










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkPastos y Forrajes
On-line version ISSN 2078-8452
Pastos y Forrajes vol.38 no.4 Matanzas Oct.-Dec. 2015
ARTÍCULO DE INVESTIGACIÓN
Caracterización faunística en un sistema silvopastoril destinado al ganado de engorde
Faunistic characterization in a silvopastoral system intended for beef cattle
J. Iraola1, E. Muñoz1, Yenny García1 y J. L. Hernández1
1Instituto de Ciencia Animal, apdo. postal 24, San José de las Lajas, Mayabeque, Cuba
Correo electrónico: jiraola@ica.co.cu
RESUMEN
La investigación se desarrolló durante cinco años, con el objetivo de evaluar el comportamiento faunístico en un sistema silvopastoril destinado al ganado de engorde. Se aplicó un modelo lineal mixto donde se usaron como efectos fijos el año, la época y la interacción año por época, y como efecto aleatorio los cuadrantes. Se determinaron los índices ecológicos número de individuos, riqueza y diversidad. También se determinó la sobrevivencia de árboles y la producción de biomasa, y se ponderó un índice de capacidad de carga biológica. Se realizaron análisis de regresiones simples entre la biomasa, los árboles y los índices número de individuos y diversidad. Se observó diferencia (p < 0,001) en el número de individuos totales entre los dos años iniciales, mientras que el segundo no difirió del tercero y el cuarto año difirió del quinto. El indicador riqueza difirió (p < 0,001) entre los diferentes años. Asimismo, hubo diferencias (p < 0,001) en la diversidad biológica y se hallaron relaciones positivas entre el componente arbóreo, la biomasa y los índices número de individuos totales y diversidad de especies. Se concluye que el sistema silvopastoril, como modelo de cobertura, posibilitó un hábitat más amplio para el desarrollo faunístico, y mejoró la relación de los índices ecológicos debido a la presencia de los árboles y el incremento de la biomasa en el sistema.
Palabras clave: Fauna, hábitat, sistemas silvopascícolas.
ABSTRACT
The research was developed during five years, in order to evaluate the faunistic performance in a silvopastoral system intended for beef cattle. A mixed linear model was applied where the year, season and the year by season interaction were used as fixed effects, and the quadrants were used as random effect. The ecological indexes number of individuals, richness and diversity were determined. The tree survival and biomass production were also determined, and a biological carrying capacity index was analyzed. Simple regression analyses were made among the biomass, the trees, and the indexes number of individuals and diversity. Difference (p < 0,001) was observed in the number of total individuals between the two initial years, while the second did not differ from the third and the fourth year differed from the fifth. The richness indicator differed (p< 0,001) among the different years. Likewise, there were differences (p< 0,001) in the biological diversity and positive relations were found among the tree component, the biomass and the indexes total individual number and species diversity. It is concluded that the silvopastoral system, as cover model, allowed a wider habitat for faunistic development, and improved the relation of the ecological indexes due to the presence of the trees and the increase of biomass in the system.
Key words: Fauna, habitat, silvopastoral systems.
INTRODUCCIÓN
La sistemática sucesión de acciones en el manejo que depende de insumos externos y prácticas adversas por parte de los productores ganaderos respecto a los recursos naturales, el suelo, la diversidad, la vegetación y los nacimientos y flujos naturales de agua, no favorece el funcionamiento equilibrado y natural de los agroecosistemas de pastizales para sostener la capacidad de vida. En consecuencia, según Fornara y Tilman (2009), Jarvis et al. (2010) y Rivera, Arbrecht y Calle (2013), si dichas acciones se mantienen o sus excesos perduran en el tiempo, los sistemas tienden a declinar la diversidad, la capacidad de ofrecer servicios ecosistémicos, o pueden colapsar.
Según señalan DeClerck (2011), Iraola (2014) y Montagnini (2015), los arreglos forestales, entre ellos los sistemas silvopastoriles y agrosilvopastoriles, frecuentemente mantienen una mayor diversidad de especies y, por ende, una mayor diversidad funcional; y, en teoría, conservan los servicios ecosistémicos previstos.
En este sentido, algunas investigaciones realizadas por Alonso et al. (2007) demostraron un aumento significativo en la riqueza de especies y el índice de diversidad biológica de Shannon, en la medida que se desarrolló un sistema silvopastoril compuesto por leucaena-guinea para la producción de leche bovina. Así, el objetivo de esta investigación fue evaluar el comportamiento faunístico en un sistema silvopastoril destinado al ganado de engorde.
MATERIALES Y MÉTODOS
Localización y condiciones edafoclimáticas. La investigación se desarrolló en áreas experimentales del Instituto de Ciencia Animal, situado a los 22° 53' de latitud norte y los 82° 02' de longitud oeste, y a 92 msnm, en el municipio San José de las Lajas _provincia Mayabeque, Cuba_, sobre un suelo Pardo con carbonato (Hernández et al., 1999); dichas áreas están dedicadas a la producción de bovinos de engorde en pastoreo.
La temperatura promedio en los años que duró la investigación fue de 23 °C, la máxima alcanzó 34 °C en agosto y la mínima hasta 5 °C en enero. La precipitación anual alcanzó los 1 426 mm, con los valores más altos en julio (244,6 mm). Los valores registrados fueron similares a los promedios históricos de los últimos 43 años (datos tomados de la estación meteorología del Instituto de Ciencia Animal de Cuba, 2014).
Características generales del área experimental. Se desarrolló una estrategia de restauración en un sistema de pastizales para bovinos de engorde en pastoreo, con una superficie física de 50 ha; aproximadamente el 60 % de esta área se caracteriza como degradada, como consecuencia del sobrepastoreo antes de comenzar la investigación. El sistema estaba dividido en tres cuartones y mantenía una carga animal de 2,5 cabezas por hectárea. No presentaba árboles maderables ni frutales, solo algunos cercos vivos y menos de 0,75 árboles dispersos por hectárea.
Se utilizó como modelo de cobertura vegetal un sistema silvopastoril, adecuado al contexto donde se desarrolló la investigación. El proceso de restauración conllevó: a) desarrollar una arquitectura de plantas por estratos de forma gradual, con mayor diversificación de la vegetación en el espacio físico; b) incluir gramíneas mejoradas, árboles maderables (590), árboles frutales (493), cercos vivos y leucaena en asociación con gramíneas, con densidad de población entre 5 000 y 7 000 plantas por hectárea y podas de mantenimiento cada 2,5 años, sin el uso de irrigación ni fertilizantes químicos; c) dejar desde tres hasta 40 cuartones con acceso limitado del ganado, reposo durante más de 50 días en el periodo lluvioso (PLL) y no menos de 60 días en el periodo poco lluvioso (PPLL), con periodos cortos de ocupación (de uno a dos días por cuartón); d) realizar las transformaciones sin detener el flujo zootécnico de cuatro ciclos productivos con genotipos Cebú y sus mestizos (1,48; 1,60; 1,80 y 1,80 animales ha-1 año-1, respectivamente), que lograron un adecuado crecimiento (desde 250 kg hasta 420 kg de PV), con buen estado físico y de salud al sacrificio; e) proporcionar, con la diversidad de la comunidad de plantas, diversos servicios ecosistémicos en el sistema.
Procedimiento experimental. Se monitoreó durante cinco años el comportamiento faunístico en el sistema. Para ello se aplicó un modelo lineal mixto y se utilizó el macro GLIMMIX mediante el software SAS versión 9.1 (2007).
En el modelo lineal mixto se consideraron, como efecto fijo, el año, la época y la interacción año por época, y como efecto aleatorio los cuadrantes. La ecuación del modelo mixto (MM) fue la siguiente:
yjkl = ì + ai + ßj + (a ß)j + Fk + eijkl
Donde:
yjkl = f (µ) valor de los individuos esperados
µ = media general o intercepto
ai = efecto fijo del i-ésimo año (i = 1,..., 5)
ßj = efecto fijo de la j-ésima época (j = 1 y 2)
(a ß)j=efecto del i-ésimo año (i = 1,..., 5) en la j-ésima época (j = 1 y 2)
Fk = efecto aleatorio asociado del k-ésimo cuadrante (k = 1, 2,..., 4)
eijkl = error aleatorio asociado a las observaciones
Mediciones. Con el propósito de observar las variaciones entre los individuos a medida que se avanzó en la restauración del sistema, se realizó una evaluación por época. Para ello se adaptó el método de transeptos, descrito por Taylor (2003). El sistema fue dividido en cuatro cuadrantes de 12,5 ha, cada uno de los cuales incluyó 10 cuartones de 1,25 ha. Se recorrió al azar en transeptos y se tomaron dos puntos por cuartón, con un radio aproximado de 7 m y una separación mínima de 25 m; en estos el observador, durante 15 minutos, cuantificó las aves observadas y también otras especies (batracios, roedores y reptiles). Entre un punto y otro se esperó hasta 15 minutos para realizar el muestreo, y también se contó con 5 minutos de adaptación antes de cuantificar las especies en los horarios matutino (7:00 a. m.-9:00 a. m.), del mediodía (11:30 a. m.-12:30 p. m.) y vespertino (4:30 p. m.-6:30 p.m.), con una frecuencia de tres días consecutivos por época en cada cuadrante.
Se calcularon los índices ecológicos número de individuos por especie (expresado en total), riqueza de especies y diversidad, y se determinó el porcentaje de aves por año. Se adaptaron los cálculos por el método propuesto por Fontenla, Rodríguez y Suri (1987), y se utilizaron las fórmulas clásicas de Shannon-Weaver (1949).
El porcentaje de árboles maderables y frutales que sobrevivieron se determinó mediante conteo individual, al final de cada periodo de lluvia. La producción de biomasa herbácea se estimó por el método propuesto por Haydock y Shaw (1975), y se realizaron 100 observaciones por cuartón. En el caso de la disponibilidad de leucaena, se adecuó el método propuesto por Mahecha et al. (2000): R2aj = 0,85. El cálculo de la biomasa (toneladas de MS) se realizó con ayuda del software CALRAC (Roche et al., 1999). Se ponderó un índice de capacidad de carga biológica (ICCB) por año para este sistema, expresado entre los individuos que se cuantificaron (incluidos los bovinos) y la biomasa.
Para los efectos que resultaron significativos en el modelo (p < 0,05) se aplicó la prueba de comparación múltiple para las medias mínimas cuadráticas, según la dócima de Tukey-Kramer (Kramer, 1956). Las variables número de individuos y riqueza se transformaron según vx. Se realizó análisis de regresión simple para explorar la relación en el tiempo entre las características del total de árboles presentes (maderables, frutales, árboles dispersos y cercos vivos), la biomasa y la fauna cuantificada en el sistema silvopastoril (número de individuos totales y diversidad) mediante el software SAS versión 9.1 (2007).
RESULTADOS Y DISCUSIÓN
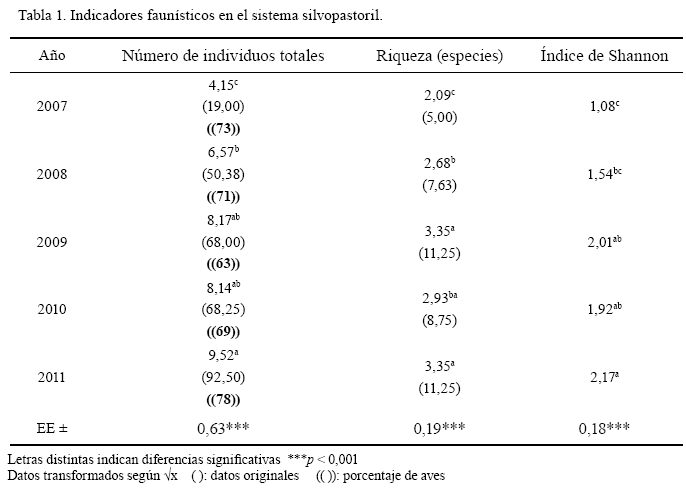
La tabla 1 muestra el comportamiento faunístico, su estabilidad y su estado durante los cinco años de evolución y desarrollo de la comunidad de plantas, como un indicador de la mejora del equilibrio ecológico. Se encontró diferencia (p < 0,001) para el efecto año en el número de individuos (aves, reptiles, batracios y roedores) entre los dos años iniciales; el segundo no difirió del tercero y el cuarto difirió del quinto, en el que se encontraron 73 individuos totales por encima del número inicial (19) en el primer año; entre estos se destacó la presencia de diferentes especies de aves. Por su parte, los efectos época y la interacción año por época no fueron significativos en el modelo.
Para el indicador riqueza se encontró diferencia (p < 0,001) entre los años. También se observó diferencia (p < 0,001) para el índice de Shannon, y en el quinto año de la investigación este indicador aumentó desde 1,08 hasta 2,17.
En el aumento del número de individuos y de la riqueza de especies en el tiempo incidió la presencia de las siguientes aves: sabanero común (Spiza americana), tomeguín (Tiaris olivaceus), garza ganadera (Bubulcus ibis), sinsonte (Mimus polyglottos), codorniz (Coturnix coturnix), entre otras, que representaron entre el 63 y 78 % de los individuos.
La respuesta en cantidad y diversidad de la fauna evidenció la capacidad del sistema silvopastoril, el cual les confirió condiciones apropiadas a las distintas formas de vida que utilizan fuentes de alimento diferentes. Por consiguiente, se infiere que aumentaron en el sistema las posibilidades de autorregulación entre las poblaciones y estas manifestaron determinado grado de estabilidad. Dicho estado se observó para los indicadores riqueza y diversidad a partir del cuarto año, dado fundamentalmente por la presencia de la leucaena y los árboles dispersos, controlados por las podas de mantenimiento. No obstante, en el quinto año se observó en todos los indicadores un crecimiento relativo, que pudo estar relacionado con la capacidad de resiliencia que los agroecosistemas agroforestales pueden ofrecer ante las perturbaciones, según plantean Rivera et al. (2013).
De acuerdo con lo señalado por Kimberly (2012), un comportamiento faunístico adecuado puede estar relacionado con las mejores posibilidades de hábitats que proporciona la diversidad de la cobertura vegetal, que a su vez posibilita mayores servicios ecosistémicos directos. En tal sentido, para el quinto año se incrementó en un 60 % la producción de biomasa herbácea, la presencia de árboles maderables en desarrollo (66 % de sobrevivencia) y frutales (44 % de sobrevivencia), así como los cercos vivos (de 2 árboles ha-1 hasta 4 árboles ha-1) y los árboles dispersos (de 117 hasta 608).
La presencia de los árboles dispersos en el sistema posibilitó mejorar las condiciones de sombra para los animales, entre otros beneficios ecosistémicos. Sin embargo, al concluir esta investigación el 53 % de ellos aún estaban en fases juveniles, y se logró como promedio 9,58 árboles ha-1 distribuidos de manera aleatoria. Según Iraola et al. (2011) e Iraola et al. (2014), este sistema demostró la posibilidad de introducir una mayor cantidad de árboles leñosos considerados de protección, que contribuirían a proporcionar en el tiempo un mayor aporte de biomasa y un mayor equilibrio ecológico en el sistema.
En correspondencia con lo argumentado, Cárdenas (2002), Carpenter et al. (2009) y Marinidou et al. (2013) plantearon que una mayor presencia de componente arbóreo y la abundante cobertura vegetal herbácea en los sistemas de pastizales indican mayores posibilidades de recursos para la presencia y refugio de la fauna. Así lo confirman algunos estudios realizados en Cuba por Alonso (2011) y en Colombia por Fajardo et al. (2009), donde se monitorearon varios sistemas silvopastoriles y sistemas de gramíneas. Se halló una diversidad de Shannon superior a 0,83 en silvopastoreo, mientras que en los sistemas de gramíneas sin árboles esta llegó solo a 0,34. En tal sentido, se coincide con estos autores en que la presencia de un componente arbóreo dentro de los sistemas ganaderos beneficia el incremento de la fauna y, a su vez, favorece una mejor conexión entre los agroecosistemas para las distintas formas de vida silvestre.
Por tanto, con un manejo adecuado por parte del hombre, la presencia de árboles contribuiría a mejorar la vida útil de los pastizales, a incrementar la diversidad funcional y a sostener determinada capacidad de carga biológica, cambiante en el tiempo y condicionada por numerosos factores en cada agroecosistema. La figura 1 muestra el ICCB ponderado para este sistema a partir de los individuos observados.
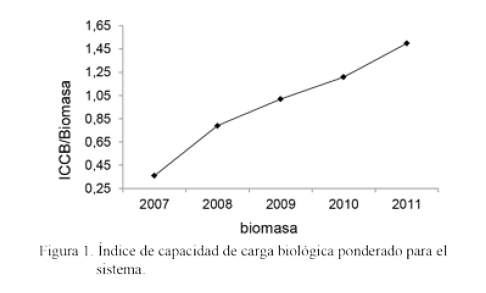
Se encontró una mejora en el índice de capacidad de carga biológica (desde 0,36 hasta 1,50); de ahí que la recuperación se alcanzara en armonía con el incremento de la carga animal de bovinos y las otras formas de vida vegetal y animal con las que se relacionó directa o indirectamente en el sistema.
Según Garbach et al. (2012) e Iraola (2014) el ICCB está definido no solo por la cantidad, diversidad, producción de nutrientes y energía de sus poblaciones vegetales, sino por su capacidad para retener y suministrar agua y propiciar hábitats favorables para el desarrollo de la vida, en los cuales cohabitan y comparten los diferentes individuos en determinado sistema. Así lo confirman las condiciones favorables que se propiciaron con el desarrollo de este sistema silvopastoril, expresado por el ICCB.
Por su parte, se encontraron relaciones positivas, con coeficientes de determinación superiores en todos los casos al 80 %, entre el componente arbóreo, la biomasa y los índices ecológicos número de individuos totales y diversidad (fig. 2). Ello indica que existió una estrecha relación entre el componente arbóreo que fue fomentándose en el sistema y el incremento de la biomasa, los cuales evidentemente tuvieron un efecto importante en la fauna, utilizada como indicador ecológico y de salud para mostrar la evolución del sistema durante los cinco años de investigación.
A partir de los resultados se evidenció el potencial que se puede alcanzar al utilizar un modelo de cobertura vegetal como los sistemas silvopastoriles. Ello coincide con lo informado por Lang et al. (2003) en Costa Rica, donde la presencia de los árboles y el incremento de la biomasa influyeron positivamente en los índices ecológicos. Otros resultados relacionados con estos índices ecológicos fueron reportados en Cuba por Alonso (2003) y Alonso et al. (2004), en silvopastoreo con más de 10 años de explotación para la producción lechera, donde se alcanzaron incrementos favorables de la biodiversidad.
La diversidad que pudo ofrecer este sistema silvopastoril, utilizado como modelo de cobertura, confirmó las ventajas dinámicas de las interacciones en dicho sistema, con una mayor capacidad de hábitats para la fauna asociada y posibilidades para la crianza animal. Sin embargo, de acuerdo con lo planteado por Vargas (2008), Laliberté y Legendre (2010) y Mouchet et al. (2010), no existen los índices ecológicos ideales que pudieran explicar todas las situaciones en los agroecosistemas. Tampoco existe un criterio universal sobre cuáles son los caracteres más importantes a medir dentro de un sistema; sino que la utilidad de los índices que se cuantifican, para poder monitorear diferentes aspectos de la diversidad funcional, depende en gran medida de los objetivos concebidos en cada investigación.
En correspondencia con lo anterior y de acuerdo con autores como Díaz et al. (2007), DeClerck (2011), Lavorel et al. (2011), Schneiders et al. (2012) y Montagnini (2015), un manejo adecuado por parte del hombre y una mayor diversidad de plantas (gramíneas, leguminosas, arbustos y árboles), conjuntamente con los componentes (suelo-animal-clima-hombre), permitirían lograr estos propósitos que se mencionan y contribuir a la capacidad del sistema de proveer servicios ecosistémicos y respuestas ante las perturbaciones. Ello no solo garantiza la presencia y refugio de las diferentes formas de vida, sino también genera una mayor capacidad de sostener vida, donde convergen muchas especies que pueden cohabitar en los agroecosistemas de pastizales. Por ende, hay que enfocar la restauración de los sistemas ganaderos hacia una ganadería más amigable con el medio ambiente, que pueda satisfacer las necesidades ecológicas, productivas y económicas de los seres humanos.
Se concluye que el sistema silvopastoril, como modelo de cobertura, posibilitó hábitats más amplios para el desarrollo faunístico y mejoró la relación de los indicadores ecológicos con la presencia de los árboles y el incremento de la producción de biomasa en el sistema.
AGRADECIMIENTOS
Se agradece al departamento de Biomatemática y a los trabajadores de la finca de ceba bovina en pastoreo Cebadero Ayala, del Instituto de Ciencia Animal.
REFERENCIAS BIBLIOGRÁFICAS
1. Alonso, J. Factores que intervienen en la producción de biomasa de un sistema silvopastoril leucaena (Leucaena leucocephala cv. Perú) y guinea (Panicum maximum cv. Likoni). Tesis presentada en opción al grado de Doctor en Ciencias Agrícolas. Cuba: Instituto de Ciencia Animal, 2003.
2. Alonso, J. Los sistemas silvopastoriles y su contribución al medio ambiente. Rev. cub. Cienc. agríc. 45:107-115, 2011.
3. Alonso, J.; Torres, O.; Ruíz, T. E.; Febles, G.; Cárdenas, G. & Achan, G. Estudio de la avifauna asociada a un sistema silvopastoril leucaena-guinea con diferentes edades de establecimiento. Rev. cub. Cienc. agríc. 38:203-210, 2004.
4. Alonso, J.; Valenciaga, Nurys; Sampaio, R. A. & Leite, G. L. D. Diversidad zoológica asociada a un silvopastoreo leucaena-guinea con diferentes edades de establecimiento. Pesq. agropec. bras. 42 (12):1667-1674, 2007.
5. Cárdenas, G. Cobertura arbórea y diversidad de aves en paisajes fragmentados en Cañas, Costa Rica. Tesis Mag. Sc. Turrialba, Costa Rica: CATIE, 2002.
6. Carpenter, S. R.; Mooney, H. A.; Agard, J.; Capistrano, Doris; DeFries, Ruth S.; Díaz, Sandra et al. Science for managing ecosystem services: Beyond the Millennium Ecosystem Evaluation. Proceedings of the National Academy of Sciences. 106 (5):1305-1312, 2009.
7. DeClerck, F. Biodiversidad funcional en sistemas agroforestales. Agroforestería Neotropical. 1 (1):2-4, 2011.
8. Díaz, Sandra; Lavorel, Sandra; Bello, F. de; Quétier, F.; Grigulis, K. & Robson, T. M. Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Sciences. 104 (52):20684-20689, 2007.
9. Fajardo, D.; Johnston-González, R.; Nerira, L.; Chará, J. & Murgueitio, E. Influencia de los sistemas silvopastoriles en la diversidad de aves en la Cuenca del rio La Vieja. Recursos Naturales y Ambiente. (58):9-16, 2009.
10. Fontenla, J. L.; Rodríguez, R. & Suri, M. Estructura y organización de dos comunidades de Coccidae (Insecta: Homoptera) en dos cultivares de cítricos. La Habana: Instituto de Ecología y Sistemática, 1987.
11. Fornara, D. A. & Tilman, D. Ecological mechanisms associated with the positive diversity-productivity relationship in an N-limited grassland. Ecology. 90 (2):408-418, 2009.
12. Garbach, K.; Lubell, M.; Fabrice, M. & DeClerck, A. J. Payment for ecosystem services: The roles of positive incentives and information sharing in stimulating adoption of silvopastoral conservation practices. Agr. Ecosyst. Environ. 156:27-36, 2012.
13. Haydock, K. P. & Shaw, N. H. The comparative yield method for estimating dry mather yield of pasture. Aust. J. Exp. Agric. Anim. Husb. 15:663-670, 1975.
14. Hernández, A.; Pérez, J. M.; Bosch, D. & Rivero, L. Nueva versión de la clasificación genética de los suelos de Cuba. La Habana: AGRINFOR, 1999.
15. Iraola, J. Rediseño y manejo de un arreglo agroforestal para mejorar la capacidad de carga biológica con ganado de engorde. Tesis en opción al grado de Doctor en Ciencias Veterinarias. Mayabeque, Cuba: Instituto de Ciencia Animal, 2014.
16. Iraola, J.; Hernández, J. L.; Muñoz, E.; Díaz, A. & Moreira, E. Árboles maderables y frutales en sistemas silvopastoriles con ganado de carne. Nota técnica. ACPA. (1):24, 2011.
17. Iraola, J.; Muñoz, E.; García, Y.; Hernández, J. L. & Moreira, E. Estrategia de rediseño ecológica para la recuperación de pasturas degradadas en un sistema agrosilvopastoril de engorde bovino. Memorias III Convención Internacional Agrodesarrollo 2014. (Eds. J. M. Iglesias, J. Suárez y Nayda Armengol). Matanzas, Cuba: EEPF Indio Hatuey. p. 1040-1043, 2014.
18. Jarvis, A.; Touval, J. L.; Castro, M.; Sotomayor, L. & Hyman, G. G. Assessment of threats to ecosystem in South America. J. Nat. Conserv. 18 (3):180-188, 2010.
19. Kimberly, F. The remarkable resilience of ant assemblages following major vegetation change in an arid ecosystem. Biol. Conserv. 148:96-105, 2012.
20. Kramer, C. Y. Extention of multiple range tests to group means with unequal numbers of replications. Biometrics. 12:307-310, 1956.
21. Laliberté, E. & Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology. 91:299-305, 2010.
22. Lang, I.; Gormley, L.; Harvey, Celia A. & Sinclair, F. Composición de la cominidad de aves en cercas vivas de Río Frío, Costa Rica. Agroforestería en las Ámericas. 39-40:86-92, 2003.
23. Lavorel, Sandra; Grigulis, K.; Lamarque, Penélope; Colace, Marie P.; Garden, D.; Girel, J. et al. Using plant functional traits to understand the landscape distribution of multiple ecosystem services. J. Ecol. 99 (1):135-147, 2011.
24. Mahecha, Liliana; Duran, C.; Rosales, M.; Molina, C. H. & Molina, E. Consumo de pasto estrella africana (Cynodon plectostachyus) y leucaena (Leucaena leucocephala) en un sistema silvopastoril. Pasturas Tropicales. 22 (1):26-30, 2000.
25. Marinidou, E.; Finegan, B.; Jiménez-Ferrer, G.; Delgado, D. & Casanoves, F. Concepts and a methodology for evaluating environmental services from trees of small farms in Chiapas, México. J. Environ. Manage. 114:115-124, 2013.
26. Montagnini, Florencia. El papel de los sistemas agroforestales en la restauración a la escala del paisaje. VIII Congreso Internacional de Sistemas Agroforestales para la Producción Pecuaria y Forestal Sostenible y III Congreso Nacional de Sistemas. Silvopastoriles. Iguazú, Misiones, Argentina, 2015.
27. Mouchet, M. A.; Villéger, S.; Mason, N. W. & Mouillot, D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Funct. Ecol. 24:867-876, 2010.
28. Rivera, L. F.; Armbrecht, I. & Calle, Z. Silvopastoral systems and ant diversity conservation in a cattle-dominated landscape of the Colombian Andes. Agr. Ecosyst. Environ. 181:188-194, 2013.
29. Roche, A.; Larduet, R.; Torres, V. & Ajete, A. CALRAC. Programa de computación para el cálculo de raciones en rumiantes. Rev. cub. Cienc. agric. 33:13-19, 1999.
30. SAS. SAS User's guide: Statistics. Version 9.1.3. Cary, NC, USA: SAS Institute, 2007.
31. Schneiders, A.; Daele, T. V.; Landuyt, W. V. & Reeth, W. V. Biodiversity and ecosystem services: Complementary approaches for ecosystem management? Ecol. Indic. 21:123-133, 2012.
32. Shannon, C. E. & Weaver , W. The mathematical theory of comunication. Urbana, Illinois: University of Illinois Press, 1949.
33. Taylor, Rachel. ¿Cómo medir la diversidad de aves presentes en los sistemas agroforestales? Agroforestería en las Ámericas. 39-40:117-123, 2003.
34. Vargas, S. Rediseño, manejo y evaluación de un agroecosistema de pastizal con enfoque integrado, para la producción de leche bovina. Tesis en opción al grado de Doctor en Ciencias Veterinarias. Villa Clara, Cuba: Universidad Central Marta Abreu de Las Villas, 2008.
Recibido el 16 de abril del 2015
Aceptado el 24 de septiembre del 2015


{kind=link}
{kind=link}