolismo intermediario y el funcionamiento de órganos o sistemas (Sorensen et al., 2005); aunque se ha señalado que las concentraciones moderadas son beneficiosas para la nutrición de los rumiantes, ya que ocurre una mejora en el patrón de síntesis de ácidos grasos volátiles y un incremento del flujo de nutrientes al tracto posterior, así como una reducción del meteorismo, los parásitos gastrointestinales y las emisiones entéricas de metano (Rochfort et al., 2008).
El objetivo de este estudio fue caracterizar cualitativamente el perfil de metabolitos secundarios de la fracción comestible de plantas leñosas seleccionadas por vacunos en silvopastoreo, en un bosque tropófilo semicaducifolio de Venezuela.
MATERIALES Y MÉTODOS
La investigación se realizó en la Estación Experimental San Nicolás (8º 49´ 58´´ de latitud Norte y 69º 48´ 00´´ de longitud Oeste), adscrita a la Universidad Central de Venezuela. Se seleccionó una superficie de 81 ha, con suelos clasificados como Fluventic Haplustepts y Vertic Endoaquepts, de textura arcillosa fina mixta, no ácida e isohipertérmica (Abarca, 2005).
Durante el periodo experimental la unidad climatológica de la estación registró 1.253,8 mm de precipitación; 5,9 ± 1,01 horas de insolación; 27,7 ± 0,62 ºC de temperatura; 1.832,5 mm de evapotranspiración potencial y 72,2 ± 14,3 % de humedad relativa. La relación precipitación-temperatura permitió definir un periodo lluvioso (mayo-noviembre) y uno seco (diciembre-abril).
Se utilizaron 14 vacunos, machos enteros mestizos Brahman x Holstein, de 390,4 ± 18,0 kg de PV, en pastoreo continuo. De la superficie considerada, 50 ha correspondieron a un bosque tropófilo semicaducifolio, de bajo nivel de intervención antrópica; mientras el resto estaba ocupado por vegetación herbácea (Cynodon nlemfuensis, Sporobolus indicus, Axonopus sp., Mimosa pudica e Hyptis suaveolens, entre otras), con un rendimiento de 969,3 ± 28,6 kg MS/ha, y una relación hoja:tallo de 0,53 ± 0,15. Esta información se obtuvo a partir de cuadrículas de 0,25 m2, localizadas cada 10 m sobre transectas en forma de zig-zag, de aproximadamente 120 m de longitud, las cuales se ubicaron en función de la variación vegetal y la topografía del área de muestreo.
A los animales se les suministró agua ad libitum y una mezcla comercial de sales y minerales, compuesta por: 20 % de Ca; 10 % de P; 9 % de Na; 0,6 % de S; 0,25 % de Cu y 80 ppm de I; entre otros.
Para permitir la selectividad de los animales en condiciones de la carga animal usual para las sabanas naturales de la zona, y en función de la biomasa graminiforme, se emplearon vacunos adicionales, con el objetivo de garantizar una presión de pastoreo de 6 kg de MS/100 kg de PV, y una variación de peso de 450 ± 35 g/animal/día. Quince días previos al inicio de la evaluación, estos fueron desparasitados con clorhidrato de levamisol (1 mL/50 kg de peso) y se les aplicó 1 mL/100 kg de PV de un complejo multivitamínico (500.000 UI/mL vit. A, 75.000 UI/mL vit. D3 y 50 UI/mL vit. E).
La selectividad de la biomasa leñosa se registró durante el periodo seco, ya que solo en este lapso de tiempo los productores permiten el silvopastoreo de los bosques, para después promover la recuperación del ecosistema durante el periodo lluvioso. La composición estructural de la biomasa leñosa se estimó mediante la evaluación de muestras de heces, colectadas de forma individual y directamente del recto, entre las 8:00 y 10:00 h, cada 15 días.
Las muestras se depositaron en bolsas plásticas y se almacenaron a -18 ºC, hasta su deshidratación en estufa con ventilación forzada a 65 ºC, durante 48 h. A continuación, se molieron en criba de 1 mm Ø. De este material se tomaron 2 g, que se sometieron a procesos de rehidratación con alcohol etílico al 50 % v/v, tamizado (180 µ) y remoción de los pigmentos vegetales con hipoclorito de sodio; el montaje se realizó en láminas semipermanentes, a razón de 3 láminas de 24 x 24 mm por cada muestra (Holechek, 1982).
Los fragmentos epidérmicos de la biomasa leñosa se observaron a través de un microscopio binocular, a 400 X, y se compararon con patrones fotográficos digitales obtenidos del tejido epidérmico foliar de las plantas leñosas presentes en el bosque, las cuales se identificaron botánicamente en el Herbario Víctor Manuel Badillo (MY), de la Universidad Central de Venezuela.
El diseño fue completamente aleatorizado y se tomaron muestras de cinco plantas por cada especie leñosa seleccionada por los animales. Las muestras se agruparon por planta, de acuerdo con dos periodos del año (seco y lluvioso), y se colectaron mensualmente con el empleo de tijeras de poda y a razón de 200 g/planta. A los fines de la toma de muestras, se consideraron como susceptibles de ramoneo por los vacunos adultos las estructuras vegetales con diámetro inferior a 6 mm y altura máxima de 2 m.
El perfil cualitativo de los metabolitos secundarios se determinó de acuerdo con la metodología propuesta por Rondina y Coussio (1969), con las modificaciones sugeridas por Galindo et al. (1989) y García (2003). Para ello, se tomaron 25 g de cada muestra y se maceraron con 25 mL de etanol al 98 %, durante 4 h, en un equipo Soxhlet. El extracto obtenido se filtró en papel de 4 µm de diámetro hasta un volumen de 50 mL. A partir de esta solución, y por el procesamiento sucesivo de filtrados y residuos sólidos, se obtuvo un total de seis fracciones. Después se realizaron las determinaciones cualitativas de 15 grupos de metabolitos: alcaloides, á-aminos, carbohidratos reductores solubles, cardenólidos, cianógenos, esteroles, fenoles totales, flavonoides, lectinas, mucílagos, quinonas, saponinas, taninos extractables (totales y condensados) y terpenoides. La presencia o ausencia de estos compuestos se sistematizó a partir de: la presencia cuantiosa (+++), moderada (++) o leve (+), y la ausencia (-). En el caso de las saponinas, se empleó la altura de la espuma como criterio, y se consideraron los siguientes niveles: abundantes (> 14 mm), moderados (10-14 mm) o bajos (< 10 mm), y ausentes (0 mm).
RESULTADOS Y DISCUSIÓN
En las tablas 1 y 2 se muestran los perfiles cualitativos de los metabolitos secundarios, en las dos épocas del año, correspondientes a la fracción comestible de las plantas leñosas seleccionadas por los vacunos en silvopastoreo.
Las plantas ramoneadas pertenecían a las familias botánicas siguientes: Boraginaceae (Rochefortia spinosa), Fabaceae (Enterolobium cyclocarpum, Inga laurina, Machaerium humboldtianum, Pithecellobium lanceolatum, Samanea saman y Senna sp.), Polygonaceae (Coccoloba caracasana) y Sterculiaceae (Guazuma ulmifolia).
En un inventario florístico realizado en el bosque semicaducifolio, Ojeda et al. (2012) informaron que en una densidad de 463 plantas/ha existían cuatro especies (G. ulmifolia, P. lanceolatum, S. saman y Guadua angustifolia), que representaban el 80,2 % de la población de las plantas leñosas; de estas, las tres primeras acumularon los mayores registros para el Índice de Valor de Importancia (IVI). El IVI es una variable que se relaciona con el consumo voluntario, ya que, en ausencia de limitantes físicas o químicas, la distribución espacial influye en la oportunidad que presentan los animales en pastoreo de tener contacto con una especie en particular (Provenza y Balph, 1988), lo cual permite explicar que el 66,7 % de las especies seleccionadas en esta evaluación fueran fabáceas.
La preferencia de los rumiantes en silvopastoreo por esta familia ha sido documentada por otros autores (Benezra et al., 2003; Mayer et al., 2003; Rutter, 2006). Si bien los mayores componentes de la ración tienden a mantener su prevalencia en diferentes circunstancias de manejo, se debe señalar que las variaciones en la composición botánica de la dieta pueden ser influenciadas por las condiciones climáticas que regulan la variabilidad temporal y espacial de la oferta forrajera; así como por algunos factores intrínsecos de la planta, entre los que se destacan el valor nutricional, la arquitectura, la edad y el estado fisiológico del animal, la intensidad de pastoreo, la experiencia alimentaria, el aprendizaje social (individual y colectivo) y la respuesta neofóbica (Marquardt et al., 2009).
Por otra parte, los 15 metabolitos secundarios evaluados se encontraron en la fracción comestible de las plantas leñosas. Asimismo, con independencia del periodo, las plantas presentaron esteroles, terpenoides, fenoles totales y taninos extractables (totales y condensados).
Los esteroles y los terpenoides tuvieron concentraciones de leves a moderadas, con una mayor presencia en E. cyclocarpum y P. lanceolatum, en el periodo seco; mientras que en el lluvioso predominaron en I. laurina, P. lanceolatum y S. saman.
Asimismo, se identificaron mezclas de esteroides (tonos azules) y triterpenoides poliglicosilados (coloraciones verdes y amarillas) que indican la presencia de ß-citosterol y estigmasterol, metabolitos que se relacionan con los procesos de activación del crecimiento vegetal y la biosíntesis de saponinas esteroidales (C27) y triterpénicas (C30) de potencial efecto tóxico (Marcano y Hasegawa, 2002). Como grupo, los esteroles son abundantes en Morus alba (García, 2003); mientras que en Venezuela se reportan en concentraciones variables en Acacia flexuosa, Acacia glomerosa, Acacia macracantha, Albizia lebbeck, Albizia caribaea, Calliandra crugieri, Cassia grandis, Cassia fistula, Pithecellobium dulce, S. saman, Gliricidia sepium, Leucaena macrophylla, Lysiloma latisiliquum y Enterolobium contortisiliquum (Baldizán et al., 2006).
En cuanto a los terpenoides, Marcano y Hasegawa (2002) señalan que estos metabolitos presentan actividad biocida, sedante y antihelmíntica (monoterpenos); efectos alelopáticos, alérgenos y tóxicos a microorganismos del rumen (sesquiterpenos); hipersensibilidad, disnea, hipotensión, irritaciones cutáneas y de las mucosas, neuropatía y alopecia (diterpenos); así como defaunantes y alteradores de la presión sanguínea (triterpenos).
Las saponinas, aunque no se encontraron en P. lanceolatum, G. ulmifolia y C. caracasana, tuvieron niveles de leves a moderados en el resto de las plantas, con independencia del periodo. De naturaleza esteroidal o triterpénica, se vinculan con alteraciones de las membranas celulares, por lo que se asocian a la destrucción de protozoarios ruminales y facilitan el flujo de proteína microbial al tracto posterior. En elevadas cantidades, las saponinas se relacionan con el meteorismo por ser potentes espumantes; así como por reducir el colesterol sanguíneo y hepático y la tasa de crecimiento, inhibir la actividad del músculo liso y algunas enzimas, y regular la absorción de nutrientes (Francis et al., 2002).
Los fenoles totales tuvieron presencia de leve a cuantiosa, que fue mayor durante el periodo lluvioso, particularmente en I. laurina y S. saman. Estos compuestos aromáticos no siempre constituyen factores antinutricionales, al formar parte de todas las plantas vasculares de importancia en la producción animal, ya sea en forma libre (semillas, frutos y tejidos muertos) o polimerizados, como es el caso de los taninos y la lignina. En este sentido, se ha informado la presencia de fenoles totales en géneros comunes al bosque en estudio, tales como: Acacia, Albizia, Cadaba, Caesalpinia, Calliandra, Cassia, Copernicia, Enterolobium, Gliricidia, Inga, Lablab, Leucaena, Lysiloma, Macroptilium, Moringa, Morus, Pithecellobium y Trichantera, entre otros (Romero et al., 2000; García, 2003; Baldizán et al., 2006; García y Medina, 2006).
En el caso de los taninos, estos mostraron un patrón de distribución similar al de los fenoles totales, con un aumento durante el periodo lluvioso para la fracción de taninos condensados en I. laurina, P. lanceolatum, S. saman y G. ulmifolia. Como mecanismo de defensa ante los depredadores, en algunas plantas vasculares es evidente su incremento durante los periodos de crecimiento activo. Aunque con frecuencia los taninos son identificados como factores antinutricionales, por su potencial efecto negativo en el consumo voluntario, la digestión de nutrientes y la respuesta animal, se consideran de bajo impacto nutricional cuando su presencia es de 2-4 % de biomasa seca ingerida. Incluso, se les asignan propiedades beneficiosas para el rumiante en pastoreo, ya que reducen la degradación ruminal de la proteína ingerida e incrementan su flujo al tracto posterior. Además, junto con las saponinas, actúan contra los parásitos gastrointestinales y reducen las emisiones entéricas de metano (Ramírez-Restrepo y Barry, 2005; Goel y Makkar, 2012).
Por otra parte, los alcaloides están altamente asociados al genotipo y son poco dependientes de factores ecológicos tales como el clima, las estaciones o el periodo del año y la disponibilidad de agua. Así, en este estudio estuvieron ausentes en P. lanceolatum y G. ulmifolia, y con presencia leve en el resto de las plantas evaluadas. Con frecuencia estos metabolitos son considerados como parte del mecanismo de defensa de las plantas, por su carácter neurotóxico; además, actúan como reservas de nitrógeno y en la fijación de ciertos metales, debido a su capacidad quelatante (Marcano y Hasegawa, 2002). Los niveles encontrados en las plantas leñosas seleccionadas sugieren que la capacidad de detoxificación ruminal es capaz de evitar los efectos adversos al macroanimal, y que pueden ser empleados como fuentes de nitrógeno y minerales para el crecimiento microbial.
Independientemente del periodo, los carbohidratos reductores solubles estuvieron ausentes en el 55,6 % de las plantas leñosas seleccionadas, con presencia moderada en I. laurina. En los esquemas de silvopastoreo en ambientes de amplia biodiversidad, como los del presente estudio, no se espera que estos intermediarios del metabolismo energético causen problemas al animal, por lo que deben constituir un aporte de energía fácilmente fermentable al medio ruminal.
Los restantes metabolitos mostraron presencia leve o estuvieron ausentes independientemente del periodo evaluado, lo que indica el poco efecto que ejercieron sobre los rumiantes en silvopastoreo en esta formación boscosa, como consecuencia de la variedad en la biomasa seleccionada. En este sentido, resaltan los niveles moderados de los grupos á-aminos y mucílagos, durante el periodo seco, en E. cyclocarpum y G. ulmifolia, respectivamente; así como en R. spinosa y E. cyclocarpum (á-aminos) y G. ulmifolia (flavonoides y mucílagos), durante el lluvioso.
Algunos á-aminos son considerados metabolitos secundarios, por su capacidad de generar proteínas anómalas; mientras que los mucílagos, restringidos en este estudio a su presencia exclusiva en G. ulmifolia, se identifican como fibra soluble de naturaleza viscosa, con características análogas a las de las gomas. En la literatura no se informa (Marcano y Hasegawa, 2002) que estos compuestos constituyan factores antinutricionales, con excepción de los que presentan propiedades cáusticas e irritantes, los cuales no fueron identificados en las plantas de este estudio.
Los flavonoides (flavonas, flavonoles, antocianidinas, leucoantocianidinas, catequinas, charconas, isoflavonas y quinonas; entre otros) solo se encontraron en tres especies (S. saman, Senna sp. y G. ulmifolia), y constituyen un grupo que depende del componente genético. Además, son altamente polares y se asocian a la función de secuestradores de radicales libres (antioxidantes), antisenescentes, diuréticos y antimicrobiales (García, 2003).
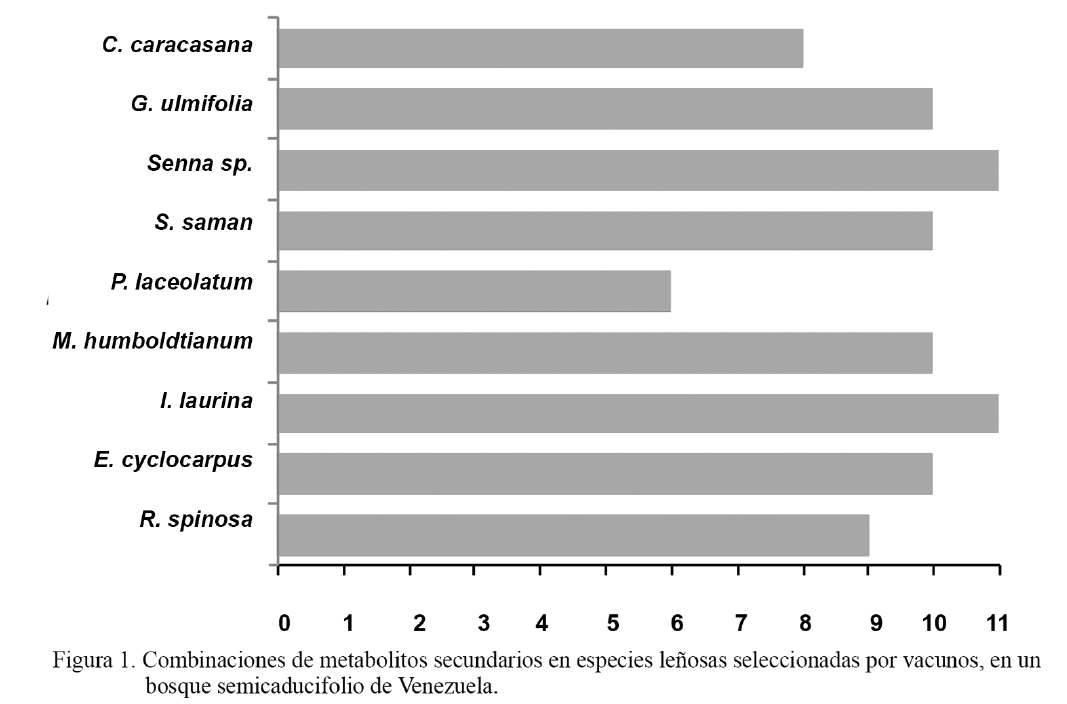
Según las diferentes combinaciones de metabolitos secundarios identificados en la fracción comestible de las plantas leñosas seleccionadas por los vacunos, la menor combinación correspondió a P. lanceolatum, con 6 metabolitos diferentes; y la mayor, a I. laurina y Senna sp., con 11 metabolitos en cada especie (fig. 1). En términos relativos, e independientemente del periodo del año, el 66,7 % de las especies presentó entre 10-11 metabolitos secundarios en su biomasa comestible.
Al comparar las fabáceas con el resto de las familias botánicas, estas fueron superiores en cuanto al número de metabolitos secundarios, los que estuvieron de modo concurrente en una misma especie.
En general, las especies fijadoras de nitrógeno sintetizan una mayor diversidad de estructuras secundarias, como estrategia defensiva en un medio que les resulta particularmente agresivo, ya que los defoliadores las seleccionan por su mayor valor biológico, lo que se asocia frecuentemente a la oferta de fracciones nitrogenadas. Esta relación, derivada de los procesos coevolutivos entre plantas y animales, constituye un aspecto fundamental para comprender las diferencias quimiotaxonómicas entre las especies leñosas y su impacto potencial en el plano nutricional del rumiante en silvopastoreo (Launchbaugh et al., 2001).
Finalmente, se debe señalar que algunos autores demuestran la existencia de interacciones entre los metabolitos secundarios, denominadas tradicionalmente interacciones antinutriente-antinutriente. Así, en el caso de la asociación taninos-saponinas, se indica un efecto aditivo en la disminución de la digestibilidad verdadera y la tasa de producción de gas in vitro, cuando ambos metabolitos están presentes en la ración (Makkar et al., 1995). De igual modo, se indica que la presencia de lectinas en plantas con taninos permite eliminar los efectos adversos de estos compuestos fenólicos sobre la amilasa (Fish y Thompson, 1991), o que los taninos reducen el efecto deletéreo de glucósidos cianogénicos, al interferir en la liberación de ácido cianhídrico (Goldstein y Spencer, 1985).
CONCLUSIONES
La evaluación del perfil cualitativo de metabolitos secundarios en la fracción comestible de especies leñosas seleccionadas por vacunos en silvopastoreo, en un bosque tropófilo semicaducifolio de Venezuela, permitió identificar una amplia variedad de compuestos presentes en la biomasa seleccionada, algunos de potencial efecto tóxico para los rumiantes. Esto debe ser considerado en futuras evaluaciones, para definir su valor forrajero en el marco de estrategias sustentables de silvopastoreo, en condiciones tropicales.
AGRADECIMIENTOS
Los autores agradecen al Consejo de Desarrollo Científico y Humanístico de la Universidad Central de Venezuela, por el apoyo financiero para la realización de esta investigación (Proyecto CDCH N° PG-01-36-4995-2002).
REFERENCIAS BIBLIOGRÁFICAS
1. Abarca, O. I. Conflictos de intensidad de uso de la tierra en las estaciones experimentales de la Universidad Central de Venezuela. Análisis espacial con sistemas de información geográfica. Agronomía Trop. 55 (2):289-313, 2005.
2. Baldizán, A. & Chacón, E. Utilización del recurso bosque de los llanos centrales con rumiantes. En: Memorias. I Simposio Tecnologías Apropiadas para la Ganadería de los Llanos de Venezuela. Valle de la Pascua, Venezuela: Universidad Nacional Experimental Rómulo Gallegos, 2007.
3. Baldizán, A.; Domínguez, C.; García, D. E.; Chacón, E. & Aguilar, L. Metabolitos secundarios y patrón de selección de dietas en el bosque deciduo tropical de los llanos centrales venezolanos. Zootecnia Trop. 24 (3):213-232, 2006.
4. Benezra, M., Cecconello, G. & Camacho, Fanny. Selección de especies leñosas en un bosque seco tropical por vacunos adultos usando análisis histológico fecal. Zootecnia Trop. 21 (1):73-85, 2003.
5. Fish, B. C. & Thompson, L. U. Lectin tannin interactions and their influence on pancreatic amylase activity and starch digestibility. J. Agric. Food Chem. 39 (4):727-731, 1991.
6. Francis, G.; Kerem, Z.; Makkar, H. P. & Becker, K. The biological action of saponins in animal systems: a review. Braz. J. Nutr. 88 (6):587-605, 2002.
7. Galindo, W.; Rosales, M.; Murgueitio, E. & Larrahondo, J. E. Sustancias antinutricionales en las hojas de guamo, nacedero y matarratón. Livest. Res. Rural Dev. 1 (1), 1989. http://www.lrrd.org/lrrd1/1/mauricio.htm [07/03/2014].
8. García, D. E. Evaluación de los principales factores que influyen en la composición fitoquímica de Morus alba (Linn.). Trabajo de grado. Matanzas, Cuba: EEPF Indio Hatuey, Universidad de Matanzas, 2003.
9. García, D. E. & Medina, María G. Composición química, metabolitos secundarios, valor nutritivo y aceptabilidad relativa de diez árboles forrajeros. Zootecnia Trop. 24 (3):233-250, 2006.
10. García, D. E.; Medina, María G.; Humbría, J.; Domínguez, C.; Baldizán, A.; Cova, L. et al. Composición proximal, niveles de metabolitos secundarios y valor nutritivo del follaje de algunos árboles forrajeros tropicales. Arch. Zootec. 55 (212):373-384, 2006.
11. Goel, G. & Makkar, H. P. S. Methane mitigation from ruminants using tannins and saponins. Trop. Anim. Health Pro. 44 (4):729-739, 2012.
12. Goldstein, W. S. & Spencer, K. C. Inhibition of cyanogensis by tannins. J. Chem. Ecol. 11 (7):847-857, 1985.
13. Holechek, J. L. Sample preparation technique for microhistological analysis. J. Range Manage. 35 (2):267-268, 1982.
14. Kumar, R. Anti-nutritional factors, the potential risks of toxicity and methods to alleviate them. In: A. Speedy and P. L. Plugliese, eds. Legume trees and other fodder trees as protein source for livestock. Rome: FAO. p. 145-160. FAO Animal Production and Health Paper No. 102, 1992.
15. Launchbaugh, K. L.; Provenza, F. D. & Pfister, J. A. Herbivore response to anti-quality factors in forages. J. Range Manage. 54 (4):431-440, 2001.
16. Makkar, H. P. S.; Blümmel, M. & Becker, K. In vitro effects of and interactions between tannins and saponins and fate of tannins in the rumen. J. Sci. Food Agric. 69 (4):481-493, 1995.
17. Marcano, D. & Hasegawa, M. Fitoquímica orgánica. 2a. ed. Caracas: Consejo de Desarrollo Científico y Humanístico. Universidad Central de Venezuela, 2002.
18. Marquardt, S.; Márquez, A.; Bouillot, Hélène; Beck, S. G.; Mayer, Andrea; Kreuzer, M. et al. Intensity of browsing on trees and shrubs under experimental variation of cattle stocking densities in southern Bolivia. Forest Ecol. Manag. 258 (7):1422-1428, 2009.
19. Mayer, Andrea C.; Stöckli, Veronika; Huovinen, Christine; Konold, W.; Estermann, Beda L. & Kreuzer, M. Herbage selection by cattle on sub-alpine wood pastures. Forest Ecol. Manag. 181 (1-2):39-50, 2003.
20. Mazid, M.; Khan, T. A. & Mohammad, F. Role of secondary metabolites in defense mechanisms of plants. Biol. Medic. 3 (2):232-249, 2011.
21. Ojeda, A.; Obispo, N.; Canelones, C. E. & Muñoz, D. Selección de especies leñosas por vacunos en silvopastoreo de un bosque semicaducifolio en Venezuela. Arch. Zootec. 61 (235):355-365, 2012.
22. Portillo, C. A. & Sánchez, G. A. Extent and conservation of tropical dry forests in the Americas. Biol. Conserv. 143 (1):144-155, 2010.
23. Provenza, F. D. & Balph, D. F. Development of dietary choice in livestock on rangelands and its implications for management. J. Anim Sci. 66 (9):2356-2368, 1988.
24. Ramírez-Restrepo, C. A. & Barry, T. N. Alternative temperate forages containing secondary compounds for improving sustainable productivity in grazing ruminants. Anim. Feed Sci. Tech. 120 (3-4):179-201, 2005.
25. Rochfort, S.; Parker, A. J. & Dunshea, F. R. Plant bioactives for ruminant health and productivity. Phytochemistry. 69 (2):299-322, 2008.
26. Rondina, R. & Coussio, J. Estudio fitoquímico de plantas medicinales argentinas. Rev. Invest. Agropec. (Serie 2). 6 (2):352-366, 1969.
27. Romero, Claudia; Palma, J. M. & López, J. Influencia del pastoreo en la concentración de fenoles totales y taninos condensados en Gliricidia sepium en el trópico seco. Livest. Res. Rural Dev. 12 (4):4, 2000. http://www.fao.org/ag/aga/agap/frg/lrrd/lrrd12/4/rome124.htm. [12/03/2014].
28. Rutter, S. M. Diet preference for grass and legumes in free-ranging domestic sheep and cattle: Current theory and future application. Appl Anim. Behav. Sci. 97 (1):17-35, 2006.
29. Sorensen, J. S.; McLister, J. D. & Dearing, Denise. Plant secondary metabolites compromise the energy budgets of specialist and generalist mammalian herbivores. Ecology. 86 (1):125-139, 2005.
Recibido el 5 de junio de 2014
Aceptado el 17 de noviembre de 2014


{kind=link}
{kind=link}
{kind=link}