










My SciELO
 Custom services
Custom servicesServices on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkBiotecnología Aplicada
On-line version ISSN 1027-2852
Biotecnol Apl vol.26 no.2 La Habana Apr.-June 2009
REVIEW
The angiogenic process and cancer
El proceso angiogénico y el cáncer
Omar López 1, Yack Magariño 2, René Delgado 3
1Centro de Estudio para las Investigaciones y Evaluaciones Biológicas, IFAL, UH. Ave. 23 #21425 entre 214 y 222, La Coronela, La Lisa, Ciudad de La Habana, Cuba
2Centro de Química Biomolecular, CQB. Calle 200, Esq. 21, Playa, Ciudad de La Habana, Cuba
3Centro Nacional Coordinador de Ensayos Clínicos, CENCEC. Calle 200, Esq. 21, Atabey, Playa, Ciudad de La Habana, Cuba
ABSTRACT
Angiogenesis, the growth of new capillaries from pre-existing vessels, is closely related to processes essential for the organism, which are entirely physiological, for example embryogenesis, reproductive cycle, and wound healing, and is also associated with pathological conditions such as tumor progression, metastasis, diabetic retinopathy, hemangioma, arthritis, psoriasis and atherosclerosis, among other chronic diseases. The development of specific anti-angiogenic agents arises as an attractive therapeutic approach to treat cancer and other angiogenesis-dependent diseases. Many of these agents are being evaluated in clinical trials and have shown promising antitumor activity. This review attempts to provide a comprehensive overview of key knowledge accumulated on angiogenesis and its role in cancer, including the components of signal transduction pathways that have been explored in this process. Additionally, this review focuses on the current approaches for the discovery of new compounds that inhibit angiogenesis, emphasizing on the clinical developmental status of antiangiogenic drugs.
Keywords: Angiogenesis, cancer, tumor.
RESUMEN
La angiogénesis, el crecimiento de nuevos capilares a partir de los vasos pre-existentes, se encuentra estrechamente relacionado con procesos esenciales para el organismo, que son puramente fisiológicos; por ejemplo la embriogénesis, el ciclo reproductivo y la cicatrización de heridas. También se encuentra asociado a condiciones patológicas como el desarrollo de tumores, metástasis, retinopatía diabética, hemangioma, artritis, psoriasis y arterosclerosis, entre otras enfermedades crónicas. El desarrollo de compuestos anti-angiogénicos específicos se plantea como un atractivo enfoque terapéutico para el tratamiento del cáncer y otras enfermedades dependientes de la angiogénesis. Muchos de estos agentes están siendo evaluados en ensayos clínicos y han mostrado actividad antitumoral prometedora. Esta revisión pretende dar una visión general de los principales conocimientos acumulados acerca de la angiogénesis y su papel en el cáncer, incluyendo también los componentes de las vías de transducción de señales que se han investigado de este proceso. Además esta revisión se centra en los enfoques actuales para el descubrimiento de nuevos compuestos que inhiben la angiogénesis, con énfasis en el estado de desarrollo clínico del producto antiangiogénico.
Palabras clave: Angiogénesis, cáncer, tumor.
INTRODUCTION
The establishment and maintenance of a vascular network is an essential requirement in growth of healthy and neoplasic tissues. The cardiovascular system is the first organ system to be developed and start functioning during embryogenesis (1). Formation of new blood vessels involves two processes: vasculogenesis and angiogenesis; so it is very important to comprehend their respective conceptual differences. Vasculogenesis is the primary in situ differentiation of endothelial cells (EC) from their mesodermic precursors and its subsequent structuring in a primary capillary plexus (2). This is a highly regulated process under physiological conditions which is activated during a short period of time until inhibition, for example embryogenesis (3). On the other hand, angiogenesis is defined as the formation of new blood vessels from the pre-existent vascular layer. This event occurs during the embryonary development, also in postnatal life of the organism (4).
Angiogenesis has to be inhibited or absent in adult tissues under normal conditions. It only takes place during wound healing and in the female reproductive cycle (5). These processes are associated to the so-called: physiological angiogenesis and are produced under strict functional control mechanisms to achieve a proper balance between the positive and negative regulators (stimulators and inhibitors of angiogenesis, respectively) (6).
However, many diseases are related to the pathological angiogenesis. It has been described as one of the particular properties of cancer, playing a key role for tumor development, invasion and metastases. This only occurs in response to a stimuli or damage, breaking the balance between the regulators; and leading to an overproduction of stimulators and/or a suppression of angiogenic inhibitors (6). When the resting EC become activated by a pro-angiogenic signal, they release degradative enzymes that allow their migration, proliferation and final differentiation to form new blood vessels. Almost every field of medicine has to deal, in some way or another, with angiogenesis, associated to physiopathological processes. All the organ systems without exception are related to many diseases having angiogenesis as an essential component (5). The present work is a review about angiogenesis as a process involved in oncological pathologies, also a comment on the development and use of angiogenesis inhibitors to fight cancer.
BACKGROUND
Two important moments occurred in almost two centuries on the currently approved concept of angiogenesis as a key process for cancer development. The first one in 1887, when the British surgeon John Hunter described the process of angiogenesis as a formation of new blood vessels, which was continued during the XIX century by studying the details of the vascular anatomy of tumors. The second occurred between 1960s and 1970s decades, while beginning the research about angiogenesis related to tumor growth, that is, when Dr. Judah Folkman stated in 1971, the imaginative hypothesis that tumors depend on the angiogenic processes in growth (7). However, it was not until 1990 that angiogenesis became central for studying cancer (6), with massive uprising of antiangiogenic products that are being evaluated in clinical trials.
Research is being currently focused on obtaining compounds to modify the biology of the vascular endothelium to prevent tumor growth. These include: inhibiting endothelial proliferation, directly or by blocking the receptors of the growth factors associated to the vascular endothelium < negative regulation of the expression of these growth factors (8); and prevention of EC migration by inhibiting the activity of matrix metalloproteinases (MMP).
CHARACTERIZING THE ANGIOGENIC PROCESS
Angiogenesis is a stimuli-induced process with hypoxia as a main player (9), but it could be also induced by metabolic stimuli, as hypoglycemia and acidosis (10). It is known that events such as: local vasodilation, increased vascular permeability and accumulation of extravascular fibrin occur in response to an angiogenic stimulus (11).
The process of tumor angiogenesis is a well structured process, following a series of events, as depicted in figure 1, which are enumerated as follows:
1. Injured or ill tissues produce proteins named growth factors, which diffuse through the tissues towards the neighboring blood vessels.
2. These factors bind their specific receptors in the surface of EC on those blood vessels.
3. After this binding, the EC become activated. Signals are transduced from the cell surface into the nucleus, and the synthesis machinery of the EC begins to produce new molecules, including enzymes as the MMP.
4. The enzymes dissolve the surrounding basal membrane of the vessel, opening wholes on it.
5. The EC begin to divide (proliferate), migrating out of the vessel through the wholes in the basal membrane following the stimuli gradient.
6. At this point of the process, specialized molecules called adhesion molecules help to organize this migration by allowing the interaction between cells, also between these cells and the matrix.
7. The MMP are produced to dissolve the extracellular matrix in the leading edge of the growing blood vessel. The surrounding matrix also remodels as the vessel enlarges.
8. The EC are elongated and begin to adopt the tubular form of a blood vessel.
9. The nearest new blood vessels became connected to form a link allowing blood circulation.
10. Finally, the newly formed tubular blood vessel stabilizes by appearing smooth muscular cells and pericytes that bring structural support. Then, blood starts to flow (12).
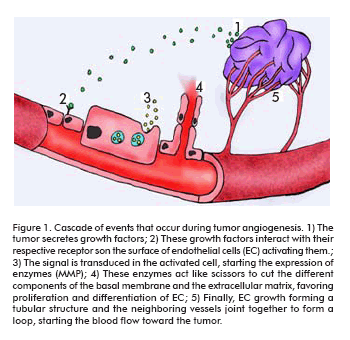
This cascade of events is divided into an activation phase and a resolution phase. The first one comprises the initiation and progression by degradation of the basal membrane, cellular migration invasion to the extracellular matrix, EC proliferation and formation of capillary lumen. Resolution comprises termination and maturation of blood vessels, EC proliferation, the end of cellular migration, the basal membrane becoming reconstituted and maturating the complex endothelial junctions (11).
RELATIONSHIP AMONG THE DIFFERENT ANGIOGENIC MEDIATORS
Angiogenesis, as any other biological process, involves many cellular components and aspects. Taking care of all the previous considerations, it depends on the control of EC functions, such as proliferation, migration and interactions with the extracellular matrix (13).
The EC usually exist in an inactive state into the vasculature, regulating the flux of nutrients of different biologically active molecules and also blood cell function. This role, as maintenance barrier of the endothelium, is exerted through receptors anchored in the endothelial membrane for many molecules, such as: growth factors, metabolites like nitric oxide and serotonin, and through specific junction molecules or other governing the interaction between these cells and the matrix (14).
EC activation can be positively or negatively regulated by several molecules, including the growth factors themselves and cytokine-related peptides, which are synthesized in normal or malignant cells (11). Moreover, they include the best positive angiogenesis regulators known so far as the vascular endothelium growth factor (VEGF) (15), and the basic fibroblast growth factor (bFGF) (16). The effect of these factors opposes other cytokines ones, such as: the transforming growth factor beta (TGF-β) (17) and the tumor necrosis factor alpha (TNF-α) (18). Both are polypeptides acting as pro-angiogenic factors in vivo; however, they do not favor endothelial growth in vitro. This has supported the hypothesis of these cytokines promoting angiogenesis indirectly, by inducing the production of positive regulators by stromal and chemo-attracted inflammatory cells (19), and tumor cells. Other cytokines have been described as angiogenesis regulators in vivo, such as: interleukin-1 (IL-1) (20), the hepatocyte growth factor (HGF) (21), the epidermal growth factor (EGF) (22) and the platelet-derived growth factor (PDGF) (23), among others. Many of the interactions of EC with the extracellular matrix are favored by extracellular proteolytic events that depend on a set of proteases and their inhibitors, produced by activated EC and other cell types (tumor cells, macrophages, etc.) (24, 25). Among them, MMP play a key role for migration of normal or tumor cells throughout the organism (26). The MMP comprise a family of at least of 16 Zn2+-dependent endopeptidases that function extracellularly and are constitutively expressed (12, 25).
These are soluble enzymes that are secreted to the extracellular space, except the membrane-associated MMP (27) which bear a transmembrane domain in the carboxyl end and are subsequently localized on the cellular surface (28). The MMP act as zymogens (the regulatory domain should be dissociated away from the catalytic domain for the enzyme to be activated). This dissociation is autocatalytic or due to the action of several enzymes such as: furin, plasmin or any other membrane-associated MMP, occurring on the surface of many cell types (29).
Proteolytic degradation of the extracellular matrix requires a strict regulation. It starts at transcriptional level by specific activated transcriptional complexes bind to regulatory elements in the genes coding for those proteolytic enzymes; also regulated by the balance of zymogens and their catalytically active counterparts; and by the binding of the activated enzymes and the zymogens to the tissular inhibitors of metalloproteinases (TIMP) (30, 31).
Once activated, the MMP can be inactivated by the above mentioned inhibitors and by the binding of plasmatic proteins as microglobulin-α2. The total balance of expression and activation of MMP is favored against the levels of TIMP and inhibition (29).
The cellular invasion, proliferation and migration processes not only depend on the enzymatic system, growth factors and their receptors, but also on the adhesion molecules involved (32). They are classified in four families according to their structure and biochemical properties: integrins, the immunoglobulin superfamily, cadherins and selectins (33, 34).
For example, during invasion and migration, integrins mediate the interaction between EC and the extracellular matrix. Similarly, during the final stage of the angiogenic process (i.e., the formation of capillary links) they play an essential role in cell-to-cell and cell-matrix interactions (35).
ANGIOGENESIS-ASSOCIATED PATHOLOGIES
Some pathologies (such as ischemic tissue damage (36) or cardiac deficiency (37)) are associated to a low angiogenic activity, but generally neovascularization processes are associated to pathologies involving high angiogenic activity, such as: chronic inflammatory processes, rheumatoid arthritis (38), Crohn´s syndrome (inflammatory bowel disease)(39), diabetic retinopathy (40), hemangioma (41), psoriasis (42, 43), endometriosis (proliferation of endometrial tissue outside of the uterus) (44), and mainly cancer (either solid or hematological tumors)(45). More recently, angiogenesis has also been involved in other disorders of varied origin such as: arteriosclerosis (46), obesity, asthma, infectious diseases, neurodegeneration and hypertension (47).
ANGIOGENESIS AND CANCER
Tumors begin to grow as small aggregates of neoplasic cells outgrowing the basal layer, without any significant input of blood. Therefore, these aggregates are avascular (48). This pre-vascular form of the tumor is denominated in situ carcinoma and is highly dependent on the proximal blood vessels for dioxygen and nutrients supply. The size of the carcinoma remains steady, without significant increase over time and limited by the balance of cellular proliferation and apoptosis/cell death (49).Without vasculature, the tumor cannot grow beyond 1-2 mm long, because it only takes the oxygen and nutrients by diffusion. The oxygen only diffuses up to 100 mm between the capillary and the cell, only covering 3 to 5 lines of cells around the capillary (48).
Once the tumor is connected to circulation by new blood vessels, it can grow and tumor cells disseminated throughout the body, metastasing (50). This demonstration generated the novel concept that cancer can be therapeutically treated by antagonizing the angiogenic process. The timing recommended for the antiangiogenic treatment is that sufficient to guarantee the affluence of blood vessels to the tumor for adequate chemotherapy, but avoiding an abundant blood supply promoting metastasis, as represented in figure 2.
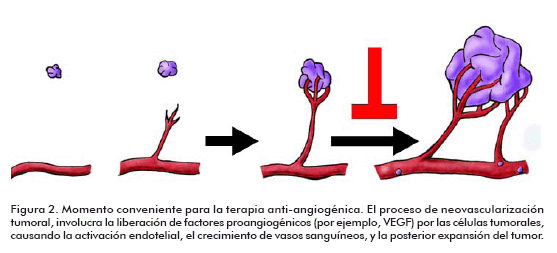
The process of tumor neovascularization comprises the release of pro-angiogenic factors (e.g., VEGF) by tumor cells, the growth of blood vessels and further expansion of the tumor.
The main triggering cause of angiogenesis toward the tumor is the hypoxic damage due to the increased distance between tumor cells and capillaries. Hypoxia induces the expression of VEGF and its receptor through the hypoxia inducible factor 1a (HIF-1α) (51), which is also a factor attracting macrophages (52-54). Over time, some carcinoma cells can gain the capacity to express certain genes coding for natural angiogenic grow factors. On the other hand, some pro-inflammatory cells (mainly lymphocytes and macrophages) (55) are attracted by the tumor, also secreting grow factors (56).
The capacity of tumors to induce and sustain the angiogenic process is attributed to two main changes. The first is a process of loss of function, by decreasing or negatively regulation of endogenous inhibitory proteins such as trombospondin-1 (TSP-1) (57) and IFN-α (58). The second is a gain-of-function event, by inducing pro-angiogenic growth factors. Among these growth factors, VEGF has being the main target of tumor angiogenesis suppression studies, due to its overexpression as part of a change in most of the tumors towards an angiogenic phenotype (59-62).
The newly synthesized blood vessels are not exactly similar to that pre-existent in healthy tissues. Tumor blood vessels show a disproportionately increased inner diameter in respect to the thick of the vascular wall, and a lower numbers of smooth muscle cells and pericytes. This implies a notable thickness and fragility of the vascular wall, favoring the development of aneurism-like dilations, irregular blood flow, and zones without blood irrigation and increased vascular permeability (63).
Besides, tumor blood vessels express specific markers as endoglins (CD-105/EDG), which are dimeric components of the cell surface acting as TGF-β receptors. This complex is abundantly expressed in the tumor endothelium but not in the normal blood vessels (44).
The EC of the immature blood vessels require signals to survive, or are eliminated by program cell death or apoptosis. The VEGF inhibits apoptosis of these EC in the immature tumor blood vessels by inducing the expression of the antiapoptotic Bcl2, A1 XIAP and surviving proteins (64).
Tumors with high levels of VEGF produce an excessively permeable vasculature susceptible to leakage, increasing the interstitial pressure inside the tumor. The morphology of tumor blood vessels also impairs blood flow, oxygen and nutrient supply. All these influence the conventional antitumoral therapy, due to a misbalanced distribution of the chemotherapeutic agents inside the tumor with some of its regions unreached by these agents, therefore, help neoplasic cells to survive. Additionally, oxygen levels are not homogeneous inside the tumor, with relatively hypoxic zones being insensitive to radiotherapy (65, 66). Consequently, the use of anti-VEGF agents could be ideal to aid chemotherapy and radiotherapy, hampering tumor growth while improving treatment for tumor destruction (67-69).
Frequently, when a primary tumor is destroyed, secondary tumors will arise if metastasis developed. In this sense, the main source of the Angiostatin (a natural endogenous antiangiogenic) is eliminated with primary tumor removal, favoring the development of secondary tumors (63). Angiostatin is commonly produced by solid tumors or cells at their environment without affecting primary tumor growth. By this mechanism, the primary tumor develops while endogenously controlling the development of distant tumors (44).
Tumors produce more angiogenesis promoting than inhibitory factors. This could be explained by the increased half life of inhibitory factors in the blood stream (for example, angiostatin circulates up to 5 days, while pro-angiogenic factors last less than 5 days). Thus, pro-angiogenic factors mainly act in the vicinity of the tumor, while inhibitors easily act on distant tumors (50).
Most of the neoplasic cells show increased genetic instability, with frequent mutations arising and causing cellular changes that generate resistance to cytotoxicity of conventional antineoplasic drugs (70). When referring to tumor genetics, several oncogenes have to be mentioned, such as V-ras, K-ras, V-raf, fos and the E6 protein of the human papillomavirus 16, all of them promoting angiogenesis by producing growth factors, cytokines and proteolytic enzymes (71, 72). On the contrary, there are tumor suppressor genes, such as Rb, c-jun and p53, the last causing HIF- 1a degradation, inhibiting VEGF production and stimulating the TSP-1 inhibitor (54).
On the other hand, EC are homogenous, diploid and genetically stable (49), showing very rarely spontaneous mutations (73). Therefore, therapeutic pharmacological strategies targeting EC will encounter less interference of the resistance processes described for other antitumorals (49).
TUMOR ANGIOGENESIS AS THERAPEUTIC TARGET
Research in the field of tumor angiogenesis follows three main directions: 1) identification of positive and negative regulators; 2) characterization of the mechanisms of action and the identification of either natural or synthetic inhibitors; and 3) quantification of the neovascularization in tumor biopsies as predictive tool for diagnosis (74).
Folkman et al. were the first to identify and isolate angiogenesis inhibitors. Their work encouraged other laboratories of the scientific-academic and pharmaceutical community throughout the world to develop angiogenesis inhibitors, some of them currently in use against cancer.
Starting from results of ongoing clinical trials (Table 1), the antiangiogenic therapy seems to be a promising strategy to treat cancer. They allow identifying several compounds showing antiangiogenic activity, with inhibitors of VEGF and its receptors as the most advanced candidates (75, 76). Recently, the Bevacizumab (Avastin), a monoclonal antibody against the VEGF-A was approved by the FDA as the first systemic antiangiogenic (77).

Other inhibitors of angiogenesis under study are the shark cartilage (78) that recently failed in a phase III clinical trial; several flavonoids as genistein, derived from soybean that is able to inhibit MMP-2 expression in prostate cancer cells (79). Additionally, there was demonstrated that apigenin, another natural flavonoid, show antiangiogenic effects involving the nitric oxide pathway (80). Other recent studies have shown the epigallocatechin-3-gallate, the main flavonoid of green tea (Camellia sinense) (81), as having antitumoral and antiangiogenic properties (82-84). One if the molecular effects of this compound is to stimulate the expression of endogenous inhibitors of MMP (TIMP-1 and TIMP-2)(85).
Another promising strategy uses the genetic immunization technology with VEGF-A DNA as active immunotherapeutic strategy (86).
Many patents have been filed of products used as antitumorals for antiangiogenic treatment: a truncated VEGFR to inhibit the expression of VEGF, used for gene therapy of colon and lung cancer; a nucleotide sequence coding for a soluble receptor that inhibits several pro-angiogenic growth factors and completely inhibit cancer angiogenesis; and the use of astaxanthin or its esters as active ingredient of products to treat cancer and other angiogenic disorders; etc. (87).
Kerbel et al. have studied other vascular modulators drugs called vascular disruption agents. They act by acutely occluding tumor vasculature, originating a vast central necrotic zone inside the tumor, although recruiting bone marrow-derived circulating endothelial precursors that cause a re-growth of tumors after treatment appreciated as a thick edge of viable cells at tumor surface. Previous results showed that the antiangiogenic therapy inhibits mobilization of levels of circulating endothelial precursors, with antiangiogenic pre-treatment increasing the therapeutic efficacy of vascular disruption agents, decreasing the thickness of the viable cells (42).
The next generation of antiangiogenic drugs should have to improve clinical efficacy by targeting multiple proangiogenic factors. This approach was recently validated by studies on oncology drugs known as multitarget tyrosine kinase inhibitors that act simultaneously on several growth factor receptors, such as: VEGFR 1 to 3, PDGFR, bFGF and the receptor of the epidermal growth factor. The pharmakynetic and pharmacodynamic properties of these compounds help to explain clinical observations currently entering in phase II and III clinical trials (88, 89).
CONCLUSIONS
There are convincing evidences on the benefits of inhibiting angiogenesis through several molecular mechanisms. This review pretended to overview the knowledge on angiogenesis and its role in cancer, also focused on the components of the signal transduction pathways involved in this process. The concept of cancer treatment in this century, considering the inhibition of angiogenesis, essential to tumor progression, represents an alternative to fight tumor resistance against therapeutic drugs. The antiangiogenic therapy is being available to surgery, chemo- and radio-therapy. The recent evaluation of antibodies, directed against VEGF as adjuvant therapy, has shown significant clinical benefits in cancer patients.
The introduction of several antiangiogenic agents or molecules in the arsenal against cancer has changed the natural history of this disease in humans. Now, the antiangiogenic drugs or medicines bring the opportunity to test not only a new class of antitumoral agent, but also a new and relevant mechanism of action. Given the success of most of the products produced by biotechnological means, the search for new products coming from natural sources constitute a significant challenge in Pharmacology up to date. Many of these and other discoveries would suggest strategies to improve the clinical benefits of the antiangiogenic therapy.
REFERENCES
1. Fischer C, Schneider M, Carmeliet P. Principles and therapeutic implications of angiogenesis, vasculogenesis and arteriogenesis. Handb Exp Pharmacol 2006;176(2):157-212.
2. Schmidt A, Brixius K, Bloch W. Endothelial precursor cell migration during vasculogenesis. Circ Res 2007;101(2):125-36.
3. Ratajska A, Czarnowska E. Vasculogenesis of the embryonic heart: contribution of nucleated red blood cell to early vascular structures. Cardiovasc Hematol Disord Drug Targets 2006;6(3):219-25.
4. Demir R, Saval Y, Hupertz B. Vasculogenesis and Angiogénesis in the early human placenta. Acta Histochem 2007;109(4):257-65.
5. Velásquez OC. Angiogenesis and vasculogenesis: inducing thr growth of new blood vessels and wound healing by stimulation of bone marrow-derived progenitor cell mobilization and homing. J Vasc Surg 2007;45 Suppl A:A39-47.
6. Ribatti D, Nico B, Cribellato E, Roccaro AM, Vacca A. The history of the angiogenic switch concept. Leukemia 2007;21(1):44-52.
7. Folkman, J. Tumor angiogenesis: therapeutic implications. N Engl J Med 1971;285(21):1182-86.
8. Scagliotti GV, Selvaggi G. The place of targeted therapies in the management of non small cell bronchial carcinoma. Target therapies in lung cancer management. Rev Mal Respir 2006;23(5 Pt 3):137-47.
9. Fischer I, Gagner JP, Law M, Newcomb EW, Zaqzaq D. Angiogenesis in gliomas: biology and molecular pathophysiology. Brain Pathol 2005;15(4):297-310.
10. Emanueli C. Angiogenesis gene therapy to rescue ischaemic tissue: achievement and future directions. Br J Pharmacol 2001;133(7):951-59.
11. Pepper MS. Role of MMP and PA/plasmina systems in angiogénesis. Arterioscler Thromb Vasc Biol 2001;17(4):605-19.
12. Walsh DA. Pathophysiological mechanisms of angiogenesis. Adv Clin Chem 2007;44:187-221.
13. Liao F, Li Y, OConnor W, Zanetta L, Bassi R, Santiago A, et al. Monoclonal antibody to vascular endotelial-cadherin is a potent inhibitor of angiogenesis tumor growth and metastasis. Cancer Res 2000;60(6):6805-10.
14. Ruegg C, Hasmim M, Lejeune FL, Alguisi GL. Antiangiogenic peptides and proteins: from experimental tools to clinical drugs. Biochim Biophys Acta 2006;1765(2):155-77.
15. Carmeliet P. VEGF as a key mediator of angiogenesis in cancer. Oncology 2005;69 Suppl 3:4-10.
16. Shing Y, Folkman J, Sullivan R, Butterfield C, Murray J, Klagsbrun M. Heparin affinity: purification of a tumor-derived capillary endothelial cell growth factor. Science 1984;223(4642):1296-9.
17. Tunyogi-Csapo M, Koreny T, Vermes C, Galante JO, Jacobs JJ, Glant TT. Role of fibroblasts and fibroblast-derived growth factor in periprosthetic angiogenesis. J Orthop Res 2007;25(10):1378-88.
18. Fugita S, Saika S, Kao WW, Fugita K, Miyamoto T, Ikeda K, et al. Endogenous TNF-alpha suppression of neovascularization stroma in mice. Invest Ophthalmol Vis Sci 2007;48(7):3051-5.
19. Izzi L, Attisana L. Ubiquitin-dependent regulation of TGF-β signaling in cancer. Neoplasia 2006;8(8):677-88.
20. Naldini A, Leali D, Pucci A, Morena E, Carraro F, Nico B, et al. Cutting edge: IL-1 beta mediates the proangiogenic activity of osteopontin-activated human monocytes. J Immunol 2006;177(7):4267-70.
21. Mikroulis D, Papanas N, Maltezos E, Bougioukas G. Angiogenic growth factors in the treatment of peripheral arterial disease. Curr Vasc Pharmacol 2007;5(3):195-209.
22. Hoffmann S, Glaser S, Wunderlich A, Lingelbach S, Dietrich C, Burchert A, et al. Targeting the EGF/VEGF-R system by tyrosine-kinase inhibitors-a novel antiproliferative/ antiangiogenic strategy in thyroid cancer. Langenbecks Arch Surg 2006;391(6):589-96.
23. Magnusson PU, Looman C, Ahgren A, Wu Y, Claesson-Welsh L, Heuchel RL. Platelet-derived growth factor receptor beta constitutive activity promotes angiogenesis in vivo and in vitro. Arterioscler Thromb Vasc Biol 2007;27(10):2142-9.
24. Borkakoti N. Matrix Metalloprotease inhibitors. Biochem Soc Trans 2004;32(Pt 1):17-20.
25. Bogaczewicz J, Dudek W, Zubilewicz T, Wronski J. The role of matrix metalloproteinases and their tissue inhibitors in angiogenesis. Pol Merkur Lekarski 2006;21(121):80-5.
26. Davis GE, Saunders WB. Molecular balance of capillary tube formation versus regression in wound repair: role of matrix metalloproteinases and their inhibitors. J Investig Dermatol Symp Proc 2006;11(1):44-56.
27. Cauchard JH, Robinet A, Poitevin S, Bobichon H, Maziere JC, Bellon G, et al. UVA-mediated down-regulation of MMP- 2 and MT1-MMP coincides with impaired angiogenic phenotype of human dermal endothelial cells. Biochem Biophys Res Commun 2006;345(2):681-7.
28. Rundhaug JE. Matrix metalloproteinase, angiogenesis and cancer. Clin Cancer Res 2003;9(2):551-4.
29. Oku N, Matsukawa M, Yamakawa S, Asai T, Yahara S, Hashimoto F, et al. Inhibitory effect of green tea polyphenols on membranetype 1 matrix metalloproteinase, MT1-MMP. Biol Pharm Bull 2003;26(9):1235-8.
30. Bogusiewicz M, Stryjecka-Zimmer M, Rechberger T. Activity of matrix metalloproteinases-2 and -9 (MMP-2 and MMP-9) and content of their tissue inhibitors in endometrial cancer-a preliminary study. Ginekol Pol 2007;78(5):366-72.
31. Hrabec E. A novel matrix metalloproteinase inhibitor in patients with advanced cancer of the ovary. J Cancer Res Clin Oncol 2002;128(8):197-204.
32. Dano K, Behrendt N, Hoyer-Hansen G, Johnsen M, Lund LR, Ploug M, et al. Plasminogen activation and cancer. Thromb Haemost 2005;93(4):676-81.
33. Dunehoo AL, Anderson M, Majumdar S, Kobayashi N, Berkland C, Siahaan TJ. Cell adhesion molecules for targeted drug delivery. J Pharm Sci 2006;95(9):1856-72.
34. Liebner S, Caballaro U, Dejana E. The multiple languages of endothelial cell-tocel communication. Arterioscler Thromb Vasc Biol 2006;26(7):1431-8.
35. Humphries MJ. Adhesion Molecules as important angiogenesis components. Trends Pharmacol Sci 2000;21(1):29.
36. Arenillas JF, Sobrino T, Castillo J, Dávalos A. The role of angiogenesis in damage and recovery from ischemic stroke. Curr Treat Options Cardiovasc Med 2007;9(3):205-12.
37. Miura S, Saku K. Regulation of angiogenesis and angiogenic factors by cardiovascular medications. Curr Pharm Des 2007;13(20):2113-7.
38. Koch AE, Distler O. Vasculopathy and disordered angiogenesis in selected rheumatic diseases: rheumatoid arthritis and systemic sclerosis. Arthritis Res Ther 2007;9 Suppl 2:S3.
39. Koutroubakis IE, Xidakis C, Karmiris K, Sfiridaki A, Kandidaki E, Kouroumalis EA. Potential role of soluble angiopoietin-2 and Tie-2 in patients with inflammatory bowel disease. Eur J Clin Invest 2006;36(2):127-32.
40. Hernández C, Simo R. Strategies for blocking angiogenesis in diabetic retinopathy: from basic science to clinical practice. Expert Opin Investig Drugs 2007;16(8):1209-26.
41. Hasan Q, Tan ST, Gush J, Peters SG, Davis PF. Steroid therapy of a proliferating hemangioma: histochemical and molecular changes. Pediatrics 2000;105(1):117-20.
42. Man XY, Zheng M. Advances in pathogenesis of psoriasis. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2006;35(6):673-7.
43. Nisolle M, Alvarez ML, Colombo M, Foidart JM. Pathogenesis of endometriosis. Gynecol Obstet Fertil 2007;35(9):898-903.
44. Balakin KV, Tkachenko SE. VEGF/VEGFR signalling as a target for inhibiting Angiogenesis. Expert Opin Investig Drugs 2007;16(1):83-107.
45. Klement H, St Croix B, Milsom C, May L, Guo Q, Yu JL, et al. Atherosclerosis and vascular aging as modifiers of tumor progression, angiogenesis, and responsiveness to therapy. Am J Pathol 2007;171(4):1342-51.
46. Tapping a research-rich vein (Editorial). Nat Med 2003; (6): 627.
47. Chaplain MA, McDougall SR, Anderson AR. Mathematical modeling of tumor-induced angiogenesis. Annu Rev Biomed Eng 2006;8:233-57.
48. Masferrer JL, Leahy KM, Koki AT, Zweifel BS, Settle SL, Woerner BM, et al. Antiangiogenic and antitumor activities of cyclooxygenase-2 inhibitors. Cancer Res 2000;60(5):1306-11.
49. López O, Bequet M, Hernández P, Araña M de J, Perea SE, López P. Actividad angiogénica en el carcinoma cervical asociado a virus de papiloma humano: oncogenes virales como blanco terapéutico. Biotecnol Apl 2001;18(2):95-6.
50. Kyzas PA, Stefanou D, Batistatou A, Agnantis NJ. Hypoxia-induced tumor angiogenic pathway in head and neck cancer: an in vivo study. Cancer Lett 2005;225(2):297-304.
51. Lewis CE, Pollard JW. Distinct role of macrophages in different tumor microenvironments. Cancer Res 2006;66(2):605-12.
52. Dome B, Hendrix MJ, Paku S, Tovari J, Timar J. Alternative vascularization mechanism in cancer: pathology and therapeutic implications. Am J Pathol 2007;170(1):1-15.
53. van den Beucken T, Koritzinski M, Wouters BG. Translational control of gene expression during hypoxia. Cancer Biol Ther 2006;5(7):749-55.
54. Murdoch C, Lewis CE. Macrophage migration and gene expression in response to tumor hypoxia. Int J Cancer 2005;117(5):701-8.
55. Brahimi-Horn C, Pouyssequr J. The role of hypoxia inducible factor in tumor metabolism growth and invasion. Bull Cancer 2006;93(8):E73-80.
56. Kang JH, Kim SA, Chang SY, Hong S, Hong KJ. Inhibition of trichostatin A-induced antiangiogenesis by small-interfering RNA for thrombospondin-1. Exp Mol Med 2007;39(3):402-11.
57. Yanase N, Kanetaka Y, Mizuguchi J. Interferon-alpha-induced apoptosis via tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-dependent and -independent manner. Oncol Rep 2007;18(4):1031-8.
58. Lieto E, Ferraraccio F, Orditura M, Castellano P, Mura AL, Pinto M, et al. Expression of vascular endothelial growth factor (VEGF) and epidermal growth factor receptor (EGFR) is an independent prognostic indicator of worse outcome in gastric cancer patients. Ann Surg Oncol 2008;15(1):69-79.
59. Kopczynska E, Dancewicz M, Kowalewski J, Kardymowicz H, Tyrakowski T. The estimation of serum concentration of vascular endothelial growth factor in patients with non-small cell lung cancer. Pol Merkur Lekarski 2007;22(132):536-8.
60. Timar J, Paku S, Tovari J, Dome B. Rationale of antiangiogenic therapy. Magy Onkol 2006;50(2):141-51.
61. Dallas NA, Fan F, Gray MJ, Van Buren G, Lim SJ, Xia L, et al. Functional significance of vascular endothelial growth factor receptors on gastrointestinal cancer cells. Cancer Metastasis Rev 2007;26(3-4):433-41.
62. Karter SK. Clinical strategy for the development of angiogenesis. Oncologist 2000;23(9):51-3.
63. Ferrara N. Vascular endothelial growth factor and the regulation of angiogenesis. Recent Prog Horm Res 2000;55:15-35.
64. Griffioen AW. Cancer is angiogenesis dependent. Blood 2000;95(8):3023-34.
65. Lee H. Molecular and cellular aspects of tumor cell invasion. Nat Med 2000;5(1):167-70.
66. Zhong H, Bowen JP. Antiangiogenesis drug design: multiple pathways targeting tumor vasculature. Curr Med Chem 2006;13(8):849-62.
67. Hurwitz KG. Understanding cancer. Cancer Res 2004;16(6):38-50.
68. Jain DC. Tumor angiogenesis: vascular growth in solid tumors. Ann Oncol 2001;5(2):329-36.
69. Ching TT. The COX-2 inhibitors in human cancer. J Biol Chem 2000;275(8):11397-403.
70. Kerbel R. Tumor angiogenesis: past, present and the near future. Carcinogenesis 2000;21(1):505-15.
71. López-Ocejo O, Viloria-Petit A, Bequet-Romero M, Mukhopadhyay D, Rak J, Kerbel RS. Oncogenes and tumor angiogenesis: the HPV-16 E6 oncoprotein activates the vascular endothelial growth factor (VEGF) gene promoter in p53 independent manner. Oncogenes 2000;19:4611-20.
72. Quesada AR, Chapuli RM, Medina MA. Anti-angiogenic drugs: from bench to clinical trials. Med Res Rev 2006;26(4):483-530.
73. Ferrara N, Kerbel R. Angiogenesis as a therapeutic target. Nature 2005;438:967-74.
74. Zhong H, Bowen JP. Molecular design and clinical development of VEGFR kinase inhibitors. Curr Top Med Chem 2007;7(14):1379- 93.
75. Shojaei F, Ferrara N. Antiangiogenesis to treat cancer and intraocular neovascular disorders. Lab Invest 2007;87(3):227-30.
76. Grossi F, Aita M. Bevacizumab and nonsmall-cell lung cancer: starving the enemy to survive. Expert Opin Biol Ther 2007;7(7):1107-19.
77. Hassan ZM, Feyzi R, Sheikhian A, Bargahi A, Mostafaie A, Mansouri K, et al. Low molecular weight fraction of shark cartilage can modulate immune responses and abolish angiogenesis. Int Immunopharmacol 2005;5(6):961-70.
78. Kumi-Diaka JK, Hassanhi M, Merchant K, Horman V. Influence of genistein isoflavone on matrix metalloproteinase-2 expression in prostate cancer cells. J Med Food 2006;9(4):491-7.
79. Erdogan A, Most AK, Wienecke B, Fehsecke A, Leckband C, Voss R, et al. Apigenin-induced nitric oxide production involves calciumactivated potassium channels and is responsible for antiangiogenic effects. J Thromb Haemost 2007;5(8):1774-81.
80. Katiyar S, Elmets CA, Katiyar SK. Green tea and skin cancer: photoinmunology, angiogenesis and DNA repair. J Nutr Biochem 2007;18(5):287-96.
81. Zhang Q, Tang X, Lu Q, Zhang Z, Rao J, Le AD. Green tea extract and (-)-epigallocatechin-3-gallate inhibit hypoxia- and seruminduced HIF-1alpha protein accumulation and VEGF expression in human cervical carcinoma and hepatoma cells. Mol Cancer Ther 2006;5(5):1227-38.
82. Carlson Jr, Bauer BA, Vincent A, Limburg PJ, Wilson T. Reading the tea leaves: anticarcinogenic properties of (-)-epigallocatechin3- gallate. Mayo Clin Proc 2007;82(6):725-32.
83. Kerbel RS. Antiangiogenic therapy: a universal chemosensitization strategy for cancer? Science 2006;312(5777):1171-5.
84. Garbosa S. Tumor gelatinases and invasion inhibited by the green tea flavanol epigallocatechin-3-gallate. Cancer 2001;4(2):822-32.
85. Bequet-Romero M, Ayala M, Acevedo BE, Rodríguez EG, López-Ocejo O, Torrens I, et al. Prophylactic naked DNA vaccination with the human vascular endothelial growth factor induces an anti-tumor response in C57BL/6 mice. Angiogenesis 2007;10:23-34.
86. Harrison Ch. Patent watch. Nat Rev Drugs Disc 2007;6:262-3.
87. Shaked Y, Ciarocchi A, Franco M, Lee CHR, Man S, Cheung AM, et al. Therapy-induced acute recruitment of circulating endothelial progenitor cells to tumors. Science 2006;313:1785-7.
88. Hilberg F, Roth GJ, Krssak M, Kautschitsch S, Sommergruber W, Tontsch-Grunt U, et al. BIBF 1120: triple angiokinase inhibitor with sustained receptor blockade and good antitumor efficacy. Cancer Res 2008;68(12):4774-82.
89. Spannuth WA, Sood A, Coleman RL. Angiogenesis as a strategic target for ovarian cancer therapy. Nat Clin Pract Oncol 2008;5(4):194-204.
Received in December, 2007.
Accepted for publication in October, 2008.
Omar López. Centro de Estudio para las Investigaciones y Evaluaciones Biológicas, IFAL, UH. Ave. 23 #21425 entre 214 y 222, La Coronela, La Lisa, Ciudad de La Habana, Cuba. E-mail: omarlopez@infomed.sld.cu
