










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista de Protección Vegetal
On-line version ISSN 2224-4697
Rev. Protección Veg. vol.32 no.2 La Habana May.-Aug. 2017
ARTÍCULO ORIGINAL
Transmisión de la enfermedad marchitez sorpresiva en palma aceitera por Lincus curvatus Campos & Roelly Macropygium reticulare Fabricius (Hemiptera: Pentatomidae)
Transmission of the sudden wilt disease in the O x G oil palm (Elaeis oleifera × Elaeis guineensis) by the bugs Lincus Curvatus Campos & Roell and Macropygium reticulare F. (Hemiptera: Pentatomidae)
René Asipuela-Haro¹*, Ángel Torres², María A. Martínez Rivero3
¹ Jefe del Departamento de Sanidad Vegetal, Plantación Palmar del Rio. El Coca, Ecuador.
² Supervisor del Departamento de Sanidad Vegetal, Plantación Palmar del Rio. El Coca, Ecuador.
3 Centro Nacional de Sanidad Agropecuaria (CENSA), Apartado 10, San José de las Lajas, Mayabeque, Cuba.
RESUMEN
En Ecuador, provincia Orellana, se estudió el comportamiento de Lincus curvatus Campos & Roell y Macropygium reticulare Fabricius como posibles vectores de la enfermedad Marchitez sorpresiva en cultivos híbridos de palma aceitera (Elaeis oleifera × Elaeis guineensis). Se determinó el sitio de hospedaje de ambos insectos, los portadores de flagelados, así como la trasmisión de la enfermedad con L. curvatus en cultivos de 26 meses y con M. reticulare en cultivos de cuatro y ocho años. Se compararon los valores mediante un análisis de comparación múltiple de proporciones empleando el software CompaproWin 2.0. Se recolectaron 1025 especímenes, donde el 76,98 % correspondió a L. curvatus y el 23,02 % a M. reticulare. De los ejemplares de L. curvatus,el 80,48 % se halló en el pedúnculo de las inflorescencias y el 19,52 % en las entrelíneas del cultivo; el 100 % de M. reticulare se halló solamente en las entrelíneas del cultivo. Se confirmó que el 83,40 % de las poblaciones de L. curvatus y el 59,75 % de M. reticulare portaban flagelados (Familia: Tripanosomatidae). Se confirmó también la trasmisión de la enfermedad mediante ambas especies. Con L. curvatus, los síntomas se reprodujeron en el 70,8 % de las plantas evaluadas con el primer caso encontrado a los 27 días de haber depositado los insectos en el pedúnculo. Con M. reticulare, los síntomas se reprodujeron en el 25,0 % en palmas de cuatro años y en el 16,67 % en cultivos de ocho años después de los 140 y 144 días, respectivamente, de haberse depositado los insectos en las raíces
Palabras clave: Elaeis guineensis, Elaeis oleifera, Insecto vector, Lincus, Macropygium, marchitez sorpresiva, Phytomonas.
ABSTRACT
The behavior of Lincus curvatus Campos & Roell and Macropygium reticulare (Fabricius) as possible vectors of the disease known as sudden wilt in the oil palm hybrid (Elaeis oleífera × Elaeis guineensis) was studied in the province of Orellana, Ecuador. The hosting site of both insects and those carrying flagellates were determined. Disease transmission with L. curvatus in 26-month-old crops and with M. reticulare in 4- and 8- year- old crops was also determined. The values within these determinations were compared by a multiple comparison analysis of proportions using the software Compapro Win 2.0. Of 1025 specimens collected, 76.98 % corresponded to L. curvatus and 23.02 % to M. reticulare. Of the specimens of L. curvatus, 80.48 % was found on the inflorescence peduncles and 19.52 % in the crop interlines; 100 % of M. reticulare was found only in the crop interlines. It was confirmed that 83. 40 % of Lincus sp. n. and 59.75 % of M. reticulare populations carried flagellates (Family: Tripanosomatidae). The disease transmission by both species was confirmed. With L. curvatus the symptoms were reproduced on 70.8 % of the plants evaluated, with the first case reported 27 days after the insects were deposited on the peduncle. With M. reticulare, the symptoms were reproduced in 25.0 % of the 4 year and in 16.67 % of the 8 year palms after 140 and 144 days, respectively, of being deposited the insects on the roots.
Key words: Elaeis oleifera, Elaeis guineensis, Vector insect, Lincus, Macropygium, Sudden wilt, Phytomonas.
INTRODUCCIÓN
En Sur y Centro América, la Marchitez sorpresiva constituye una de las enfermedades de primer orden con carácter letal en el cultivo de palma aceitera. En Ecuador, la enfermedad está afectando los materiales tradicionales Guineensis y los híbridos interespecíficos O × G (Elaeis oleifera HBK Cortés × Elaeis guineensis Jacq.) (1).
El agente causal de la enfermedad está identificado como un flagelado protozoario de la familia Tripanosomatidae: Phytomonas staheli McGhee & McGhee (2), que infecta el floema de las plantas de la familia Arecaceae, y que, en Cocos nucifera Linneo, causa la enfermedad conocida como Hartrot (3). En zonas endémicas y no endémicas, este protozoario fue aislado del tronco de palmas E. guineensis enfermas(4).
Este protozoario se aisló, por primera vez, a partir del látex de Euphorbia pilulifer Jacq por Lafont en 1909 (5); desde entonces, la familia Euphorbiaceae se consideró reservorio de Phytomonas, posteriormente, se logró aislar P. staheli a partir del látex de Euphorbia hyssopifolia L., Euphorbia characias L.y Euphorbia pinea L. (6).
Los estudios de plantas asociadas al cultivo de palma aceitera, informaron la presencia de la familia Euphorbiaceae con 12 especies (7), es decir, que los cultivos se hallan rodeados de especies vegetales reservorios de P. staheli, los que se hallan a la espera de un insecto vector para continuar su ciclo de vida heteroxenico en invertebrados y plantas o invertebrados y vertebrados (8). Se informa que el género Lincus es vector de este protozoario (9, 10) y las especies involucradas en la transmisión son: Lincus croupius Rolston, Lincus apollo Dolling, Lincus dentiger Breddin y Lincus lethifer Dolling(11). Otra especie sospechosa es Macropygium reticulare Fabricius., pero no se notifica la reproducción de los síntomas de la enfermedad, aunque se le relaciona debido a que se observaron agujereando raíces adventicias de la palma aceitera (12).
Para Ecuador y la región neotropical, se informa por primera vezsobre la existencia de Lincus curvatus Campos & Roell, recolectado en las entrelíneas y pedúnculos de palmas enfermas (13). Reproducir los síntomas de la enfermedad Marchitez sorpresiva en palmas híbridas, a través de L. curvatus. y M. reticulare, fue el objetivo del presente trabajo.
MATERIALES Y MÉTODOS
El estudio se desarrolló en la región amazónica ecuatoriana, provincia Orellana, en la plantación Palmar del Río (S0 17.169 W 77 06.102). El promedio anual de precipitación es de 3250 mm, con picos en los meses de diciembre a mayo y periodos ligeramente secos entre julio a septiembre, la humedad relativa promedio es de 89 % y la temperatura media 25,5 °C.
Sitios preferenciales de hospedaje
Para definir el sitio preferencial de hospedaje de L. curvatus y M. reticulare, se seleccionaron cultivos de 26 meses de edad, los que se clasificaron según cuatro índices de mortalidad mensual en: Inicial (0,00 a 0,0 9%), Bajo (0,10 a 0,21 %), Medio (0,22 a 0,66 %) y Alto (0,67 a 1,12 %).
En cada índice se escogieron 16 palmas enfermas, para un total de 64 palmas en las que se recolectaron los insectos y contaron los hospedados, tanto en la corona de racimos como en las plantas de las entrelíneas del cultivo, del primer sitio se extrajeron los racimos, se disectaron y se procedió con la captura (14); en el segundo sitio, se tomó como referencia la palma enferma y se exploraron las entrelíneas y se procedió con la recolección de los insectos.
Los insectos capturados se separaron por géneros y se mantuvieron vivos en envases de cristal para la siguiente fase de estudio. Se registró la cantidad de individuos por cada género y el sitio de recolección.
Población portadora de flagelados
La población de portadores de flagelados se determinó a partir de los insectos capturados. En condiciones de laboratorio, los ejemplares se trituraron de forma individual, siguiendo la metodología de Thomas y McCoy (15), se preparó una placa mediante un frotis y se procedió a la tinción con Lugol al 2 % y a la observación en un microscopio a 40x. Se registró la cantidad de insectos que dieron positivos y negativos para flagelados.
Según los índices de mortalidad, se compararon los cuatro grupos definidos en cuanto al porcentaje de población portadora y no portadora de flagelados presentes en cada sitio de hospedaje. Para el análisis se empleó la comparación múltiple de proporciones, mediante el software CompaproWin 2.0 (16).
Reproducción de la enfermedad en palmas sanas
Una nueva recolección de insectos se efectuó en palmas enfermas, con el objetivo de hospedarlos en palmas sanas para reproducir los síntomas de la enfermedad. Para garantizar la trasmisión del agente causal, de la muestra total se extrajo una submuestra equivalente al 10 % de la población total; en laboratorio se procedió con la trituración, la tinción y la observación a 40x, según la metodología de Thomas y McCoy (15).
Los insectos restantes se sometieron a pruebas de sobrevivencia, se clasificaron por sexo y se depositaron en cajas Petri; se mantuvieron en estas cajas durante siete días con agua y alimento. Los insectos muertos y enfermos se reemplazaron. Luego de la etapa de sobrevivencia, se trasladaron a campo y se dispusieron en las palmas libres de flagelados e insectos para lo cual, mediante una extracción de savia, se descartó la presencia de flagelados en las plantas seleccionadas; mediante la revisión de las inflorescencias y entrelíneas se confirmó la ausencia de estos insectos.
La reproducción de la enfermedad con L. curvatus se realizó en palmas sanas, para ello se depositaron diez insectos adultos en las inflorescencias tiernas (cinco machos y cinco hembras) y, para evitar el escape de los insectos, las inflorescencias se enfundaron con una bolsa de papel incluyendo el pedúnculo. En total se seleccionaron 24 palmas sanas y se emplearon 240 insectos adultos.
La reproducción con M. reticulare se ejecutó en palmas sanas, para lo cual se seleccionaron raíces tiernas y expuestas, las que se introdujeron en los envases plásticos a través de las perforaciones realizadas en cada envase y se depositaron diez insectos adultos; las bocas de estos envases se sellaron con las tapas correspondientes para evitar el escape de los insectos. Con esta especie se realizaron dos experimentos, donde se evaluó la tolerancia a la enfermedad según la edad de cultivo; el primero sobre ocho palmas en cultivos de cuatro años, el segundo sobre 12 palmas en cultivos de ocho años. En total se emplearon 80 y 120 insectos, respectivamente.
Durante todo el estudio, las palmas recibieron el manejo agronómico estándar y el seguimiento respectivo hasta la aparición de los primeros síntomas. Se compararon las cantidades de palmas que reprodujeron los síntomas de la enfermedad Marchitez sorpresiva mediante un análisis de comparación múltiple de proporciones, empleando el software CompaproWin 2.0 (16).
RESULTADOS Y DISCUSIÓN
En la primera recolección se capturaron 1025 insectos, de estos el 76,98 % correspondió a L. curvatus y el 23,02 % a M. reticulare.
L. curvatus
Sitios preferenciales de hospedaje
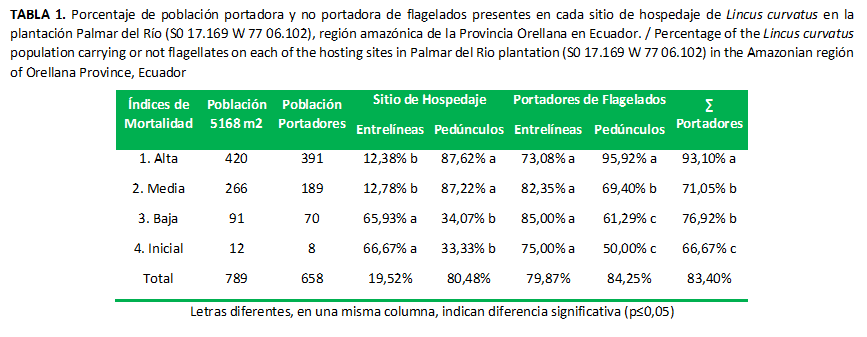
Existen diferencias significativas entre los sitios de hospedaje (Tabla 1). Los cultivos con mortalidad alta y media presentaron las mayores poblaciones de insectos hospedados en el pedúnculo, posiblemente debido a que en este órgano se aglutina la mayor concentración de nutrientes para la formación de los racimos (17) y el insecto aprovecha para alimentarse y, a la vez, infecta la planta; mientras que los cultivos con mortalidad baja e inicial presentaron las mayores poblaciones de insectos en las entrelíneas, estos sitios le ofrecen al insecto un ambiente ideal para su desarrollo, similar a las requeridas por Lincus spurcus, que prefiere bosques húmedos y sombríos para su sobrevivencia (14).
En los lotes con altos índices de mortalidad, el control del vector deberá dirigirse a las inflorescencias y racimos, a través de aplicaciones biológicas o químicas (18); mientras que en los lotes con bajos índices de mortalidad, el control debe ser dirigido a las entrelíneas del cultivo, para impedir que el vector migre al pedúnculo. Varios autores sugieren para el control de estos insectos las aplicaciones biológicas antes que las químicas (19,20) para evitar destruir los enemigos naturales del vector y de otras plagas.
Población portadora de flagelados
Las poblaciones de insectos portadores de flagelados que se recolectaron en el pedúnculo presentaron diferencias significativas, los portadores se encuentran entre el 50 % y 96 %, en proporción con el índice de mortalidad (Tabla 2), un porcentaje acorde a lo informado en otra investigación (21) . El amplio rango de portadores puede estar en dependencia de la densidad y el tipo de vegetación circundante al cultivo, las altas densidades de vegetación atraerán los insectos y es probable que algunos insectos eviten las entrelíneas y migren directamente al pedúnculo de las palmas sanas, donde desarrollan su ciclo de vida libres de flagelados (14) , mientras que los insectos que desarrollan su ciclo en las entrelíneas son más propensos a contraer flagelados, debido a la presencia de especies vegetales que se consideran reservorios naturales del flagelado Tripanosomatidae (6) .
Se registró el 83,40 % de los insectos recolectados como portadores de flagelados, los que fueron contraídos durante su etapa de vida (22) de las Laticíferas como Apocynaceae, Asclepiadaceae, además en las Euphorbiaceae, Solanaceae, Urticaceae, Cecropiaceae y Musaceae que comúnmente circundan las palmas (1).
Las poblaciones de los insectos portadores de flagelados hallados en entrelíneas no presentaron diferencias significativas; esto puede estar dado a que los insectos, en este sitio, tienen un mayor contacto con variadas especies vegetales de la familia Euphorbiaceae, consideradas reservorios de flagelados, como Chamaescyce hypericifolia L., Euphorbia heterophylla L. y Chamaescyce hirta L. (1,2); de esta manera los insectos tienen mayor probabilidad para adquirir el flagelado durante el proceso de alimentación.
M. reticulare
Sitios preferenciales de hospedaje
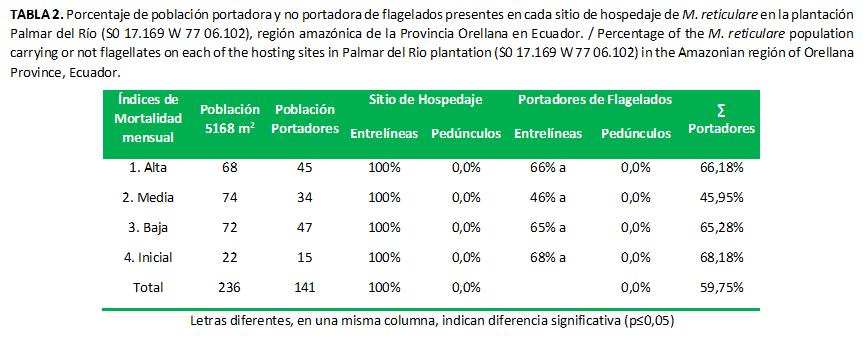
En su totalidad, este género se recolectó en las entrelíneas del cultivo. En la cobertura vegetal sembrada (Pueraria sp.) y en las malezas circundantes, no se hallaron hospedados sobre pedúnculos de palmas enfermas (Tabla 2); contrario a lo descrito por otros autores, quienes recolectaron especímenes en palmas enfermas (10) y en inflorescencias de cocoteros (23). Esto puede deberse a que cada comunidad de insectos evoluciona en dependencia de la disponibilidad de recursos en el hábitat; es decir, que la dinámica poblacional y hábitos de hospedaje es distinta en cada región (24).
Población portadora de flagelados
Las poblaciones de insectos portadores de flagelados, que se hallaron hospedados en las entrelíneas, no presentaron diferencias significativas y los índices de mortalidad no tuvieron efecto sobre los porcentajes de portadores (Tabla 2); esto puede estar dado por la escasa diversidad de especies vegetales en las entrelíneas evaluadas, o posiblemente la dieta preferencial de este género no incluye una amplia variedad de especies vegetales, por lo que la probabilidad de adquirir el flagelado es menor.
Reproducción de la enfermedad en palmas sanas
En la segunda recolección de insectos se encontró que el 92,7 % de L. curvatus y el 78,9 %de M. reticulare fueron portadores de flagelados, lo que garantizó la trasmisión de la enfermedad.
L. curvatus
Se reprodujo la enfermedad sobre cultivos híbridos; los síntomas de la enfermedad se presentaron en el 70,8 % de las 24 palmas evaluadas. La primera reproducción se observó a los 27 días y el último a los 129 días, mientras que en los reportes de otros autores, luego de enjaular insectos en palmas sanas, reprodujeron los síntomas entre los cuatro a seis meses (12).
En este estudio, los primeros síntomas se observaron en menor tiempo a lo informado por otros autores, lo que pudiera estar relacionado con la edad biológica y la resistencia genética de las palmas híbridas (25). Las condiciones climáticas también modulan los tiempos de mortalidad; es común en esta región que los casos de la enfermedad se incrementen en los meses secos y calurosos, contrario a los meses húmedos y lluviosos.
La muerte de las palmas, posterior a la manifestación de los primeros síntomas, estuvo entre los 15 a 36 días; mientras que en otra investigación se informó que ocurrió posterior a los cinco y ocho meses (21), lo que puede estar influenciado por la especie del género Phytomonas presente en cada región.
Posterior a la aparición de los síntomas, al extraer la savia de los pedúnculos florales y observarla al microscopio, se comprobó la presencia de flagelados en el 100 % de las palmas con síntomas de la enfermedad. Los síntomas fueron similares a los descritos en Guineensis, tales como pudriciones en los racimos, raíces, paquete de flechas, así como el secamiento ascendente del tercio inferior de la planta (26).
M. reticulare
En palmas de cuatro años (Tabla 3) se logró reproducir los síntomas en el 25 % de las ocho palmas evaluadas, los cuales se expresaron a partir de los 140 días hasta los 146 días; en palmas de ocho años de edad se reprodujeron los síntomas en el 16,67 % de las 12 palmas evaluadas, el primer caso se reportó a partir de los 144 días y el último a los 155 días; sin embargo, otros autores no lograron reproducir la enfermedad luego de más de un año de investigación con este género (12), así como que después de inyectar en palmas sanas suspensiones de flagelados extraídos de palmas enfermas, describen que a las 24 horas los flagelados se hallaron muertos y no se logró reproducir los síntomas (27).
Es probable que la metodología desarrollada para esta investigación sea favorable para la sobrevivencia del insecto vector, así como del flagelado. Es preciso destacar que el hospedaje fue obligado, lo que pudo influir notablemente en la transmisión del agente causal y, por ende, en la reproducción de los síntomas.
Estadísticamente se puede observar diferencias significativas entre los años evaluados; existe menor mortalidad en palmas de mayor edad, lo que pudiera estar asociada a algún tipo de tolerancia debido a la lignificación de sus tejidos (28), por donde circula el protozoario flagelado.
En el seguimiento de esta investigación se observó que, además de la reproducción de los síntomas, los insectos se alimentaron de las raíces menos lignificadas y se observaron apareamientos, posteriormente posturas y nuevas generaciones sobre las raíces.
CONCLUSIONES
Se comprueba que las poblaciones de L. curvatus y M. reticulare son potenciales portadores de flagelados, aunque este último en menor porcentaje, difiriendo sustancialmente sus hábitos de hospedaje, donde L. curvatus prefiere las inflorescencias y las entrelíneas del cultivo, mientras que M. reticulare prefiere únicamente las entrelíneas y elige las malezas circundantes a la palma.
Se demuestra que, tanto L. curvatus y M. reticulare, son vectores de la enfermedad en palmas híbridas. Las inoculaciones de flagelados mediante insectos adultos hospedados en las inflorescencias y raíces, respectivamente, permitieron reproducir la enfermedad en mayor porcentaje con el género Lincus.
REFERENCIAS
1. Alvañil Á. Marchitez sorpresiva y Hartrot: una revisión. Revista Palmas. 1996;17(1):11-8.
2. Porcel B. DF, Opperdoes., Noel B., Madoui M., Hammarton T., Field M., Da Silva C., Couloux A., Poulain J. The streamlined genome of Phytomonas spp. relative to human pathogenic kinetoplastids reveals a parasite tailored for plants. PLoS Genetics. 2014;10(2):e1004007.
3. Magalhães MM, Miguens FC, Keller DG. Requerimento de hemina no crescimento de Tripanosomatídeos floemáticos. Confict. 2013;1:25.
7. Cantuca S, Quevedo E, Peña EA, Coral OC. Reconocimiento taxonómico de plantas asociadas con la palma de aceite (Elaeis guineensis Jacq.) en plantaciones de la zona de Tumaco. Revista Palmas. 2001;22(1):27-37.
10. Liceras L, Hidalgo, JL. Lincus sp. (Hem.: Pentatomidae), agente vector de la “marchitez subita” de la palma aceitera en el Perú. Rev Peruana Entomol. 1987;30:103-4.
11. Louise C, Dollet M, Mariau D. Research into Hartrot of the coconut, a disease caused by Phytomonas (Trypanosomatidae), and into its vector Lincus sp. (Pentatomidae) in Guiana. Oléagineux. 1986;41(10):437-49.
12. Genty P. Resultados preliminares sobre el papel del Pentatomidae del género Lincus en la transmisión de la marchitez de la palma en América Latina. Revista Palmas. 1985;6(2):15-7.
15. Thomas D, McCoy R, Norris R, Espinoza A. Electron microscopy of flagellated protozoa associated with marchitez sorpresiva disease of African oil palm in Ecuador. Phytopathology. 1979;69(3):222-6.
16. Duvergel Y, Miranda I. COMPAPROP: Sistema para comparación de proporciones múltiples. Rev Protección Veg. 2015;29(3):231.
18. Asipuela-Haro R, Guevara-Poma M, Ramírez HP, Martínez Rivero MA. Evaluación de hongos entomopatógenos como controladores biológicos de Lincus sp. (Hemiptera: Pentatomidae). Rev. Protección Veg. 2015;30:94.
19. Resquín G, Garrido I, Quesada E. Combined use of entomopathogenic fungi and their extracts for the control of Spodoptera littoralis (Boisduval) (Lepidoptera: Noctuidae). Biological Control. 2016;92:101-10.
21. Di Lucca AGT, Chipana T, Fernando E, Talledo Albújar MJ, Dávila Peralta W, Montoya Piedra YC, et al. Slow wilt: another form of Marchitez in oil palm associated with trypanosomatids in Peru. Tropical Plant Pathology. 2013;38(6):522-33.
22. Dollet M, Jean J François. Myndus spp., (Fulgoromorpha, Cixiidae) public enemy number one of the coconut palm, Cocos nucifera. CIRAD - Symposia 2014(1):39-40.
26. Rojas ET. Detección y manejo de la Marchitez sorpresiva (MS) en híbrido OxG. Revista Palmas. 2013;34(4):89-93.
28. Huber D, Wilhelm NS. The role of manganese in resistance to plant diseases. Manganese in soils and plants: Springer; 1988. p. 155-73.
Recibido: 1/7/2016
Aceptado: 21/9/2017
* Autor para correspondencia: René Asipuela-Haro. E-mail: rasipuela@palmardelrio.com


{kind=link}
{kind=link}
{kind=link}