










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkVaccimonitor
On-line version ISSN 1025-0298
Vaccimonitor vol.17 no.3 Ciudad de la Habana Sept.-Dec. 2008
Síntesis intratecal de C3c e inmunoglobulinas en niños con meningoencefalitis bacteriana
Intrathecal synthesis of C3c and immunoglobulins in children with bacterial meningoencephalitis
Bárbara Padilla-Docal, Alberto Juan Dorta-Contreras, Raisa Bu Coifiu-Fanego, Elena Noris-García, Hermes Fundora-Hernández, Jesus Callol-Barroso, Marlen González-Hernández
Laboratorio Central de Líquido Cefalorraquídeo (LABCEL), Facultad de Ciencias Médicas "Dr. Miguel Enríquez", Instituto Superior de Ciencias Médicas de La Habana. Ramón Pintó 206, Luyanó. Ciudad de La Habana, Cuba.
Correo electrónico: barbara.padilla@infomed.sld.cu
RESUMEN
Las meningoencefalitis bacterianas constituyen una fuente importante de morbilidad, mortalidad y discapacidad en diferentes regiones del mundo. El objetivo del presente trabajo es conocer si el sistema de complemento puede estar involucrado en la lisis de las bacterias productoras de meningoencefalitis a través de la liberación de C3c al líquido cefalorraquídeo. Se estudiaron siete pacientes con edad promedio de 3 años, que ingresaron en el Hospital Pediátrico de San Miguel del Padrón, a los que se les realizó una punción lumbar diagnóstica y se les aislaron los gérmenes siguientes: Neisseria meningitidis, Streptococcus pneumoniae y Haemophilus influenzae. La cuantificación de los niveles de C3c, albúmina e inmunoglobulinas mayores en suero y líquido cefalorraquídeo se realizó en placas de inmunodifusión radial. Los resultados obtenidos fueron recogidos en un reibergrama. El total de los pacientes estudiados mostraron síntesis intratecal del componente C3c del sistema de complemento. Este hecho evidenció la activación de este sistema en alguna de sus vías y que una vez cumplidas sus funciones biológicas, ha sufrido un proceso de degradación y liberación al LCR en forma de C3c.
Palabras clave: Complemento, C3c, reibergrama, líquido cefalorraquídeo, Neisseria meningitidis, Streptococcus pneumoniae, Haemophilus influenzae.
ABSTRACT
Bacterial meningoencephalitis is an important source of morbidity, mortality and disabilities in different regions of the world. The objective of this paper is to know if the complement system can be involved in producing-meningoencephalitis bacterial lysis through C3c release into cerebrospinal fluid. Seven patients with an average age of 3 years-old, who attended the Pediatric Hospital of San Miguel del Padrón, were studied by lumbar puncture diagnosis. Neisseria meningitidis, Streptococcus pneumoniae and Haemophilus influenzae were isolated from the samples taken. The quantification of C3c, albumin and main immunoglobulins in serum and in cerebrospinal fluid were carried out by radial immunodifusion. Results were analyzed using a Reibergram. All patients showed C3c intrathecal synthesis. This fact demonstrates that the activation of this system has occurred in some of their three pathways and once its biological functions are fulfilled, it suffered a degradation and release process into cerebrospinal fluid as C3c.
Keywords: Complement, C3c, reibergram, cerebrospinal fluid, Neisseria meningitidis, Streptococcus pneumoniae, Haemophilus influenzae.
Introduccion
La meningitis bacteriana ha sido reconocida desde hace varios siglos como un gran síndrome en la literatura médica universal, encontrándose descripciones de la misma desde el siglo XVI. Viesseux en el año 1805 asoció esta patología a un cuadro con características epidémicas asociado a la presentación de un exantema purpúrico, mencionado como "fiebre purpúrica maligna" (meningococemia), con compromiso del sistema nervioso central (SNC). Del 75% al 80% de los agentes responsables incluyen: Neisseria meningitidis, Streptococcus pneumoniae, Haemophilus influenzae, en dependencia de las áreas geográficas.
A pesar de la actual existencia de distintos antibióticos para lograr una terapéutica eficaz, esta enfermedad puede producir un cuadro potencialmente grave. La colonización de la nasofaringe (interacción entre receptores del huésped y factores patogénicos bacterianos tales como: fimbrias, cápsula, etc, los cuales son cruciales para el establecimiento del germen a nivel de los receptores de las células epiteliales) y la posterior invasión del agente infeccioso, constituyen las primeras etapas, seguido de la invasión del hospedero y el acceso al SNC (1).
El sistema del complemento es un importante mecanismo de defensa que interactúa con los anticuerpos fijadores de complemento y es un importante mediador humoral de la inflamación. La activación de la cascada del complemento es esencial en la restricción del paso de bacterias al SNC. Esto se evidencia en aquellos individuos con déficit en algunos de los componentes de la cascada mencionada (C5, C6, C7, C8), particularmente relacionados con infecciones por N. meningitidis (2).
El complemento se define actualmente como un sistema multimolecular compuesto por más de 20 proteínas: 7 séricas, 5 reguladoras de membrana, una reguladora sérica y 8 receptores de membrana celular que se unen a los fragmentos derivados de la activación de este sistema que se caracteriza por ser una reacción enzimática en cascada en que la proteólisis de un componente activa al siguiente, produciéndose un fenómeno de amplificación (3, 4).
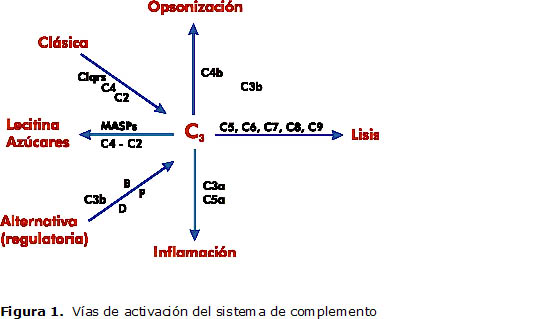
El sistema se activa a través de tres vías: la clásica, la alternativa y de la lectina, en forma diferente y convergen a nivel de C3, el cual activa el extremo final de la cascada, promoviendo la inflamación, eliminación de patógenos y facilitando la respuesta inmune (5) (Figura 1).
Del componente C3 se deriva la principal opsonina, que es, además, la molécula central más importante del sistema de complemento. Es activada por las tres vías y a su vez los productos de su activación median la opsonización, actividad anafiláctica y la activación de los compuestos efectores finales del complemento. El déficit de C3 se manifiesta por infecciones piógenas severas, causadas principalmente por organismos encapsulados (5, 6).
El objetivo del presente trabajo es conocer si el sistema de complemento puede estar involucrado en la lisis de las bacterias productoras de meningoencefalitis a través de la liberación de C3c al LCR.
Materiales y Métodos
Se colectaron muestras de suero y LCR de 7 niños con un promedio de 3 años de edad, con diagnóstico de meningoencefalitis bacterianas por Neisseria meningitidis serogrupo B en 5 pacientes y Streptococcus pneumoniae tipo b y Haemophilus influenzae en un paciente cada uno, durante los años 2006 y 2007, que ingresaron en el Hospital Pediátrico de San Miguel del Padrón, Ciudad de La Habana. Estas muestras fueron tomadas de rutina en los ingresos con signos sugestivos de meningoencefalitis.
Las muestras fueron colectadas en el momento del ingreso, coincidiendo con el inicio de los signos/síntomas y fueron conservadas a -80 oC hasta el momento de su uso. Se cuantificaron los niveles de C3c, IgG, IgA e IgM (inmunoglobulinas mayores) en suero en placas de inmunodifusión radial NOR Partigen y en LCR en placas LC Partigen (Dade-Behring, Marburg).
Para poder conocer la funcionalidad de la barrera sangre-LCR se procedió a cuantificar la albúmina en suero en placas de inmunodifusión radial NOR Partigen y en LCR en placas LC Partigen (Dade Behring, Marburg).
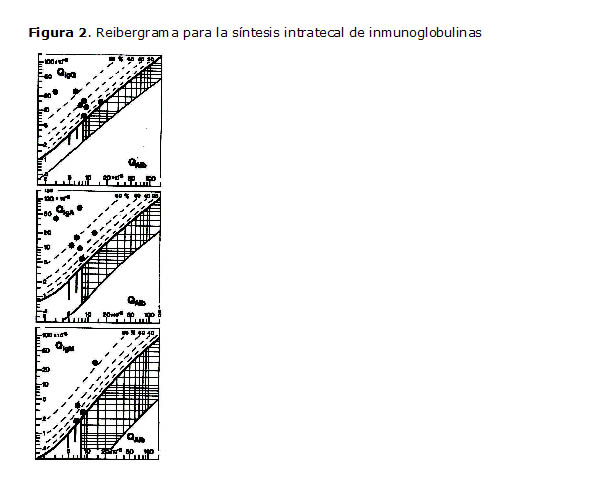
Una vez obtenidos los resultados se procedió a colocar los mismos en un reibergrama diseñado para C3c (7). Los de las inmunoglobulinas mayores también fueron colocados en los reibergramas correspondientes (8).
Los reibergramas o gráficos de las razones de Reiber son diagramas en los que básicamente se analiza de forma integrada la funcionalidad de la barrera sangre-LCR y la síntesis intratecal de las inmunoglobulinas mayores, y más recientemente ha sido utilizado para la síntesis de IgE y el C3c (7, 9), que de forma aislada no lograrían tener un mayor impacto en el diagnóstico de algunas enfermedades asociadas a determinados patrones.
Los analitos que deben cuantificarse para confeccionar un reibergrama son: albúmina, IgA, IgM, IgG, IgE y C3c, tanto en LCR como en suero. La razón LCR/suero (Q) es el cociente obtenido de la división entre la concentración del analito en el LCR y en el suero.
La curva hiperbólica más fuerte representa la línea de descripción (Q límite) entre la fracción de inmunoglobulina derivada del SNC y la fracción de la sangre. Valores ubicados por encima de esas líneas discriminatorias representan síntesis intratecal de IgG, IgA, IgM u otro componente a analizar. Las curvas de puntos indican la fracción intratecal en porcentaje con respecto a la concentración total de la inmunoglobulina en el LCR (10, 11, 12).
El límite de rango de referencia para Q Alb entre la concentración normal e incrementada de esta proteína junto con un reducido entorno (disfunción de la barrera sangre-LCR) se indica a través de las barras verticales. Esta razón albúmina depende de la edad, es decir, que a medida que aumenta la edad se produce una permeabilidad diferente que resulta algo mayor. Para su señalización hay tres barras verticales más pronunciadas: la primera (de izquierda a derecha) señala el límite hasta los 15 años (Q Alb=5x10³), la segunda hasta los 40 (Q Alb=6,5x10³) y la tercera hasta los 60 (Q Alb=8x10³). Estos datos pueden utilizarse para el uso del reibergrama en los departamentos o consultas. Sólo se tendría que plotear la Q Alb y la Q inmunoglobulina del paciente a partir de las concentraciones de albúmina, IgA, IgM e IgG en suero y LCR (10, 11,12).
Resultados y Discusión
En la Figura 2 se observa el gráfico de las razones de Reiber para la evaluación de la síntesis intratecal de inmunoglobulinas mayores. Obsérvese que todos los pacientes mostraron síntesis intratecal de IgG e IgA. Con respecto a la IgM, 4 de los pacientes mostraron síntesis de esta inmunoglobulina.
En la Figura 3 se observa el reibergrama para la evaluación de la síntesis intratecal de C3c. Se aprecia que todos los pacientes estudiados mostraron síntesis de este componente.
Este diagrama, además, nos permite conocer el estado de la barrera hematoencefálica a través de la razón albúmina (Q Alb); nuestros pacientes son niños, por lo que su valor normal de Q Alb debe ser menor o igual a 5, por tanto, de los 7 pacientes estudiados, 6 mostraron disfunción de la barrera sangre -LCR, para un 85,7%.
Las muestras utilizadas en este estudio pertenecen a pacientes con diagnóstico de meningoencefalitis por Neisseria meningitidis, Streptococcus pneumoniae y Haemophilus influenzae, provenientes del Hospital Pediátrico de San Miguel del Padrón, localizado en Ciudad de la Habana. Estas muestras fueron recibidas en nuestro laboratorio desde el mes de noviembre del 2006.
Para comprender los mecanismos a través de los cuales el sistema inmunológico participa en la defensa del hospedero contra estos patógenos y cómo estos son capaces de evadir dichos mecanismos y producir enfermedad, se hace necesario recurrir a los factores de virulencia de los mismos. Entre estos se encuentran:
" La secreción de la IgA proteasa, la que rompe la IgA secretora y sérica e inactiva el sistema de defensa del hospedero. Este mecanismo facilita la adherencia, colonización y la subsiguiente infección de mucosas, por lo que desempeña un papel importante en la patogénesis (13).
" La presencia de una cápsula que protege al microorganismo de la fagocitosis, de la desecación y de la acción de algunas lisozimas. Además, lo protege contra la lisis, lo que le otorga mayor posibilidad de supervivencia en la circulación sanguínea. Esta cápsula está compuesta por un polisacárido y su estructura le confiere la propiedad de inactivar al complemento e interferir la opsonización, evitando la fagocitosis y la lisis mediada por el complemento (14-16).
" Presencia de lipooligosacáridos, los cuales inducen una cascada de citocinas que incluye el factor de necrosis tumoral alfa e interleucina 1 y 6, que activan la vía alterna del complemento, alteran la permeabilidad de la barrera hematoencefálica y provocan la activación policlonal de los linfocitos B (14).
" Las proteínas de membrana externa son componentes estructurales de la misma con importantes funciones para mantener con vida a dichas bacterias (17).
" Las fimbrias o pili son estructuras filamentosas de naturaleza proteica ubicadas en la superficie celular, cuya función es facilitar la adherencia a la mucosa bucal o nasofaríngea y al endotelio vascular (18, 19).
La primera línea de defensa del hospedero y la más importante contra estos patógenos es la integridad de las mucosas; una vez que atraviesa esta membrana y alcanza el torrente sanguíneo, la defensa principal es el sistema de complemento y la presencia de anticuerpos específicos. La vía clásica requiere la formación de complejos antígeno-anticuerpos (Ag-Ac) para activar el factor C1 y así sucesivamente a toda la cascada del complemento, culminado en la lisis bacteriana (20). Cuando no existen Anticuerpos específicos, se activa la vía alterna a partir del factor C3 y la capacidad de estos patógenos para evadirla le permite sobrevivir en la circulación sanguínea. El ácido siálico facilita la unión del factor H (proteína reguladora del complemento) al factor C3, impidiendo así la activación subsiguiente de toda la cascada (21). Por otra parte, la presencia de anticuerpos y del sistema de complemento desempeñan un papel importante en la defensa del hospedero, provocando la lisis bacteriana, la fagocitosis por monocitos o polimorfonucleares o neutralizando los efectos mediados por la endotoxina (22).
Como se ha planteado anteriormente, el complemento es un sistema de proteínas séricas que intervienen en la defensa inmunológica del hospedero. La interacción de una bacteria con el suero de un sujeto normal desencadena la activación en cascada del complemento, bien por la vía clásica a través de los componentes C1, C2 y C4 o bien por la vía alternativa, mediante el factor D, C3 y factor B, produciéndose la lisis y activación de C3 y la subsecuente activación de los componentes finales del complemento C5-9. El fragmento C3b también se une a un producto derivado del factor B (llamado Bb) para formar el C3bBb, conocido como convertasa del C3 de la vía alternativa. Asimismo, la activación de la vía clásica produce la escisión de C4 y C2, dando lugar al complejo C4b2a o convertasa de la vía clásica. Tanto una como otra intervienen en la activación de C3 con la posterior formación del complejo de ataque a la membrana C5-9, que se inserta en las membranas celulares y produce la lisis osmótica de la célula. El déficit de los componentes del sistema del complemento conlleva a la incapacidad de producir la lisis bacteriana, por lo que la fagocitosis leucocitaria está alterada (23, 24).
El déficit de la vía alternativa o vía final común está asociado de manera especial a infecciones (meningitis y neumonía) debido a N. Meningitidis (23), el déficit de la vía clásica generalmente se asocia a enfermedades autoinmunes (25) y el de los componentes iniciales del complemento está asociado con infecciones causadas por Streptococcus pneumoniae, Haemophilus influenzae y N. meningitidis (26).
Durante una meningitis la concentración de complemento en el LCR aumenta ligeramente, pero no lo suficiente como para poder producir una opsonización de las bacterias encapsuladas que, como es sabido, es un proceso previo a la fagocitosis. Por tanto, la actividad bactericida en el LCR es mínima. La concentración de inmunoglobulinas es también muy baja. El aumento de inmunoglobulinas que se produce en el LCR en las meningitis es también escaso en comparación con el que se produce en el suero al mismo tiempo. En algunas meningitis se produce una síntesis intratecal de inmunoglobulinas que pueden ser críticas en su defensa. Una de las características típicas de las meningitis bacterianas es la pleocitosis neutrofílica que puede llegar a convertir el LCR en verdadero pus. Sin embargo, esta pleocitosis es ineficaz como mecanismo defensivo al carecer de la suficiente actividad de opsonización y bactericida (22, 23).
En todos los pacientes estudiados ha ocurrido síntesis intratecal y liberación al LCR de C3c. La síntesis intratecal de C3c y su liberación al LCR significa, en primer lugar, que ha sucedido la activación del sistema de complemento en alguna de las tres vías en que interviene y, además, que se le ha dado un uso biológico a esta proteína y luego que esta ya ha cumplido sus funciones biológicas, ha sufrido un proceso de degradación y liberación al LCR en forma de C3c.
Estos resultados nos permiten profundizar en los mecanismos involucrados en la inmunopatogenia de la meningoencefalitis bacteriana, entidad que tiene la capacidad de evadir los mecanismos de defensa del organismo debido a sus componentes superficiales que desvían o modulan el sistema inmune.
Referencias
1. Dougherty JM, Roth RM. Cerebral spinal fluid: Emerg Med Clin North Am 1986;4:281-297.
2. Conly JM, Ronald AR. Cerebrospinal fluid as a diagnostic body fluid: Am J Med 1983;75:102-107.
3. Abbas AK. Disease of Inmunology. In: Kumar, Robbins and Cotran. Pathologic Basic of diseases. 7 th edition. Saunders 2005;192-26.
4. Wen l, Atkinson J P. Clinical and laboratory evaluation of complement deficiency. JACI 2004;113:585-93.
5. Behrman r. Disorders of the complement system. En: Nelson. Textbook of Pediatrics, Philadelphia 2000;16:628-34.
6. Perkins SJ, Sim RB. Molecular modelling of human complement component C3 and its fragments by solution scattering. Eur J Biochem 1986;157:155-168.
7. Dorta-Contreras AJ, Noris-García E, Padilla-Docal B, Rodríguez-Rey A, Bu-Coifiu Fanego R, Magraner-Tarrau ME, et al. Reibergrama para la evaluación de la síntesis intratecal de C3c. Arq Neuropsiquiatr 2006; 64 (3-A):585-8.
8. Dorta-Contreras AJ. Reibergramas: elemento esencial en el análisis inmunológico del líquido cefalorraquídeo. Rev Neurol 1999; 28:996-8.
9. Dorta-Contreras AJ, Reiber H, Noris-García E. Reibergrama para la evaluación de la síntesis intratecal de IgE. Rev Neurol 2004;39: 794-5[ STANDARDIZEDENDPARAG]
10. Schuller E, Sagar H. Central Nervous system IgG synthesis in multiple sclerosis. Application of a new formula. Acta Neurol Scand 1983;67:365-77.
11. Reiber H. Flow rate of cerebrospinal fluid (CSF) -a concept common to normal blood- CSF barrier function and to dysfunction in neurological diseases. J Neurol Sci 1994;122:189-203.
12. Reiber H. The hyperbolic function: a mathematical solution of the protein flux/CSF flow model for blood-CSF barrrier function. J. Neurol Sci 1994;126:243-245.
13. Van Alphen L. The molecular epidemiology of Haemophilus influenzae. Rev Med Microbiol 1993; 4:159-66.
14. Geme ST, Cuther D Influence of pili, fibrils, and capsule on in vitro adherence by Haemophilus influenzae. Mol Microbiol 1996; 21(1):21-31.
15. Van Deuren M, Brandtzaeg P, ven der Meer J. Update of meningococcal disease with emphasis on pathogenesis and clinical management. Clin Microbiol Rev 2000;13:144-66.
16. Bethell D, Pollard AJ. Meningococcal vaccines. Expert Rev Vaccines 2002;1:75-84.
17. Van Alphen L, Riemens T, Zanene HC. Antibody response against outer membranecomponents of Haemophilus influenzae type b strains in patients with meningitis. FEMS Microbiology letters 1983;18:189-95.
18. Clemans DL, Marrs CF, Bauer RJ, Patel M, Gilsdorf JR. Analysis of pilus ashesins from Haemophilus influenzae biotype IV strains. Infect Immun 2001; 69(11):7010-19.
19. Pollard AJ, Frasch C. Development of natural immunity to Neisseria meningitidis. Vaccine 2001; 19:1327-46.
20. Van Deuren M, Brandtzaeg P, ven der Meer J. Update of meningococcal disease with emphasis on pathogenesis and clinical management. Clin Microbiol Rev 2000;13:144-66.
21. Goldscheneider I, Gotschlich EC, Artenstein MS. Human immunity to the meningococcus. The role of humoral antibodies. J Exp Med 1969;129:1307-26.
22. Haynes B, Fauci A. Harrison TR. Introducción al sistema inmunitario. Madrid: McGraw-Hill; 1998. p. 2011-2.
23. Cuadrado Gómez LM, Palau Beato E, Pérez Venegas JJ, Gutiérrez Marcos F. Meningitis meningocócica en un paciente con deficiencia del factor 8 del complemento. Rev Clin Esp 1988;182:287-8.
24. Fijen CA, Kuijper EJ, Hannema AJ, Sjoholm AG, van Putten JP. Complement deficiencies in patients over ten years old with meningococcal disease due to uncommon serogroups. Lancet 1989;2(8663):585-8.
25. Ross SC, Densen P. Complement Deficiency States and infection: Epidemiolgy, pathogenesis and consequences of neisserial and other infections in an immune deficiency. Medicine (Baltimore). 1984; 63:243-73. [Medline].
26. Overturf GD. Related Articles, Indications for the immunological evaluation of patients with meningitis. Clin Infect Dis. 2003;36:189-94. [Medline].
Recibido: Septiembre de 2008
Aceptado: Noviembre de 2008


{kind=link}
{kind=link}
{kind=link}