










My SciELO
 Custom services
Custom servicesServices on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Protección Vegetal
Print version ISSN 1010-2752On-line version ISSN 2224-4697
Rev. Protección Veg. vol.32 no.2 La Habana May.-Aug. 2017
ARTÍCULO ORIGINAL
Hospedantes de Lincus curvatus Campos & Roell y Macropygium reticulare Fabricius y especies vegetales reservorios de Phytomonas sp. en plantación de palma aceitera O×G
Hosts of Lincus curvatus Campos & Roell and Macropygium reticulare Fabricius and plant reservoirs of Phytomonas sp. in the O×G oil palm plantation
René Asipuela-Haro1*; Wilson Galarza2; Herman Ramírez3; María de los Ángeles Martínez Rivero4
¹ Departamento de Sanidad Vegetal, Plantación Palmar del Rio, El Coca, Ecuador.
2 Gerencia Agrícola, Plantación Palmar del Rio, El Coca, Ecuador.
3 Zamorano, Escuela Agrícola Panamericana, Asistente Sanidad Vegetal, Honduras.
4 Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, Cuba.
RESUMEN
Se estudiaron las especies vegetales circundantes a la palma aceitera que actúan como hospedantes de Lincus curvatus Campos & Roell y Macropygium reticulare Fabricius y portadoras de Phytomonas sp. en los estados de Tripomastigote y Amastigote en la provincia Orellana, Ecuador. Se estableció un experimento con un Diseño de Parcelas Divididas trifactorial, con dos bloques (Alta Mortalidad 2,5 %, Baja mortalidad 0,01 %), tres parcelas principales (Entrelínea, Bordura y Selva) y tres subparcelas, en las que se evaluó el número de insectos hospedados por unidad de especie vegetal. Se identificaron 28 especies vegetales clasificadas en 18 familias. L. curvatus se halló en representantes de las familias Fabaceae, Marantaceae, Arecaceae, Heliconiaceae, Cyclanthaceae, Asteraceae, Musaceae; mientras queM. reticulare estaba presente en especies de Urticaceae, Asteraceae, Araceae, Commelinaceae, Arecaceae, Musaceae, Poaceae, Fabaceae. Las familias reservorios de Phytomonas sp. en estado Tripomastigote fueron: Euphorbiaceae y Arecaceae; mientras que en estado Amastigote se presentó en las familias Musaceae, Araceae, Alismataceae, Poaceae, Euphorbiaceae, Heliconiaceae y Marantaceae. La mayor población de L. curvatus se encontró en la parcela Selva y M. reticulare se halló en la parcela Entrelínea.
Palabras clave: Elaeis guineensis, Elaeis oleifera, Hospedantes, Lincus, Macropygium, Marchitez sorpresiva, Pentatomidae, Phytomonas sp., Vector.
ABSTRACT
Plant hosts of Lincus curvatus Campos & Roell and Macropygium reticulare Fabricius and carriers of Phytomonas sp. surrounding the oil palm were studied in the states of Tripomastigote and Amastigote in the province of Orellana, Ecuador. It was set up a multifactorial split plot design with two blocks (High mortality-2.5 % and Low mortality-0.01 %), three plots (Inter palm lines, bordering area and jungle), and three sub-plots, in which the number of insects hosted per unit of plant species was evaluated. Twenty eight plant species classified within eighteen families were identified. L. curvatus was found in: Fabaceae, Marantaceae, Arecaceae, Heliconiaceae, Cyclanthaceae, Asteraceae, and Musaceae; M. reticulare was found in: Urticaceae, Asteraceae, Araceae, Commelinaceae, Arecaceae, Musaceae, Poaceae and Fabaceae. The families reservoirs of Phytomonas sp. in the stated of Tripomastigote were: Euphorbiaceae and Arecaceae, while in the state of Amastigote were; Musaceae, Araceae, Alismataceae, Poaceae, Euphorbiaceae, Heliconiaceae, and Marantaceae. The highest population of L. curvatus was found in the jungle plot, while M. reticulare was found in the inter palm line plot.
Key words: Elaeis guineensis, Elaeis oleifera, Hosts, Lincus, Macropygium, Pentatomidae, Phytomonas sp., Sudden wilt, Vector.
INTRODUCCIÓN
La palma aceitera (Elaeis guineensis, Jacq.) es un cultivo económicamente importante que se encuentra en varios países de Sudamérica y enfrenta un amplio grupo de enfermedades, dentro de las más letales está la Pudrición de Cogollo (PC), atribuida a hongos fitopatógenos del género Phytophthora(1) y varias especies de Fusarium (2), superada únicamente a través del mejoramiento genético (Elaeis oleifera HBK Cortés, 1897 × Elaeis guineensis Jacq.),con materiales híbridos tolerantes a esta enfermedad (3).
Este recurso genético permitió establecer nuevas plantaciones, inclusive en zonas de alta presión de la enfermedad; sin embargo, a medida que los cultivos híbridos se expanden, una nueva enfermedad letal se registra devastando estas plantaciones y comúnmente se conoce como Marchitez Sorpresiva (MS). La enfermedad se reporta en materiales Guineensis y en diferentes países de Latinoamérica: Colombia, Perú, Venezuela, Brasil y Ecuador (4). El agente causal es un protozoario parásito de plantas presente en Euphorbia hyssopifolia Linneo, Euphorbia characias Linneo, Euphorbia pinea Linneo y Manihot esculenta Crantz (5). También se aisló del látex de Euphorbia pilulifer Jacq. (6); el parásito pertenece a la Familia Trypanosomatidae, Género Phytomonas,y parte de su ciclo de vida se desarrolla en las glándulas salivales de insectos fitófagos que, a su vez, se encargan de transmitir a otras plantas (7).
Dentro de los vectores de esta enfermedad se presumen varias especies del género Lincus hallados sobre las palmas: Lincus lethiferDolling, Lincus apollo Dolling, Lincus cropius Rolston (8), Lincus tumidifrons Rolston, Lincus malevolus Rolston & Mcdonald, Lincus spurcus Rolston (9), así como Macropygium reticulare Fabricius, ya que se ha observado que se alimenta sobre las raíces de la palma (10).
De la enfermedad no hay claridad sobre su epidemiología, mucho menos en cultivos híbridos, por lo que se ha hecho necesario desarrollar esta investigación. Los objetivos del presente trabajo son identificar las plantas hospedantes del vector y analizar si son portadoras de Phytomonas.
MATERIALES Y MÉTODOS
Este estudio se desarrolló en la Plantación Palmar del Río, ubicada en la Región Amazónica del Ecuador, a una altura de 263 m.s.n.m., la precipitación media anual es de 3250 mm, con picos en los meses de diciembre a mayo y periodos ligeramente secos entre julio y septiembre; la humedad relativa promedio es de 89 % y la temperatura 25,5°C. Los conteos poblacionales de insectos y las recolecciones de especies vegetales se realizaron de enero a abril de 2014.
Diseño experimental
Para detectar las especies vegetales hospedantes y la distribución de los vectores en el área de cultivo, se planteó un Diseño de Parcelas Divididas con tres factores, se establecieron dos BLOQUES (Factor A), representados por los cultivos con mortalidades mensuales Altas 2,5% (BQ1) y Bajas 0,01% (BQ2), cada bloque fue de 225 ha y dentro se establecieron tres PARCELAS principales de 25 ha (Factor B) con tres repeticiones. Cada parcela principal fue de 75 ha llamadas por su ubicación, Entrelinea (espacios entre las palmas), Bordura (espacios entre las palmas y los caminos de circulación), Selva (Vegetación selvática que se halla a continuación de los caminos), finalmente dentro de estas tres parcelas se establecieron tres subparcelas de 360 m2 (18m x 20m), en las que se evaluaron las FAMILIAS (Factor C) de especies vegetales existentes y el número de insectos hospedados. Se realizó análisis de varianza trifactoriallas medias poblacionales se compararon mediante pruebas de mínima diferencia significativas (LSD de Fisher) con nivel de significación p ≤0,05. Se empleó el paquete estadístico Infostat 2016 (11).
Recolección e identificación de especies vegetales
La recolección de las muestras vegetales se realizaron en las subparcelas de investigación, se incluyeron únicamente las especies vegetales que superaron los 60 cm de altura. Las muestras incluyeron flores, hojas y raíces; se disecaron y se preservaron según la metodología descrita por Ferrer y Lumbreras (12). La identificación se realizó según la clave taxonómica y la metodología binaria latina expuesta por Linneo y especificadas en el Código Internacional de Nomenclatura Botánica (13).
Recolección e identificación de insectos
En un formato prediseñado para este estudio, se registró la cantidad de insectos hospedados por especie vegetal. Los insectos se recolectaron con una pinza entomológica y se dispusieron en frascos oscuros mientras se trasladaron al laboratorio para la cuantificación e identificación mediante claves taxonómicas propuesta por Campos (14).
Aislamiento de Phytomonas sp. en insectos
De la muestra general de insectos, se tomó al azar una submuestra representativa (10 %), luego se procedió a triturar cada ejemplar en forma individual siguiendo la metodología de Thomas y McCoy (15); se preparó una placa mediante un frotis y tinción con Lugol al 2 % y se observó en un microscopio a 40 y 100x.
Aislamiento de Phytomonas sp. en plantas
Luego de la recolección de las plantas, en el laboratorio se procedió a la extracción de savia, se prepararon placas y se observaron en un microscopio a 40 y 100 x. Según la presencia de flagelados, se clasificó en plantas portadoras (+) y no portadoras (-). Los flagelados observados se clasificaron en Tripomastigotes y Amastigotes, de acuerdo con la descripción fisiológica propuesta por McGhee (16).
RESULTADOS Y DISCUSIÓN
Identificación de especies vegetales
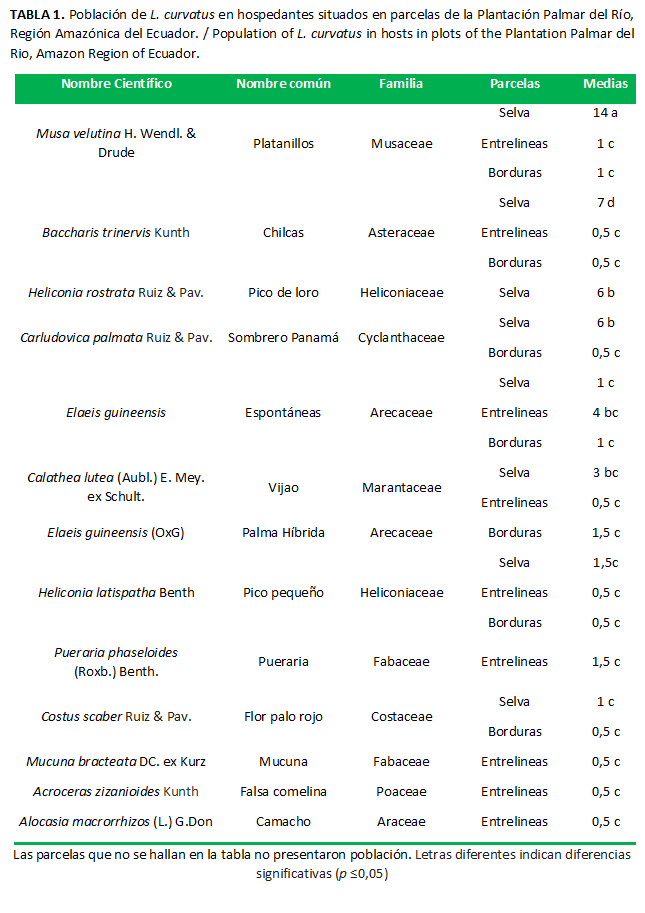
Se identificaron 28 especies vegetales pertenecientes a 18 familias, todas se encontraron circundando a los árboles de palma aceitera. Estas especies generan abundante sombra, mantienen la humedad relativa y crean un ambiente ideal para la sobrevivencia de estos pentatómidos, condición similar a lo descrito por Carbonell (17). En investigaciones desarrolladas en ocho plantaciones, referente a plantas circundantes a la palma, se identificaron 255 especies vegetales de 52 familias (18), una cantidad superior a la encontrada con relación a esta investigación; esta superioridad puede deberse a las condiciones agroclimáticas en que se hallan las plantaciones evaluadas.
Identificación de insectos hospedados
Se identificó, mediante claves taxonómicas, a L. curvatus y M. reticulare, con 109 y 157 especímenes, respectivamente.
Aislamiento de Phytomonas sp. en insectos
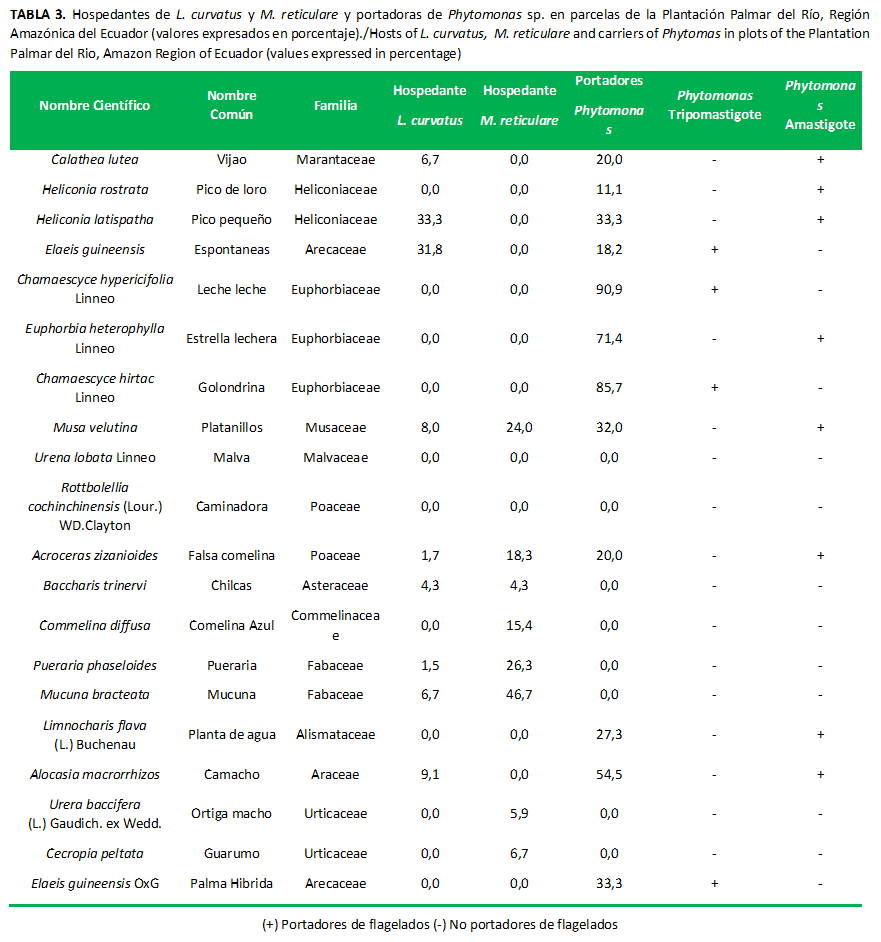
Las pruebas de tripanosomas evidenciaron 92 % y 26 % positivos para flagelados en L. curvatus y M. reticulare, respectivamente.
Aislamiento de Phytomonas sp. en plantas
De las 28 especies vegetales circundantes a la palma aceitera, se comprobó que el 43 % son potenciales portadores de flagelados; de estas en estado Tripomastigotes 33 % y en Amastigotes el 67 %; mientras que el 57 % del total de las especies evaluadas no presentaron este protozoario.
Análisis poblacional de L. curvatus
Factor A
La población promedio en los cultivos presentó diferencias significativas (p=0,0014); en el bloque de mayor tasa de mortalidad (BQ1=2,5%) se registró la mayor población de L. curvatus, lo cual coincide con lo expuesto por Dollet y Alvanil (19), que plantearon que estas poblaciones inciden en el alto índice de mortalidad. También es probable que las altas poblaciones registradas en estos bloques se deban a condiciones ambientales favorables.
Factor B
No se encontraron diferencias significativas (p=0,3621) entre las poblaciones de las PARCELAS evaluadas; sin embargo, en la parcela SELVA se registró la mayor población deL. curvatus. Las poblaciones de insectos en monocultivos se adaptan y se adentran al nuevo agroecosistema (20). Al parecer las poblaciones en forma natural viven en la vegetación selvática; sin embargo, se adaptan al nuevo agroecosistema del cultivo, migran hacia el interior y, finalmente, hacia las palmas donde ocurre la infección.
Factor C
Al comparar las poblaciones en las FAMILIAS evaluadas, existieron diferencias significativas (p=0,0063). Las mayores poblaciones de L. curvatus se hallaron hospedados en las siguientes familias: Musaceae, Asteraceae, Heliconiaceae, Cyclanthaceae, Arecaceae, Marantaceae, Fabaceae (Tabla 1). Por lo general, los pentatómidos son fitófagos, se hospedan en ambientes oscuros, húmedos y pueden sobrevivir en diferentes especies vegetales del bosque y sotobosque (21). Estas especies no se encontraron hospedadas en las partes aéreas como las hojas, las flores ni los frutos de las plantas circundantes; al contrario, se encontraron en las hojarascas, los troncos húmedos, las raíces expuestas y en ciertos tallos suculentos, además, se encontraron sobre el pedúnculo y raquis de las flores de palma aceitera; de esta forma se puede interpretar que esta especie prefiere los lugares ocultos, oscuros y húmedos.
En la interrelación Bloques × Familias, las poblaciones de L. curvatus no presentaron diferencias significativas (p=0,1244), mientras que en Parcelas × Familias se presentaron poblaciones con diferencias significativas (p=0,0365). Las mayores densidades poblacionales se hallaron en las siguientes interrelaciones; SELVA × Musaceae, SELVA × Asteraceae, SELVA × Heliconiaceae, SELVA × Cyclanthaceae, ENTRELINEAS × Arecaceae, SELVA × Marantaceae(Tabla 1). Según Altieri y Nicholls, los insectos presentan grados de adaptación en proporción con las bondades de cada agroecosistema (22); en este sentido se interpreta que esta especie está inicialmente adaptada a las condiciones de la parcela SELVA, a partir de las cuales inician un proceso de adaptación y movimiento migratorio hacia el interior del cultivo en proporción con la disponibilidad de las especies vegetales preferenciales para hospedarse y alimentarse.
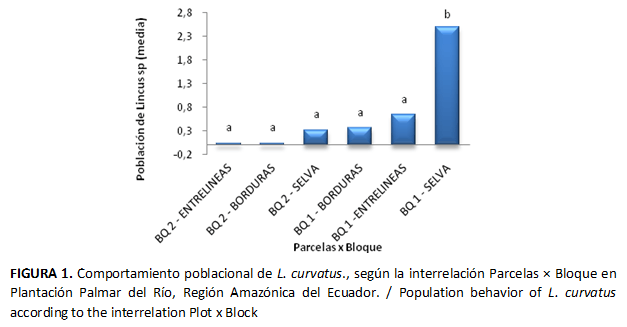
La interrelación Parcelas × Bloques presentó diferencias significativas (p=0,0370), la mayor población de Lincus curvatus se registró en la interrelación SELVA-BQ1 (Figura 1). La parcela Selva está provista de abundante vegetación, sombra y humedad; esto puede facilitar el desarrollo de altas poblaciones de vectores que se reflejan en los índices de mortalidad.
Análisis poblacional de M. reticulare
Factor A
Las poblaciones de M. reticulare, registradas en los BLOQUES, fueron diferentes estadísticamente (p=0,0255). El bloque representado por los cultivos con mayor tasa de mortalidad (2,5 %) fue donde se registró la mayor población, pudiendo interpretarse que existe una relación directa entre población y el índice de mortalidad, según Genty (23). Esta especie presenta una cantidad indeterminada de flagelados contenidos en sus glándulas salivales y tubo digestivo, lo que le convierte en un potencial vector de la enfermedad.
Factor B
El análisis estadístico demostró que no existen diferencias significativas (p=0,3048) entre las PARCELAS evaluadas; sin embargo, se describe que la parcela ENTRELINEA es donde se registró la mayor población de M. reticulare. Según Bustamante y Monroy (24), es un hecho comprobado que, dentro de su ciclo de vida, los insectos migran de acuerdo a situaciones climáticas favorables o desfavorables, de acuerdo a la temperatura, la humedad relativa y la disponibilidad de alimentos; por lo tanto, podemos explicar que los insectos hallaron condiciones favorables en las entrelíneas, lo que motiva la concentración de la población en esta parcela.
Factor C
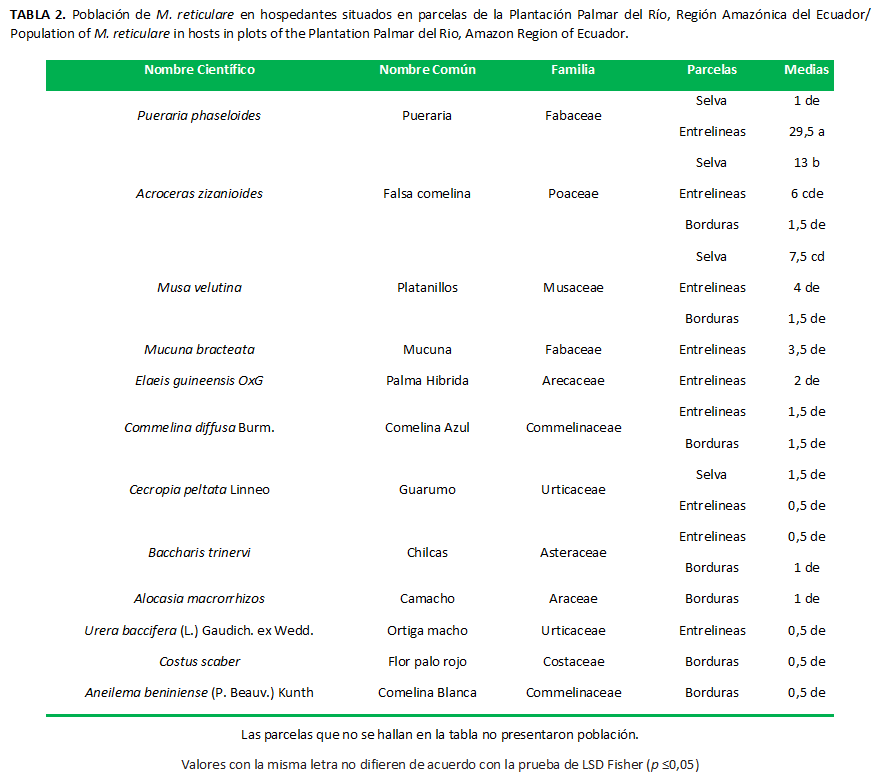
Las poblaciones de M. reticulare presentaron diferencias significativas (p=0,0025) entre las FAMILIAS evaluadas. Las mayores poblaciones de esta especie se hallaron hospedadas en las siguientes familias: Urticaceae, Asteraceae, Araceae, Commelinaceae, Arecaceae, Musaceae, Poaceae, Fabaceae (Tabla 2). Se localizaronen lugares sombríos, explorando la hojarasca húmeda y tallos suculentos, lo que coincide con lo descrito por Panizzi (25). Estas familias botánicas interactúan con el cultivo de palma, modifican el microclima del ecosistema a favor de los insectos, proveen de refugio y alimento (26) y su eliminación modificaría directamente las poblaciones.
La interrelación Bloques × Familias no reportó diferencias significativas (p=0,1257), al igual que la interrelación Parcelas × Bloques (p=0,3835). La interrelación Parcela × Familias demostró diferencias significativas (p=0,0214), las poblaciones de M. reticulare variaron según las especies vegetales (Tabla 2). Este comportamiento puede estar dado por una mayor o menor tasa de depredación y parasitismo, o un mayor o menor índice de colonización y reproducción; puede deberse a la tolerancia o intolerancia a la repelencia química, a la inhibición o disponibilidad de alimentos; o sea, a las adaptaciones y las bondades del agroecosistema (27).
Especies vegetales reservorios
Las especies vegetales, como fuente de Phytomonas en estado Ttipomastigotes, se reportaron en las familias (Tabla 3) Euphorbiaceae, Arecaceae, lo que coincide ampliamente con Rairán (28); mientras que en el estado Amastigote se encontraron en Musaceae, Araceae, Alismataceae, Poaceae, Euphorbiaceae, Heliconiaceae, Marantaceae; en ciertas familias esto coincide con lo citado por Thomas y McCoy (15). Al considerar la gran variedad de familias involucradas como reservorio de tripanosomatides, se interpreta que las especies vegetales son determinantes en la evolución de la enfermedad, debido a que los insectos pueden tomar los flagelados a partir de estas familias. El manejo de las especies vegetales circundantes a la palma reduciría el hospedaje, las poblaciones y el reservorio de los tripanosomas.
Como conclusión, podemos decir que los cultivos de palma aceitera se hallan asociadas a numerosas y variadas especies vegetales; estas, a su vez, actúan como hospedantes de los vectores de la enfermedad. Varias de estas especies también actúan como portadoras del agente causal, al igual que un grupo significativo de los insectos vectores. Desde este punto de vista, es imprescindible el control de las malezas para evitar la sobrepoblación del vector y, por ende, la infección de las palmas sanas.
Las siguientes especies no hospedaron a los vectores ni fueron reservorios del flagelado protozoario: Panicum maximun Jacq: Poaecea (Saboya), Aneilema beniniense: Commelinaceae (Comelina Blanca), Amaranthus spinosus Linneo: Amaranthaceae (Pega pega), Boehmeria nivea (L.) Gaudich: Urticaceae (Ortiga Fina), Emilia sonchifolia (L.) DC. ex Wight: Asteraceae (Emilia), Costus scaber: Costaceae (Flor palo Rojo), Piper peltatum Linneo: Piperaceae (Papel Higienico); Carludovica palmata Ruiz & Pav.: Cyclanthaceae (Sombrero de Panamá).
REFERENCIAS
1. Moreno-Chacón, Camperos-Reyes, et al. Biochemical and physiological responses of oil palm to bud rot caused by Phytophthora palmivora. Plant Physiology and Biochemistry. 2013;70:246-51.
4. Arango M, Pineda B, Martínez G, B. V. Consideraciones epidemiológicas y etiológicas de la marchitez sorpresiva en palma de aceite en la Zona Norte de Colombia. Revista Palmas. 2011;32(1):35-44.
6. Jaskowska E, Butler C, Preston G, Kelly S. Phytomonas: Trypanosomatids adapted to plant environments. PLoS Pathogens. 2015;11(1):1-17.
7. Dias A, Dos-Santos S, Lery S, Silva A, Oliveira, P B, et al. Evidence that a laminin-like insect protein mediates early events in the interaction of a Phytoparasite with its vector's salivary gland. PloS one. 2012;7(10):e48170.
9. Couturier G, Kahn F. Notes on the insect fauna on two species of Astrocaryum (Palmae, Cocoeae, Bactridinae) in Peruvian Amazonia with emphasis on potential pests of cultivated palms. Bull Inst fr Ét and. 1992;21(2):715-26.
14. Campos L, Roell T. Description of four new species of Ochlerini from South America (Hemiptera: Pentatomidae: Discocephalinae). Zootaxa. 2017;4236(3):507-520.
16. McGhee RB, WB. C. Biology and physiology of the lower Trypanosomatidae. Microbiological reviews. 1980; 44(1): 140.
17. Carbonell RM et al. Guía para el Trabajo de Campo en el Manejo Integrado de Plagas del Arroz. 2008. Inst. Inv. Arroz, CIAT; Cuba-Colombia. 82 pp.
19. Dollet M, Alvanil F, Diaz A, et al. Les pentatomides vecteurs des trypanosomes associes au hartrot du cocotier et Marchitez du palmier. Annales ANPP. 1994; 3: 65-75.
20. Fry G. Landscape ecology of insect movement in arable ecosystems: John Wiley and Sons. Bristol UK. 1996.
24. Bustamante DM, Monroy C, Rodas A. Diferenciación morfométrica de pobladores de época lluviosa y de época seca de Triatoma dimidiata (Hemiptera: Reduviidae: Triatominae), en dos áreas de Guatemala. Revista Científica. 2014;15:67-90.
25. Panizzi AR, Grazia J. Introduction to True Bugs (Heteroptera) of the Neotropics. True Bugs (Heteroptera) of the Neotropics. Springer. 2015: 3-20.
26. Montero G. Ecología de las interacciones entre malezas y artrópodos. Capítulo X. 2014: 267-305.
28. Rairán N, Mejía H, Calvache HH, Zambrano JE, Grijalva O. Distribución de las fitomonas en palma de aceite afectadas por la Marchitez sorpresiva. Revista Palmas. 2000;21(1):148-56.
Recibido: 8/9/2016
Aceptado: 21/9/2017
* Autor para correspondencia: René Asipuela-Haro. E-mail: rasipuela@palmardelrio.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}