










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Protección Vegetal
Print version ISSN 1010-2752On-line version ISSN 2224-4697
Rev. Protección Veg. vol.25 no.3 La Habana Sept.-Dec. 2010
TRABAJO ORIGINAL
DIVERSIDAD GENÉTICA DE ESPECIES SILVESTRES DEL GÉNERO Nicotiana II: CARACTERIZACIÓN MOLECULAR MEDIANTE MARCADORES RAPD
GENETIC DIVERSITY OF WILD Nicotiana SPECIES II: MOLECULARCHARACTERIZATION USING RAPD MARKERS
Marlyn Valdés de la Cruz*, Yuniet Hernández**, Alejandra Vázquez-Lobo***, Daniel Piñeiro*** y Lien González-Pérez*
*Facultad de Biología, Universidad de La Habana, Calle 25 No. 455 entre I y J Vedado, Ciudad de La Habana, Cuba. Correo electrónico: marlyn@fbio.uh.cu; **Instituto Nacional de Ciencias Agrícolas (INCA), Carretera a Tapaste km 3.5, San José de las Lajas, La Habana, Cuba; ***Instituto de Ecología, Universidad Autónoma de México (UNAM), 3er Circuito Exterior. Ciudad Universitaria. Delegación Coyoacán. Distrito Federal, México
RESUMEN
Las especies silvestres del género Nicotiana son reservorios de genes de resistencia a plagas, así como para rasgos cualitativos y fitoquímicos importantes, los cuales no están presentes en las variedades cultivadas. El objetivo de trabajo fue caracterizar la variabilidad genética de 10 especies silvestres del género Nicotiana mediante el uso de marcadores RAPD (Fragmentos Polimórficos de ADN Amplificados al Azar). De los diez cebadores usados en el ensayo RAPD, cuatro de ellos produjeron productos de amplificación que resultaron apenas visibles para ser evaluados o no fueron consistentemente reproducibles y seis cebadores produjeron un total de 117 bandas, 100% polimórficas. Todas las bandas se evaluaron para la comparación de la amplificación, para un promedio de 19,5 bandas por cebador. El mayor nivel de polimorfismo entre las especies se observó en Nicotiana velutina Wheelen y Nicotiana glutinosa L. con 35,89% y 32,47% respectivamente, ambas del subgénero Petunioides. En contraste, el menor porcentaje de polimorfismo se observó en la especie Nicotiana knightiana Goodsp (18,80%), perteneciente al subgénero Rustica. El patrón obtenido en el dendrograma coincide con la clasificación tradicional de las especies del género Nicotiana. En nueve, de las 10 especies silvestres del género Nicotiana evaluadas, todos los cebadores generaron al menos dos bandas específicas. El total de los 32 marcadores específicos de cada especie encontrados en el presente estudio podría resultar de gran utilidad en los programas de mejoramiento genético.
Palabras clave: Nicotiana; diversidad genética; polimorfismo; ADN polimórfico amplificado al azar (RAPD)
ABSTRACT
Wild Nicotiana species are reservoir of genes for several pests, besides other genes for important phytochemical and quality traits, which are not present in cultivated varieties. The application of Random Amplified Polymorphic DNA (RAPD) analysis to determine the degree of genetic variation among ten wild Nicotiana species is herein presented. Four of the ten primers used in RAPD analysis, produced amplification products that were too faint to score or could not be consistently reproduced and six primers yielded a total of 117 bands, all of them polymorphic. All the bands were scored from the comparison of amplification, with an average of 19.5 bands scored per primer. The highest level of polymorphism among the species was observed for Nicotiana velutina Wheelen and Nicotiana glutinosa L., with 35.89% and 32.47% respectively, both from subgenus Petunioides. In contrast, the lowest level of polymorphism (18,80%) was observed for Nicotiana knightiana Goodsp, subgenus Rustica. The clustering pattern matched with the traditional classification of Nicotiana species. In nine of the ten wild Nicotiana species evaluated, all the primers generated at least two specifics bands. The total of the 32 species-specific markers identified in the present study will be useful in the genetic breeding programs.
Key words: Nicotiana; genetic diversity; polymorphism; Random Amplified Polymorphic DNA (RAPD) markers
(Recibido 4-3-2010; Aceptado 17-5-2010)
INTRODUCCIÓN
La caracterización de los recursos fitogenéticos del género Nicotiana está considerada entre las líneas de investigación estratégicas del cultivo a nivel mundial. Esta constituye un factor decisivo en la solución de los problemas actuales y futuros relacionados con la productividad de los materiales comerciales, la adaptación a los cambios climáticos y el desarrollo de nuevas alternativas en la obtención de variedades mediante la utilización de métodos tradicionales y biotecnológicos. Para ello es esencial, en los programas de mejoramiento genético, la información que se tenga sobre la diversidad genética dentro y entre especies estrechamente relacionadas y de esta manera garantizar el uso racional de los recursos genéticos (1,2).
Con este fin, la estrategia de crear bancos de germoplasma ha permitido conservar el patrimonio genético vegetal y, por tanto, la variabilidad genética de cada especie y género, lo que constituye el fundamento esencial de los programas de mejoramiento genético y de selección de genotipos (3).
La medida más directa de la variabilidad genética reside en el análisis del ADN en cada individuo de la población o de una muestra de ella. En tal sentido, el desarrollo de los marcadores moleculares ha supuesto un importante avance en la mejora de las plantas, pues durante las últimas décadas los mismos han venido complementando los métodos clásicos de cruzamiento y selección (4). De esta manera, han permitido localizar genes y marcarlos, por lo que constituyen una potente herramienta biotecnológica y una alternativa para obtener nuevas variedades (5,6).
Los marcadores moleculares presentan una serie de ventajas frente a los marcadores morfológicos ya que se encuentran en cualquier especie, se detectan en estadios tempranos del desarrollo y en cualquier tipo de tejido, su análisis no depende de las condiciones experimentales, son fenotípicamente neutros y la interacción entre ellos es pequeña o nula (ausencia de epistasia) (7).
Dentro de los marcadores moleculares descritos, los Fragmentos Polimórficos de ADN Amplificados al Azar (RAPD) se basan en la utilización de un único oligonucleótido de 10pb que hibrida al azar con el ADN en estudio, por lo que la secuencia amplificada es desconocida. El polimorfismo que se observa entre distintos individuos consiste por lo tanto, en la presencia o ausencia de fragmentos de ADN amplificado (4,6). Estos marcadores han sido utilizados ampliamente para el estudio de la diversidad genética intra e inter específicas en diferentes géneros de plantas, como son: Nicotiana (8,9,10), Gossypium (11), Camellia (12), Solanum (13). Entre sus ventajas se destacan, que no requiere del conocimiento previo del genoma estudiado y que es una técnica rápida, sencilla y de bajo costo de implementación (6).
Atendiendo a estos antecedentes, el objetivo de este estudio fue caracterizar la variabilidad genética de 10 especies silvestres del género Nicotiana mediante el uso de marcadores RAPD.
MATERIALES Y MÉTODOS
Material vegetal y extracción de ADN
La procedencia y características del material vegetal en estudio aparecen descritas en Valdés de la Cruz et al. (14). Las plantas se obtuvieron a partir de semilla botánica, las cuales se sembraron en bandejas de poliestireno expandido con alvéolos (cepellones) que contenían una mezcla de suelo y materia orgánica a temperatura entre 20 y 25°C y luz natural.
A los 25 días de edad, se realizó la extracción de ADN genómico, de forma independiente a cinco individuos por especie. Para ello se partió de 100mg de tejido foliar y se desarrolló el procedimiento según la metodología descrita por Aldrich y Cullis (15), que emplea tampón basado en el Bromuro de amonio hexadeciltrimetilo (CTAB).
La integridad del ADN aislado se determinó por electroforesis en geles de agarosa al 0,8% y tinción con bromuro de etidio. La concentración del ADN se calculó a través de la medida de absorbancia a la longitud de onda de 260 nm y la calidad se determinó mediante la relación 260/280 nm, empleando para ello un espectofotómetro (Ultrospec 2100 pro, Amersham Pharmacia Biotech) (16). A partir de los resultados obtenidos se emplearon las tres mejores preparaciones de ADN por especie para los análisis moleculares y se conservaron a -20°C hasta su utilización.
Amplificación por RAPD
Las amplificaciones se desarrollaron en un volumen final de 25µL para una reacción que contenía 20ng de ADN genómico molde, 0,4µM del cebador de RAPD, 0,2mM de la mezcla de dNTP, 1,5mM de cloruro de magnesio, 1X del tampón de la enzima y 0,5U de Taq polimerasa (Promega). Se evaluaron 10 cebadores RAPD, nueve de la serie A y uno de la serie F (Operon Technologies, Alameda, CA, USA). Los mismos fueron: OPA 01 (CAGGCCCTTC), OPA 02 (TGCCGAGCTG), OPA 03 (AGTCAGCCAC), OPA 10 (GTGATCGCAG), OPA 13 (CAGCACCCAC), OPA 15 (TTCCGAACCC), OPA 16 (AGCCAGCGAA), OPA 17 (GACCGCTTGT), OPA 18 (AGGTGACCGT) y OPF 20 (GGTCTAGAGG). En cada reacción de amplificación se analizaron las tres muestras por especie descritas previamente y se realizaron tres repeticiones de las amplificaciones por cebador.
Las reacciones de PCR se desarrollaron en un termociclador Thermal cycler 2720 (Applied Biosystems), mediante el siguiente programa de amplificación: un ciclo inicial de 94°C por 4 min., seguido de 40 ciclos a 94°C (1 min.), 35°C (1 min.) y 72°C (2 min.), con un ciclo final de extensión a 72°C durante 7 min. Los productos de PCR se analizaron en geles de agarosa al 1,5% en 0,5X de tampón TBE (Tris hidroximetil aminometano (Tris-HCl) 0.04M; ácido etilendiamino-tetra-acético (EDTA) 1mM; ácido bórico 0.04M; pH 8), a 100v de voltaje constante. Las bandas se visualizaron por tinción con bromuro de etidio en un transiluminador de luz U.V. (High Performance Ultraviolet Transiluminator UVP) (16). La talla de los fragmentos amplificados se determinó por comparación con un marcador de peso molecular de ADN de 1Kb plus (Invitrogen).
Análisis de datos
Primeramente se calculó el porcentaje de polimorfismo que permite detectar cada cebador RAPD, teniendo en cuenta el total de bandas polimórficas que se obtuvo para cada uno de ellos, sobre la sumatoria de bandas polimórficas obtenidas con todos los cebadores (total) por 100 (10). Posteriormente, se realizó este mismo cálculo para determinar la variabilidad genética de cada especie, en este caso teniendo en cuenta las bandas polimórficas por especie.
La confección y procesamiento de la matriz binaria, coeficiente utilizado y método para producir el dendrograma fueron similares a los descritos por Valdés de la Cruz et al. (14). Para la formación de los grupos se consideró el índice de similitud de Dice (17), que permitió el mejor agrupamiento entre las especies.
RESULTADOS
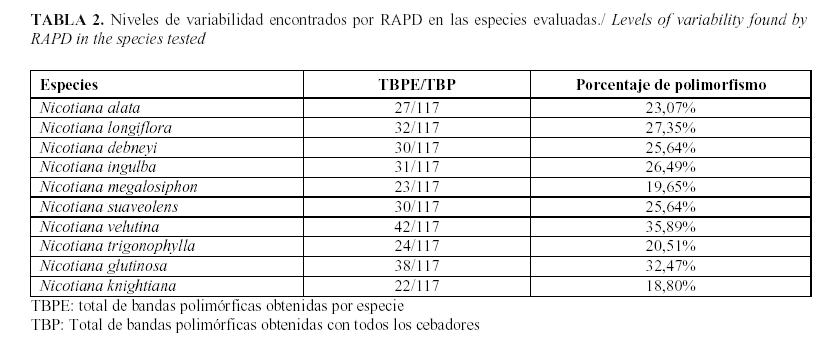
De los diez cebadores RAPD empleados para el análisis, cuatro de ellos produjeron productos de amplificación que resultaban apenas visibles para ser evaluados o no fueron consistentemente reproducibles. Por lo tanto, del total analizado sólo seis resultaron apropiados para observar los patrones de polimorfismo, con adecuada resolución y reproducibilidad. En la Tabla 1 se puede apreciar el número total de bandas obtenidas con cada cebador, en todos los casos se obtuvo un polimorfismo del 100% para un total de 117 bandas polimórficas.
El número de fragmentos amplificados para cada cebador estuvo entre 14 y 26 bandas, para un promedio de 19,5 fragmentos por cebador entre los dos subgéneros y cinco secciones de especies silvestres del género Nicotiana evaluados. La talla de los fragmentos obtenidos estuvo en el rango de 150 y 2500pb. Como ejemplo, en la Figura 1 se muestran los perfiles de bandas obtenidos con los cebadores OPA 16 y OPF 20. El número máximo de bandas se obtuvo con el cebador OPF 20, mientras que el menor correspondió con el cebador OPA 18.
Con respecto al polimorfismo detectado entre especies con el empleo del marcador RAPD, se pudo evidenciar que el mayor nivel de variabilidad se observó en las especies N. velutina Wheelen y N. glutinosa L., ambas del subgénero Petunioides, con porcentajes de polimorfismo de 35,89% y 32,47%, respectivamente. Mientras que la menor variabilidad se observó en la especie N. knightiana Goodsp (18,8%), perteneciente al subgénero Rustica (Tabla 2).
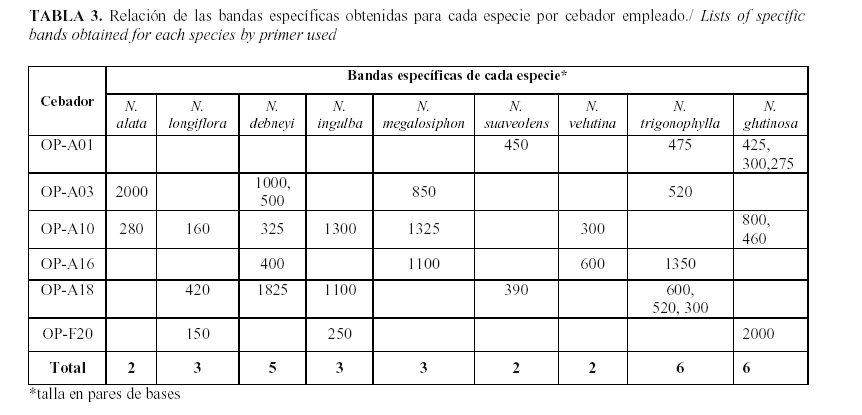
Al analizar los patrones de polimorfismo obtenidos por especie se pudo apreciar que para todos los cebadores se obtienen, al menos, dos bandas específicas por especie, excepto para N. knightiana, del subgénero Rustica (Tabla 3). De todos los cebadores evaluados el OPA-10 resultó ser el más eficiente en este sentido, pues permitió obtener bandas específicas para siete de las especies, además que permitió la detección de 24 bandas polimórficas (Tabla 1).
Por último, el análisis de conglomerados con estas especies realizado a partir de los resultados de las matrices binarias se muestra en la Figura 2. Los grupos en el dendrograma se formaron a partir del 0,40 de índice de similitud, conducen a la formación de seis grupos.
El grupo I lo integran las especies N. longiflora Cav y N. alata Link et al. Otto, ambas miembros de la sección Alatae con números cromosómicos de 2n=20 y 2n=18, respectivamente. En el grupo II se incluye la especie N. trigonophylla Dunal, perteneciente a la sección Trigonophyllae, con 2n=24 y el III también está formado por una sola especie, en este caso N. glutinosa de la sección Undulatae, con 2n=24. En el grupo IV se encuentran las especies N. debneyi Domin y N. suaveolens Lhem con números de cromosomas 2n=48 y 2n=32, respectivamente. Por su parte, el grupo V reúne a las especies N. ingulba Black, N. megalosiphon Van Huerck y Mull. Arg y N. velutina, cuyos números cromosómicos son de 2n=40 para las dos primeras y 2n=32 para la tercera.
Las especies que integran los grupos IV y V pertenecen a la sección Suaveolentes y, de manera general, los representantes de los grupos del I al V pertenecen al subgénero Petuniode. Finalmente, el grupo VI está integrado por la especie N. knightiana con número de cromosomas 2n=24, única representante dio de la diversidad genética de especies silvestres que se evaluó del subgénero Rustica.
La línea vertical indica el corte realizado para el agrupamiento teniendo en cuenta el índice de similitud de 0,40.
DISCUSIÓN
El análisis por RAPD fue adecuado para el estudio de la diversidad genética de especies silvestres del género Nicotiana. El promedio de fragmentos amplificados para cada cebador es de 19,5 y supera lo obtenido por Siva Raju et al. (10), quienes encontraron 12,7 fragmentos por cebador al analizar un total de 22 especies de Nicotiana y diferentes cebadores RAPD. Por otra parte, Zhang et al. (1,18) obtuvieron valores de 8,8 y 9,6 fragmentos por cebador al analizar la variabilidad genética en cultivares de N. tabacum empleando cebadores RAPD.
Con respecto a los niveles de polimorfismo, los mayores porcentajes se encontraron en las especies pertenecientes al subgénero Petunioides, el cual incluye especies que contienen números cromosómicos entre 18 y 48, por lo que se espera una alta variabilidad entre ellas. Estos resultados coinciden con lo obtenido por otros autores al evaluar el polimorfismo genético entre especies del género Nicotiana. Tal es el caso de Siva Raju et al. (10), quienes detectaron un 99,07% en este subgénero, al comparar los tres subgéneros mediante la técnica RAPD. Estos autores realizaron la caracterización con cebadores diferentes a los utilizados en este trabajo. Resultados similares fueron confirmados al emplear otros marcadores moleculares al emplear el Polimorfismo de Longitud de los Fragmentos Amplificados (AFLP) (19). Así mismo, Del Piano et al. (8) realizó la comparación de la variabilidad genética entre especies y variedades de Nicotiana, utilizando marcadores de RAPD (8) y de Inter Secuencias Simples Repetidas (ISSR) (20).
El polimorfismo encontrado para el subgénero Petunioides también está en correspondencia con los resultados obtenidos para los marcadores bioquímicos (Valdés de la Cruz et al. (14). En el mismo se evidenció que las especies donde se obtiene el mayor número de isoformas, para la mayoría de los sistemas analizados, fueron N. alata y N. trigonophylla, ambas pertenecientes a este subgénero.
En el caso de la especie N. knightiana (perteneciente al subgénero Rustica), no fue posible encontrar marcadores específicos para la especie con ninguno de los cebadores evaluados. Coincide además en que esta especie posee el menor número de isoformas para la mayoría de los sistemas electroforéticos analizados (14).
Otros autores han encontrado un bajo grado de variabilidad para el subgénero Rustica con respecto a Petunoides (10). No obstante, se debe ampliar el número de especies a analizar para el subgénero y comprobar la variabilidad dentro del mismo. Además, sería recomendable la aplicación de un mayor número de cebadores u otros tipos de marcadores que permitan la identificación de bandas que pudieran corresponder con marcadores específicos de cada especie.
Por otra parte, los 32 marcadores específicos de cada especie pudieran constituir marcadores importantes para la identificación de cada especie. Estas bandas resultarían de gran utilidad en los programas de mejoramiento genético, ya que pueden contribuir a la identificación inequívoca de verdaderos híbridos, monitoreando la introgresión de genes marcados y estimando el fondo genético de segregantes desea
dos, particularmente referido a la contribución genómica de las especies silvestres (5,12).
En el futuro, las bandas específicas de cada especie, pueden ser utilizadas para generar Secuencias Caracterizadas de Regiones Amplificadas o marcadores SCAR, los cuales tienen una mayor sensibilidad y reproducibilidad con respecto a los RAPD (21). El empleo de este marcador más potente eliminaría las desventajas de los RAPD, pues con este se consigue en ocasiones transformar el polimorfismo en codominante y se eliminan los problemas de reproducibilidad (7). Su importancia y aplicaciones han sido demostradas en estudios donde se han obtenido marcadores ligados a genes de resistencia a plagas como el moho azul (Peronospora hyoscyamy fsp. tabacina Adam), al virus Y de la papa (PVY) y la pudrición negra de la raíz (Chalara elegans Nag Raj y W.B. Kender) (22,23).
Al comparar los resultados del dendrograma obtenido por la técnica de RAPD con los informados para los marcadores bioquímicos (14), se puede constatar que los RAPD permitieron detectar una mayor diversidad genética, lo que demuestra el mayor poder discriminante de los marcadores moleculares de ADN (4,7). En consecuencia, a diferencia de lo obtenido para los marcadores bioquímicos, los grupos en el dendrograma se corresponden con las secciones de los subgéneros, sin embargo no siempre se obtuvo el agrupamiento de las especies de una misma sección en un mismo grupo.
Se mantienen juntas las dos especies pertenecientes a la sección Alatae, a pesar de que tienen diferente número cromosómico y comportamiento frente al moho azul. Sin embargo, las especies de la sección Suaveolentes se separan en dos grupos, todas son resistentes al moho azul aunque con diferente número cromosómico. El resto de las especies cada una forma un grupo independiente, ya que pertenecen a secciones diferentes y todas son susceptibles al moho azul. Por lo tanto, no necesariamente el agrupamiento obtenido en el dendrograma guardó relación con la resistencia o susceptibilidad al moho azul y el número cromosómico.
El objetivo de la caracterización de especies silvestres es la identificación de variabilidad genética y su empleo en la selección de posibles progenitores para los programas de mejoramiento que incluyan cruzamientos interespecíficos. En tal sentido resulta de gran importancia lograr un acercamiento a las relaciones filogenéticas entre las especies, pues esto posibilita descartar individuos que compartan caracteres similares y lograr incrementar la variabilidad genética de los cultivares comerciales. Los resultados obtenidos con ambos tipos de marcadores (bioquímicos y moleculares) demuestran la necesidad de combinar diferentes tipos de marcadores, pues los mismos se complementan y permiten aportar una información más completa en este sentido.
Se puede afirmar que el análisis por RAPD fue apropiado para la detección de la variabilidad genética entre especies de este género, aunque el uso de marcadores moleculares en estudios de germoplasma y diversidad genética es limitado por el costo de los reactivos y, en algunos casos, por el elevado costo de los equipos. Los avances técnicos que estas tecnologías han alcanzado en los últimos años los hacen más accesibles a los genetistas y a los programas de mejoramiento (24). Los análisis descritos en estos trabajos y los resultados obtenidos pueden complementar la caracterización morfológica y constituyen una herramienta a considerar en el diseño de los programas de mejoramiento. Estos constituyen, además, la primera información en Cuba sobre la variabilidad genética de especies dentro del género Nicotiana, empleando marcadores bioquímicos y moleculares.
AGRADECIMIENTOS
Los experimentos presentados en esta publicación se realizaron gracias al financiamiento ofrecido por la Universidad Nacional Autónoma de México (UNAM), como parte del doctorado conjunto UNAM-Universidad de La Habana.
REFERENCIAS
1. Zhang HY, Liu XZ, He CS, Zheng CM. Random amplified DNA polymorphism of Nicotiana tabacum L. cultivars. Biol Plant. 2005; 49:605-607.
2. Tapia E, Gutiérrez MA, Warburton M, Santacruz A, Villegas A. Characterization of mandarin (Citrus spp.) using morphological and AFLP markers. INCI. 2005; 30 (11):1-23.
3. Hidalgo R. Variabilidad genética y caracterización de especies vegetales. Boletín técnico No.8. Instituto Nacional de Recursos Fitogenético (IPGRI), Cali, Colombia. 2003; p. 2-26.
4. Agarwal M, Shrivastava N, Padh H. Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 2008; 27:617-631.
5. Cornide MT, Sánchez JE, Calvo D. Identificación de genotipos y progenitores. En: Marcadores Moleculares: Nuevos horizontes en la genética y la selección de las plantas. Cornide MT (Ed) Felix Varela, Habana; 2002. p. 212-220.
6. Liu XZ, Zhang HY. Advance of molecular marker application in the tobacco research. Afr J Biotechnol. 2008; 7(25):4827-4831.
7. Picca A, Helguera M, Salomón N, Carrera A. Marcadores moleculares. Capítulo 4. 2006; p.61. Disponible en: http://www.argenbio.org. (Consultado: 2 jun 2008)
8. Del Piano L, Abet M, Sorrenlino C, Acanfora F, Cozzolino E, Acanfora F, et al. Genetic variability in Nicotiana tabacum and Nicotiana species as revealed by RAPD procedure. Beitrege Zur Tab Int. 2000;19:1-15.
9.Siva-Raju K, Madhav MS, Sharma RK, Murthy TGK, Mohapatra T. Genetic polymorphism of Indian tobacco types as revealed by amplified fragment length polymorphism. Curr Sci. 2008; 94(5):633-639.
10.Siva-Raju K, Sheshumadhav M, Murthy TGK. Molecular diversity in the genus Nicotiana as revealed by randomly amplified polymorphic DNA. Physiol Mol Biol Plants. 2008;14(4):377382.
11.Dongre A, Parkhi V, Gahukar S. Characterization of cotton (Gossypium hirsutum) germplasm by ISSR, RAPD markers and agronomic values. Indian J Biotechnol. 2004; 3:288-393.
12.Chen L, Yamaguchi S. RAPD markers for discriminating tea germplasms at the inter-specie level in China. Plant Breed. 2005;124:404-409.
13.Singh AK, Singh M, Singh AK, Singh R, Kumar S, Kallo G. Genetic diversity within the genus Solanum (Solanaceae) as reveled by RAPD markers. Curr Sci. 2005; 90:711-716.
14.Valdés de la Cruz M, González C, Lara RM, Román MI, Hernández Y, Hernández RM, et al. Diversidad genética de especies silvestres del género Nicotiana I: Caracterización mediante marcadores bioquímicos. Rev. Protección Veg. 2010; 25(2):88-97.
15.Aldrich A, Cullis J. CTAB DNA extraction from plant tissues. Plant Mol Biol Rep. 1993;11(2):128-141.
16.Sambrook J, Russell DW. Molecular Cloning: A laboratory manual. Third Edition. Ed.: Cold Spring Harbor Laboratory Press, New York. U.S.A; 2001.
17.Dice LR. Measures of the amount of ecologic association between species. Ecology. 1945; (26):297-302.
18.Zhang HY, Liu XZ, He CS, Yang YM. Genetic Diversity among flue-cured tobacco cultivars based on RAPD and AFLP markers. Braz Arch Biol Technol. 2008; 51(6):1097-1101.
19.Ren N, Timko MP. AFLP analysis of genetic polymorphism and evolutionary relationships among cultivated and wild Nicotiana species. Genome. 2001;44: 559-571.
20.Del Piano L, Barbato L, Abet M, Sorrentino C, Cozzolino E, Cuciniello A, et al. ISSR analysis of genetic polymorphism among cultivated and wild Nicotiana species. In: Proceedings of the XLVlll Italian Society of Agricultural Genetics. 2004; ISBN 88-90062, 5-8, Ciudad Lecce, Italy. 15-18 Sep 2004.
21.Kanawapee N, Maensiri D, Bunnag S, Chantaranothai P. Development of species-specific SCAR markers for identification of three medicinal species of Phyllanthus. J Syst Evol. 2008; 46(4):614-621.
22.Milla R, Levin JS, Lewis RS, Rufty RC. RAPD and SCAR Markers Linked to an Introgressed Gene Conditioning Resistance to Peronospora tabacina D.B. Adam. in Tobacco. Crop Sci. 2005;(45):2346-2354.
23.Julio E, Verrrier JL, Dorlhac De Borne F. Development of SCAR markers linked to three diseases resistance based on AFLP within Nicotiana tabacum L. Theor Appl Genet. 2006;112:335-346.
24.Becerra V, Paredes CM. Uso de marcadores bioquímicos y moleculares en estudios de diversidad genética. Agric. Téc. 2000;60(3):270-281.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}